
Abstract
Voluntary runners are subjected to a massive increase in reactive oxygen/nitrogen species production, which can promote different oxidative stress–related diseases such as premature aging, neurodegenerative disorders, and cancer. The aims of this work were to evaluate the following in peripheral blood cells of voluntary runners: (i) DNA status; (ii) susceptibility to the in vitro insult induced by hydrogen peroxide (H2O2) as a breaking agent; (iii) capabilities of 3,5,4′-trihydroxystilbene (RESV) in counteracting DNA damage. Twenty-five male voluntary runners were compared with 20 sedentary men, as age-matched controls, and DNA status was evaluated with different versions of comet assay: alkaline, neutral, and Fpg enzyme–modified version to measure 8-OH-deoxyguanosine (8-oxo-dG) levels. The H2O2 and/or RESV treatments were performed directly on agarose-embedded cells (atypical comet assay). The results evidenced DNA damage and levels of 8-oxo-dG higher in runners than in sedentary control subjects. The runners' DNA was more prone to the in vitro–induced oxidative insult (200 μM H2O2) than that of the control group. Resveratrol (100 μM), depending on the individual basal DNA status, was able to switch from antioxidant to pro-oxidant. Our results, on the one hand, validated the proposed in vitro experimental protocol in order to measure individual DNA status. On the other hand, our data point out the importance of monitoring the athletes' redox status before subjecting them to dietary supplementation treatment.
Introduction
R
Redox imbalance is believed to play a causative role in a plethora of unwanted effects or diseases, as well as premature aging; cardiovascular, hepatic, and neurodegenerative diseases; inflammatory conditions; obesity; cancer susceptibility; and cell death. 2 –5 These effects are exerted differently according to individual variability linked with genetic variants in oxidative stress–related genes, as single-nucleotide polymorphisms in some cytosolic and/or mitochondrial antioxidant enzymes. 6
Regular aerobic exercise is well known to modulate redox condition and ameliorate quality of life, particularly in older people, in which it decreases morbidity and mortality. 4,5 However, a body of literature data has demonstrated that long-term activities, having both elevated energy expenditure and high oxygen consumption, may promote ROS production and oxidative stress conditions, depending on duration and type of exercise, as well as on the individual's lifestyle and eating habits. 5,7
Human biochemical markers of oxidative stress are usually measured in plasma and in blood cells by several methods. 8,9 Recently, there has been growing interest in exercise-induced DNA damage due to its involvement in various diseases, including carcinogenesis and neurodegeneration. 5,10,11
Previous studies have used a variety of experimental approaches to measure DNA status/damage, such as high-performance liquid chromatography, 8-OH-deoxyguanosine (8-oxo-dG) assay, micronuclei test, chromosome aberration, and sister chromatid exchanges. These methods require expertise handling large amounts of DNA and are not usually cost or time effective. 11,12
Among these, comet assay is widely considered a feasible, sensitive, low-cost, and valuable tool in monitoring, both in vivo and in vitro, DNA status. This method also permits evaluating different kinds of DNA damage induced by endogenous and/or exogenous factors, as well as the effects of some antioxidant supplementations and therapies. 11,13 –15
Natural antioxidant molecules have been extensively considered in preventing and/or reducing oxidative stress and DNA damage. 7,8,16 Among natural compounds, 3,5,4′-trihydroxystilbene (RESV), found in various plants, has attracted particular attention in the last decade for its innumerable properties. In fact, it exerts antioxidant, antiinflammatory, antiatherogenic, immunomodulator, chemopreventive, chemiotherapeutic, in vitro, and in vivo properties, as well as increased life span in yeasts, flies, and rats. 17 –19
There is, however, growing evidence that RESV exhibits pro-oxidant activities under some experimental conditions. 20 Nonetheless, the precise mechanisms exerted by resveratrol on various targets still need to be better elucidated.
The purpose of this study was to validate a protocol as a useful tool in examining the same sample: (i) the global DNA status and its 8-oxo-dG level (alkaline, neutral, and Fpg-modified comet assay versions); (ii) the susceptibility of runners' DNA to hydrogen peroxide (H2O2) in vitro–induced insult; (iii) the in vitro capability of RESV in influencing DNA status (atypical comet assay). As control samples, sedentary subjects were considered.
Materials and Methods
Inclusion criteria for subject participation were 25 male runners and 20 sedentary controls, aged 54±8, nonsmokers, and VO2max value classified as having excellent fitness levels. Subjects were nourished with a Mediterranean controlled diet, free of antioxidant supplementation, for one month before being tested. The daily run occurred under the same environmental conditions, intensity, distance, and length of weekly training (Table 1).
Informed consent from all patients was obtained in accordance with the recommendations of the declaration of Helsinki.
Blood samples (70–90 μL) were collected from each subject by finger pricks (in the group of marathon runners, 30 min after the end of daily physical activity), immediately added to 1 mL of solution A (Ca2+/Mg2+-free Hank's balanced salt solution, 20 mM ethylenediaminetetraacetic acid [EDTA], and 10% dimethyl sulfoxide, pH 7.5–7.7), and centrifuged at 800 g for 10 min. The pellet was suspended in 200 mL of phosphate-buffered saline 1×, separated in five aliquots (each 40 μL), and stored at −80°C for 10 days. 14,21 DNA damage was measured by different comet assay versions: (i) alkaline and/or neutral typical version 22 to identify different types of DNA damage; (ii) Fpg-modified version measures the 8-oxo-dG level 14 ; (iii) atypical version was performed, adding 200 μM H2O2 and/or 100 μM resveratrol directly on agarose-embedded whole blood cells, considered a vital system. 15
Eighty microliters for each sample, suspended in 520 μL of 0.5% low-melting-point agarose, was spread on a Trevigen microscope slide (20 wells) and allowed to set at 4°C for 20 min.
The tests were performed in duplicate (Fig. 1). The timing for the preparation of each slide was synchronized to simultaneously immerse all the samples in the freshly prepared lysis solution A (2.5 M NaCl, 100 mM Na2 EDTA, 10 mM Tris, NaOH [pH 10.0–10.5], and 1% lauroyl sarcosine; 1% Triton X-100 and 10% dimethyl sulfoxide were added directly before use) for 1 h at 4°C.
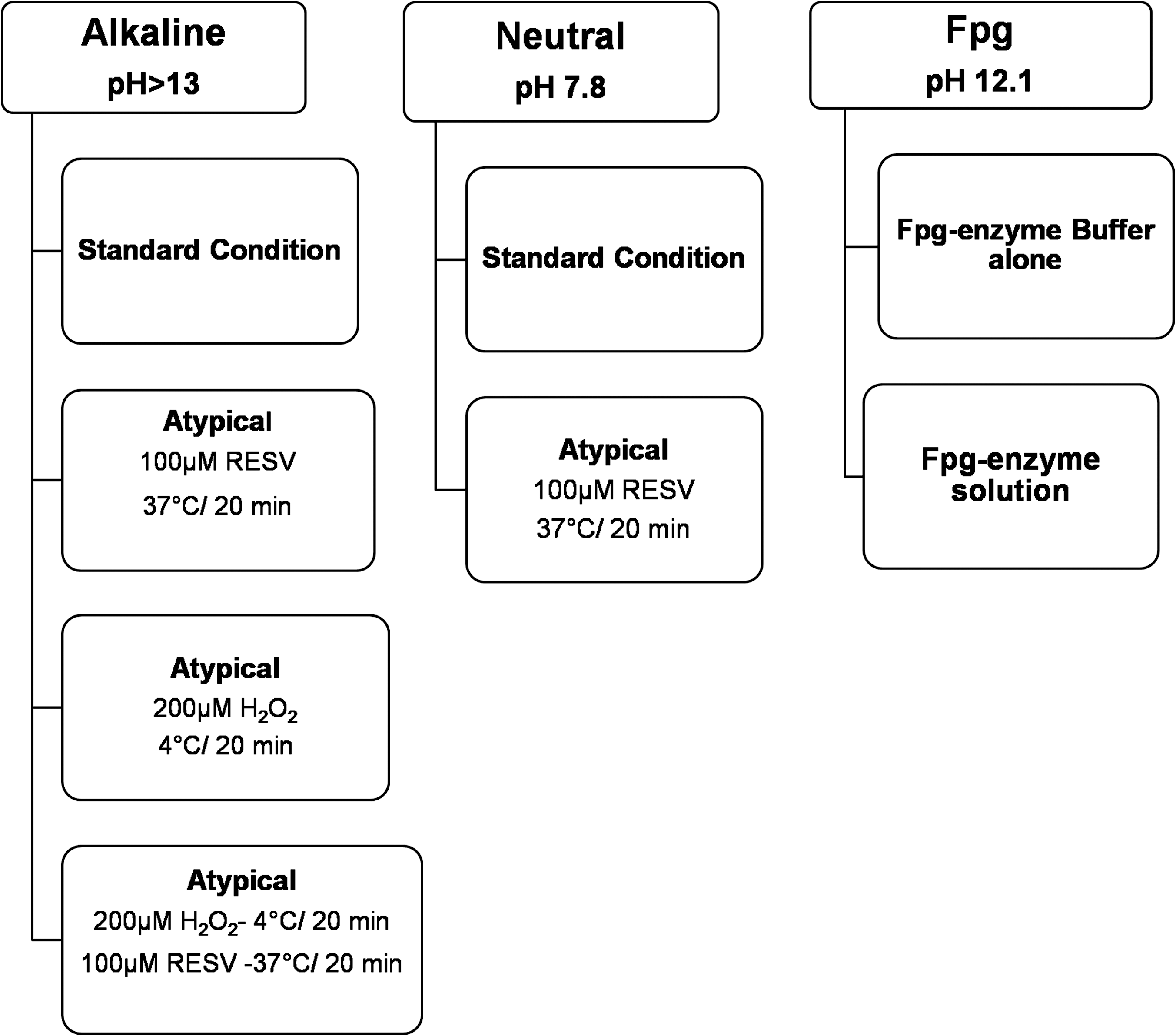
The study design planned to evaluate DNA status/damage by the different types of comet assay.
The control samples were only processed for alkaline and Fpg versions as previously described for runners' samples (Fig. 1).
Alkaline and neutral comet assay
After lysis in solution A, the samples were maintained for 20 min in a high-pH buffer (300 mM NaOH and 1 mM Na2 EDTA [pH 13.1]) to promote DNA unwinding, and then electrophoresed in the same buffer at 0.7 V/cm for 30 min for the alkaline version, and in buffer B (90 mM Tris-base, 90 mM boric acid, and 2 mM Na2 EDTA [pH 8]) at 0.5 V/cm for 20 min for the neutral version.
Fpg-modified comet assay
The Fpg FLARE™ assay kit (Trevigen, Inc., Gaithersburg, MD) was used to analyze 8-oxo-dG level, according to the manufacturer's protocol. Naked DNA (or nucleoid) was digested with Fpg DNA repair enzyme that recognizes and cuts the sites corresponding to oxidized guanine bases. For each sample, 35 μL of agarose-embedded whole blood cell suspensions was dropped in duplicate on two different slides. After 20 min at 4°C in the dark, slides were immersed in a prechilled lysis solution (provided with the FLARE assay kit), at 4°C for 60 min. After lysis, 100 μL of enzyme dilution buffer and/or Fpg enzyme solution (prediluted 1:10 in REC™ dilution buffer [Trevigen, Inc.]) was added to the samples and then placed in the humidity chamber at 37°C for 45 min. The slides were immersed twice (10 min each) in prechilled alkaline electrophoresis solution (0.3 M NaOH and 1 mM Na2 EDTA [pH 12.1]), and then electrophoresed at 0.7 V/cm for 20 min.
Atypical comet assay
Atypical comet assay was performed, treating the agarose-embedded cells with 70 μL of 100 μM RESV at 37°C for 20 min (Fig. 1), 200 μM H2O2 at 4°C for 20 min, or with 200 μM H2O2 plus 100 μM RESV, as reported in Figure 1. Then, the slides were processed as reported for standard alkaline version. 15
At the end of electrophoresis, all the slides were neutralized in the Tris buffer (0.4 M Tris-HCl [pH 7.5]), dehydrated with 70% ethanol, and stained with 40 μL of Syber Green. One hundred nucleoids per sample (50 for each of the two replicates) were randomly analyzed using the Leika epifluorescence microscope (Leika, Wetzlar, Germany), interfaced with a computer. The percentage of damaged DNA in the tail of the comet (%TDNA) was chosen as a parameter to measure DNA damage by the CASP free software. The data acquired by CASP were expressed also as a percentage of the values present in five different arbitrarily chosen classes, corresponding to different ranges of the %TDNA values: I (undamaged cells: from 0% to 14% TDNA), II (low damaged cells: from 15% to 31% TDNA), III (moderate damaged cells: from 32% to 51% TDNA), IV (high damaged cells: from 51% to 65% TDNA), and V (very high damaged cells: >65% TDNA). In addition, the values related to 8-oxo-dG level were also converted before hand to DNA break/109 Da/Gy, standard units, obtained through an indirect calibration, according to Collins et al., 23 and then as breaks/106 bp (altered bases/106 unaltered bases). 24
The analysis, regardless of the electrophoretic conditions, did not include the lateral parts of the gel, the edges, superimposed comets, and comets without distinct heads.
All the operations were kept in the dark. All the tests performed in duplicate were repeated a minimum of three times.
Statistical analyses
To look for differences between the groups, the statistical significance was assessed by the t-test for dependent samples. Data were reported as means±standard error of the mean (SEM) and values of P<.05 were considered to be statistically significant.
Results
The standard alkaline and Fpg-modified comet assay version evidenced an increase in %TDNA in runners, compared with the controls, as shown in Table 2. The data also showed that the level of 8-oxo-dG was higher in all the enrolled runners than in the sedentary controls.
Values are expressed as mean±standard error of the mean. The results expressed as %TDNA are also converted into (i) breaks/109 Da using for calculation of DNA break frequencies the calibration curve reported by Collins et al., 23 and (ii) 8-oxo-dG/106 dG using as conversion factor 1 8-oxo-Gua/109 Da is equivalent to 1 8-oxo-Gua/0.61×106 Gua as reported by Griffiths et al. 24
P<.001: significant versus correspondent control values at pH>13 and/or pH 12.1 (buffer and Fpg).
P<.001: significant versus correspondent buffer values.
%TDNA, percentage of damaged DNA in the tail of the comet.
However, there was a high inter-individual variability among runners' values measured by standard alkaline and/or neutral comet assay. To better highlight the individual differences, we chose to distribute the data (%TDNA values) of each runners' group into five different arbitrarily chosen classes (from I to V), corresponding to different ranges of %TDNA values as reported in the Materials and Methods section.
This representation allowed us to evidence that in eight runners (group A), among the 25 enrolled subjects, almost all nucleoids were included in the highest classes of damage (from III to V) (Fig. 2a). In the same runners, high levels of double strand breaks (DSBs) were evidenced by neutral comet assay (Fig. 2c).
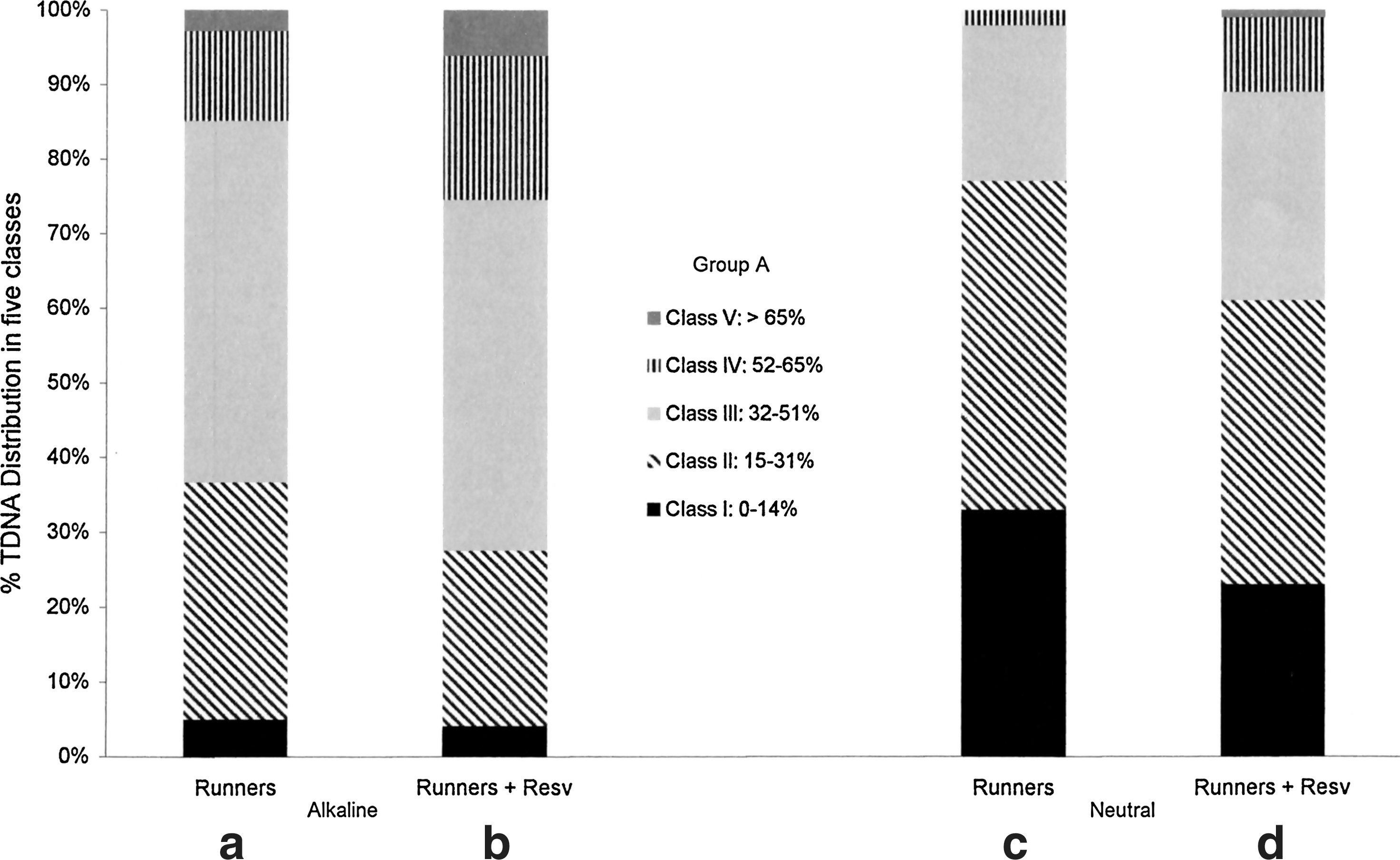
Distribution of DNA damage for Group A, expressed as five different ranges of percentage of damaged DNA in the tail of the comet (%TDNA; classes I–V as in the Materials and Methods section). Values reported in each class represent the mean of data obtained from eight athletes. Alkaline
The addition of 100 μM RESV to the samples of group A exacerbated DNA damage, augmenting in each subject the number of nucleoids in IV/V classes and in III/IV/V for alkaline and neutral assay, respectively (Fig. 2b, d).
The other 17 runners (group B) showed a large percentage of nucleoids in classes I and II (Fig. 3a). The presence of DSBs, as revealed by neutral comet assay, was very low. In this group of runners, RESV was able to ameliorate DNA status (Fig. 3b, d).

Distribution of DNA damage for Group B, expressed as five different ranges of %TDNA (classes I–V as in Materials and Methods section). Values reported in each class represent the mean of data obtained from 17 athletes. Alkaline
In view of the just discussed results, we chose to treat group B subjects (n=17 athletes), showing low DNA damage level, with H2O2 alone or combined with RESV and to examine their effects by standard alkaline and neutral versions.
The results are reported in Figure 4; the observed basal data of these subjects were utilized as control values (Fig. 3a, c). On one hand, results evidenced an H2O2-induced increase in DNA damage, particularly in alkaline buffer conditions (Fig. 4a), when compared with their basal data (Fig. 3). On the other hand, we observed that the oxidative-induced insult was only partially reverted by the sequential addition of RESV (Fig. 4b, d).
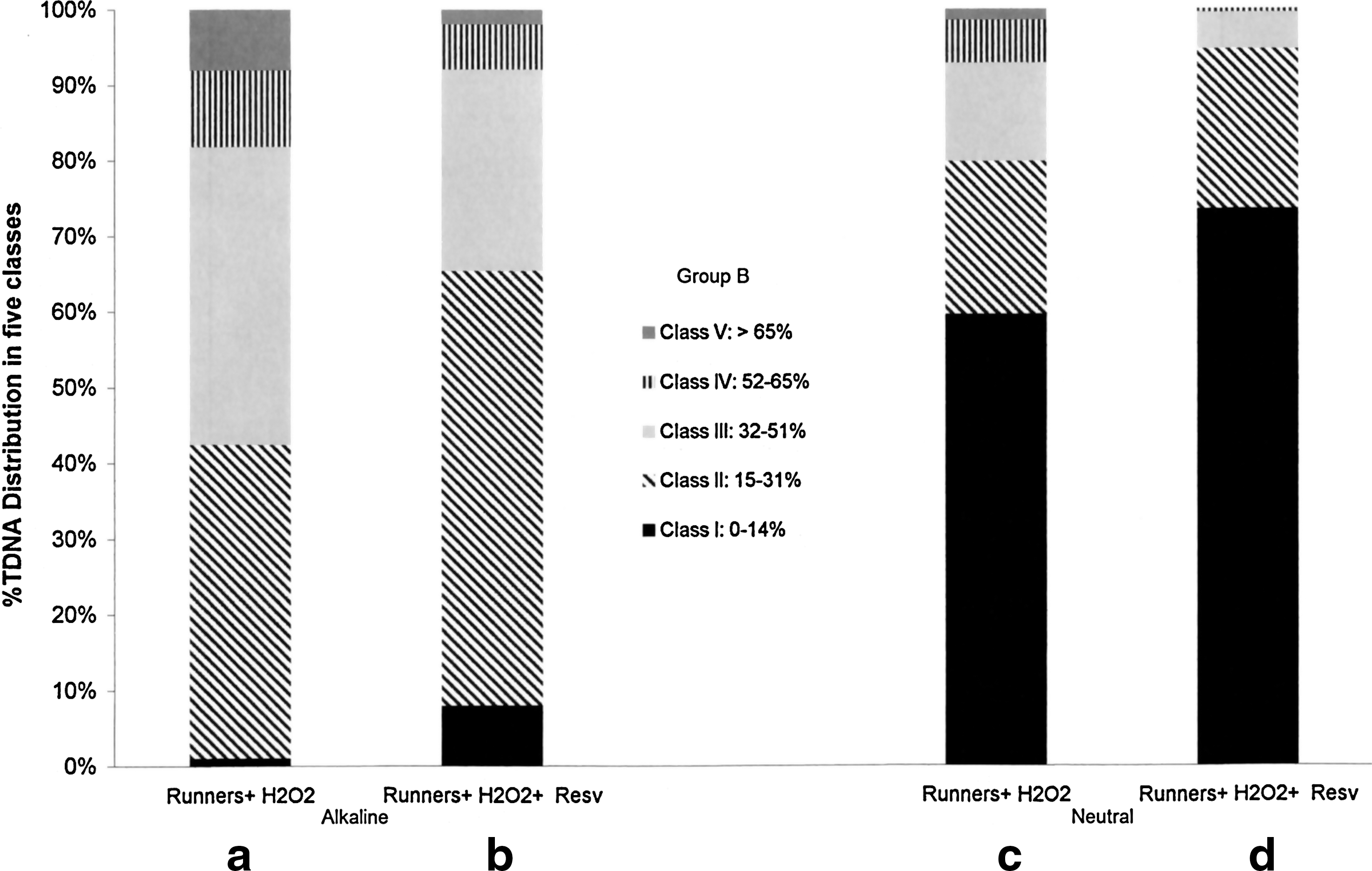
Distribution of DNA damage for Group B, expressed as five different ranges of %TDNA (classes I–V as in Materials and Methods section). Values reported in each class represent the mean of data obtained from 17 athletes. Alkaline
Discussion
Regular aerobic physical training improves quality of life, functional performance, and mean lifespan; it also reduces the risk of cardiovascular disease, upregulates immune response, and DNA repair enzyme activities, increasing cellular stress resistance by hormetic mechanisms. 5,7 Paradoxically, long-term runners, having both elevated energy expenditure and high oxygen consumption, are subjected to a massive imbalance of cellular redox status resulting in DNA damage, which is highly responsible for different types of tissue dysfunctions. 5,7,8
The increase of %TDNA reported by us in runners' whole blood cells, in agreement with data of other authors, demonstrated that regular exercise alone may not induce an adaptive response, probably depending on the individual's lifestyle and diet. 5 Furthermore, it must be considered that the level of DNA damage, at such a point, reflects the ratio between DNA damage and DNA repair. 10 In turn, the level of DNA repair is regulated differently in each individual by the expression of numerous genes involved in the different repair systems. The high levels of 8-oxo-dG observed in runners' DNA confirm the link between oxidative stress and intense physical activity; this damage may also be supported by other metabolic alterations due to daily activity, that is, an inflammatory condition. 11
DNA damage has already been reported in endurance exercise, above all in peripheral human white cells and in leukocytes. These cells are good indicators of systemic situation but may reflect changes partially at the tissue level. 11 –13
Many researchers have studied DNA damage on white blood cells (WBCs), purified lymphocytes, as well as on whole blood cells, the latter today being preferred to lymphocytes, in order to avoid the artefacts occurring during isolation of cell types. 13,21 Literature data warn against the use of whole blood cells in in vitro studies, because of the possible interference of red blood cells (RBCs) with some treatments and/or the possible contribution of catalase and glutathione peroxidise activities in suppressing DNA damaging activity elicited by various molecules, such as H2O2. 25 Furthermore, another limitation in utilizing WBCs in human studies is due to the wide variability in number, composition, and/or antioxidant capacity of WBCs, factors that could make interpreting the results difficult. 25 In our opinion, this obstacle may be overcome by the advantage of single-cell comet assay that permits integrating/evaluating the effects elicited by the treatments on the different types of blood cells. However, our experience with WBCs, utilizing typical and atypical comet assay versions, confirms that whole blood cells, if opportunely treated, maybe preferred to lymphocytes. 9,14,21 Conversely to Andreoli et al., 26 in fact, we observed that the treatment with 200 μM H2O2 at 4°C for 20 min on agarose-embedded whole blood cells (atypical comet assay) was able to induce DNA damage in the runners' white cells, confirming that interference between treatments in situ and RBCs could be bypassed using adequate concentrations of substances.
Various researchers have also demonstrated the efficacy of some antioxidant molecules in reducing/preventing DNA damage or oxidative stress in athletes. However, these molecules seem to be effective in one cellular system but may fail to protect other systems, in which they may induce a damage. 7 –9 This dual response may depend on individual lifestyle and on genetic profile, but also on the different molecule concentrations used in the treatment, evidencing the pleiotropic activity of certain antioxidants, as it may be for RESV. 20 In our experimental conditions RESV in vitro treatment reduced DNA damage in only 17 of the 25 runners who exhibited a low percentage of blood cells falling into III, IV, and V classes (Fig. 3). Recently Moller and Loft 27 indicate that supplementation with antioxidants in sedentary subjects had little effect on DNA damage and a protective effect only in the presence of a stressor. We treated Group B runners' blood cells with H2O2 as the stressor, observing a protective effect elicited by RESV.
Considering our results, we hypothesize that positive effects exerted by RESV in these subjects, having moderate basal damage and low DSBs level, maybe mediated by activation of cellular repair enzyme machinery, since agarose-embedded cells are able to maintain the efficient cellular enzymatic activities. This response may be due to a direct interaction between the phytoalexin and oxidized DNA bases (i.e., 8-oxo-guanidine), or to a possible direct antioxidant effect of RESV on cellular ROS production. When the stilbene was sequentially added to H2O2, it switched off the production of ROS, “quenching” H2O2.
Conversely, we observed that RESV behaved as a DNA damaging agent in Group A (eight runners) having per se a high percentage of nucleoids in high classes of damage, as well as high level of DSBs (Fig. 2). This type of damage usually arises during DNA repair and recombination, and sometimes leads to chromosomal aberrations, breaks, and translocations as well as oncogenic transformations. 20,28 These athletes, each di per se, could be in an oxidative stress condition due to either a deficiency in some antioxidant systems or nonfunctioning DNA repair systems. In this instance, we speculate that RESV may be unable to scavenge radical OH•, mainly responsible for oxidative DNA modifications. In contrast, RESV can promote increased oxidative DNA damage, directly exerting its DNA-cleavage activity, an effect usually observed in the presence of some ions such as Cu and Fe. 28,29 Also, the lack of protective effects by RESV in runners maybe related to elevated blood levels of ascorbic acid, as already demonstrated. 11
Our research confirms (i) the flexibility of the different comet assay versions in measuring different parameters, useful in performing a targeted therapeutic intervention; and (ii) the double-face behavior of resveratrol (100 mM in vitro), which switches from antioxidant to pro-oxidant. Our findings on stilbene suggest that its activity is dependent not only on the type of cells/tissue, concentrations/times of treatment, as reported, but also on individual DNA status and probably on the subjects' concentration of some metal ions (Cu+ and Fe).
Our results encourage the use of comet assay and underline “the good, the bad, and the ugly” 30 properties of resveratrol, and/or probably of its metabolites. In addition, they point out the need for a greater control in the wide use of some micronutrients as diet supplements, in particular when the objective is to ameliorate the athletes' performance.
It is important to take into account the well-known inter-individual variability, both at basal level and in response to treatment, by monitoring the level of oxidative status through appropriate testing before a daily intake of diet supplements is commenced.
Footnotes
Acknowledgment
This work was supported by the grant of MIUR-Ateneo (60%) (to M.R.).
Authors' Contributions
The authors' responsibilities were as follows: B.T., S.G., and S.S. conducted the experiments and collected the data; G.M. supported B.T. in analyzing data and in manuscript revision; M.F. utilized the COMET software, acquiring data and elaborating statistical data; M.R. designed the study, analyzed data, supervised its collection, and wrote the manuscript, whose revision was collegial.
Author Disclosure Statement
The authors declare that they have no competing financial interests.