
Abstract
An experiment was conducted to study the protective effect of feeding extruded and unextruded blueberry pomace (BBP) on selected metabolic parameters associated with metabolic syndrome in a model of high fructose (HF)-fed growing Sprague–Dawley rats. Treatments were as follows: (1) control (modified AIN-based diet); (2) HF diet (AIN diet with 58% fructose); (3) HF diet with 1.5% unextruded BBP; (4) HF diet with 1.5% extruded BBP; (5) HF diet with 3% unextruded BBP; and (6) HF diet with 3% extruded BBP. Compared with the control, HF feeding increased fasting plasma insulin and fasting and postprandial plasma triglycerides as well as homeostatic scores of insulin resistance and β-cell function, but not weight gain, diet intake and efficiency, abdominal fat, oral glucose tolerance, and fasting and postprandial plasma glucose, cholesterol, and leptin levels. Inclusion of unextruded or extruded BBP was effective in minimizing or ameliorating the fructose-induced metabolic anomalies, except postprandial plasma triglycerides, especially at 3% of the diet. In addition, unextruded or extruded BBP at 3% of the diet was also able to reduce plasma cholesterol and abdominal fat relative to the HF control, which may impart additional health benefits. Compared with the control, inclusion of unextruded or extruded BBP at both 1.5% and 3% resulted in lower total fat weight, and animals fed a diet supplemented with 3% unextruded BBP in fasting state or 3% unextruded BBP in fed state had lower leptin levels than the control. This is the first study demonstrating the beneficial effects of feeding blueberry pomace on health.
Introduction
G
Blueberry pomace (BBP) is a by-product of the blueberry processing industry. It consists of skin, seeds, stems, and pulp that remain after the fruit has been processed for juice and may account for up to 20% of the initial fruit weight. Because BBP is usually considered a waste material, a substantial amount, up to 50%, of polyphenolics is wasted since the polyphenolics are concentrated more in the skin and seeds of the fruit that are retained in the pomace. While the positive health effects of whole fruits, fruit juice, and other fruit products have been investigated for some time, research on fruit by-products, such as the pomace, for their potential health benefits has only begun.
Extrusion is one of the most common industrial processes used to make snacks, and its main goal is to enhance the overall digestibility and bioavailability of food nutrients. Previously, we demonstrated that extrusion processing of BBP, cranberry pomace, grape pomace, and grape seeds can enhance the biologically important small-molecular-weight procyanidins. 8,9 Extrusion has also been found to be effective in enhancing the total flavonols in cranberry pomace. 9 While its effect on total anthocyanin contents is deleterious, it is not as extensive when extruded at higher feed moisture content 8 as has been observed previously. 10,11 We have recently reported the beneficial effects of feeding cranberry pomace on selected biochemical profiles associated with metabolic syndrome in growing male Sprague–Dawley rats. 12 Using a similar in vivo model, here we report another study about the protective effects of feeding BBP on selected metabolic profiles associated with metabolic syndrome.
Materials and Methods
Chemicals
All chemicals used in the current study were high-performance liquid chromatography grade or higher and were obtained from Fisher Scientific (Hampton, NH, USA), SigmaAldrich (St. Louis, MO, USA), or SynerMed (Monterey Park, CA, USA). 6-Hydroxy-2,5,7,8-tetramethyl-2-carboxylic acid (Trolox) and 2-(6-hydroxy-3-oxo-3H-xanthen-9-yl)benzoic acid (fluorescein) were obtained from SigmaAldrich, while 2,2′-azobis(2-amidinopropane) dihydrochloride was from Wako Chemicals USA, Inc. (Richmond, VA, USA).
Extrusion of BBP
BBP consisting of stems, skins and seeds was obtained from FruitSmart (Prosser, WA, USA). The pomace was freeze-dried in a Virtis (Gardiner, NY, USA) Genesis freeze-dryer before grinding in a coffee grinder. It was mixed with cornstarch in a kitchen mixer (Classic Model K45SS; Kitchen Aid, Benton Harbor, MI, USA) at a ratio of 30:70 on a dry weight basis (a total of 2 kg/batch). Water was added to the mixture to bring the calculated moisture content to 30%. The speed of the kitchen mixture was set at 6. The mixture was then extruded using a twin-screw Haake PolyLab System extruder (Thermo Haake, Karlsruhe, Germany), the details of which have been published previously. 8
Animals and diet
The protocol was approved by the Animal Care and Use Committee of the University of Arkansas for Medical Sciences, Little Rock, AR, USA. Thirty-six male Sprague–Dawley rats (Charles River Laboratories International Inc., Wilmington, MA, USA) (46 days old, weighing 169.2±1.2 g) were used in a randomized complete block design with initial body weight used as the blocking factor. Animals within blocks were assigned at random to one of the six dietary (Table 1) treatments: (1) control (modified AIN-based diet); (2) high fructose (HF) diet (AIN-based diet containing 58% by weight of fructose and 0 kcal of starch); (3) HF diet with 1.5% unextruded BBP (UBBP); (4) HF diet with 1.5% extruded BBP (EBBP); (5) HF with 3% UBBP; and (6) HF diet with 3% EBBP. Animals were housed individually in cages with a 12-h day/night cycle and had ad libitum access to food and water. Food consumption and body weights were monitored weekly. Cages were changed weekly. An 8-day adaptation to dietary and environmental conditions was provided to animals followed by an 8-week experimental period. Animal weight at the end of the adaptation period is presented as the initial weight.
tert-BHQ, tert-butylhydroquinone; EBBP, extruded blueberry pomace; HF, high fructose; UBBP, unextruded blueberry pomace.
Sample collection and laboratory analysis
Samples of both EBBP and UBBP were collected and analyzed for their polyphenolic composition. Details about the individual laboratory procedures and instrumentation have been published for procyanidins, 8 anthocyanins, 13 and phenolic acids. 14
Fasting trunk blood was collected at the time of sacrifice, while postprandial blood samples were collected from the tail vein in Weeks 1, 3, 5, and 7 using a Microvette® capillary tube (Sarstedt AG & Co., Nümbrecht, Germany). Animals were sacrificed on Days 57, 58, and 59 of the experiment by euthanization in a CO2 chamber and decapitation. Kidney, heart, liver, and total fat (epididymal and retroperitoneal) weights were recorded at the time of sacrifice. Animals were randomized before blood collection and sacrifice such that treatments were evenly distributed across the duration. Plasma was separated from the blood by centrifugation at a force of 3480 g for 15 min at 4°C and analyzed for glucose, cholesterol, triglycerides, leptin, insulin, and antioxidant capacity. Plasma glucose, cholesterol, and triglycerides were analyzed using commercially available kits (Synermed International Inc., Westfield, IN, USA) in a 96-well plate format using a dual-pump FLUOstar Optima (BMG Labtech, Durham, NC, USA) microplate reader. Plasma insulin and leptin were determined by enzyme-linked immunosorbent assay in a 96-well plate format using commercially available kits (Millipore, Billerica, MA, USA) in a Benchmark Plus microplate spectrophotometer (Bio-Rad Laboratories, Hercules, CA, USA). Antioxidant status was determined using the fluorescence-based oxygen radical absorbance capacity (ORAC) assay 15 in fasting plasma samples obtained at the time of sacrifice. Plasma samples were diluted 800-, 50-, and 10-fold with phosphate buffer (pH 7.0) for total, hydrophilic, and lipophilic ORAC, respectively, in a 48-well plate format in triplicates for the samples and blanks and quadruplicates for the standards. In total, 35 cycles were performed for each analysis. Results were determined by differences in the area under the fluorescein decay curves among blanks, samples, and standards and expressed as micromoles of Trolox equivalents. All samples were stored at −70°C prior to analyses.
Oral glucose tolerance test
At the end of Week 6, the oral glucose tolerance test (OGTT) was performed over a period of 3 days. Rats were deprived of food for at least 12 h before administration of an oral glucose load of 2 g/kg of body weight from a 200 g/L solution. Blood samples (300 μL) were collected via the tail vein at time 0 (before administration), 30, 60, 90, 120, 150, and 180 min and analyzed for plasma glucose. Animals were randomized before the OGTT such that the same number of animals per treatment was included each day.
Insulin sensitivity indices
The homeostasis model assessment (HOMA) for insulin resistance (HOMA-IR) and that for β-cell function (HOMA-BCF) were calculated as described previously: 16 HOMA-IR=(fasting glucose [in mmol/L]×fasting insulin [in μIU/mL]/22.5 and HOMA-BCF=(20×serum insulin [in mU/L])/(plasma glucose [in mmol/L] – 3.5).
Statistical analysis
Statistical analyses of weekly food intake, weight gain, and postprandial plasma glucose, cholesterol, and triglycerides were carried out in SAS (SAS Institute, Cary, NC, USA) using PROC MIXED. Treatment, block, week, treatment×block, and treatment×week were included in the model as the fixed factors with week as the repeated measure on rats. However, block and treatment×block were removed from the final model owing to their nonsignificant effect on the response variables. Changes in serum glucose concentration after the OGTT were assessed in PROC MIXED of SAS using treatment, time, and their interaction as the fixed factors with time as the repeated measure on rats. Compound symmetry was used as the covariance structure. Fasting glucose, cholesterol, triglycerides, leptin, insulin, HOMA-IR, HOMA-BCF, final body weight, organ weights, organ weights as a percentage of body weight, and total, hydrophilic, and lipophilic ORAC were analyzed by two-way analysis of variance using SigmaPlot (Systat Software Inc., San Jose, CA, USA). Postprandial plasma insulin and leptin were also analyzed using the same model. Treatment, block, and their interactions were included in the model as the fixed factors.
Results
Polyphenolic content, diet composition, diet consumption, weight gain, and organ weights
Contents of selected polyphenolics present in BBP are presented in Table 2. Extrusion of BBP increased the lower-molecular-weight procyanidins considerably. Monomeric procyanidins increased by more than 100% after extrusion, with similar increases observed for dimeric and trimeric procyanidins. There was also some increase in tetrameric procyandins. The rest of the procyanidins decreased after extrusion, with the most decrease being for polymers. Total anthocyanins were reduced by about 40%. Extrusion also reduced the contents of most phenolic acids, except ferulic acid and 3,4-dihydroxybenzoic acid. No other polyphenolics were analyzed.
DP, degree of polymerization; ND, not detected.
All ingredients of the diet, except BBP, were purified. Diets were isocaloric (3834±13.5 kcal/kg) and isonitrogenous (17.9% crude protein). Diets were also balanced for crude fat (4.5%), crude fiber (4.0%), and other macro- and micronutrients (Table 1). Initial weight was similar for all the animals across treatments, and no significant effect (P>.10) of feeding HF or BBP was observed on final body weight (Table 3). Similarly, weekly body weight, weekly diet intake, or dietary efficiency was similar across treatments (Fig. 1). There was no significant dietary effect (P>.10) on heart, liver, and kidney weights (Table 3). Although there was no increase in total fat weight due to HF feeding compared with control diet, a slight reduction in total fat weight occurred with BBP and even more so when EBBP was included in the diet. When organ weights were expressed as a percentage of body weight, no significant effect (P>.10) was observed in heart and liver weights. However, kidney weight as a percentage of body weight showed a slight increase with HF feeding in all animals, including those fed BBP, and a slight reduction in fat weight as a percentage of body weight was found in animals fed EBBP.
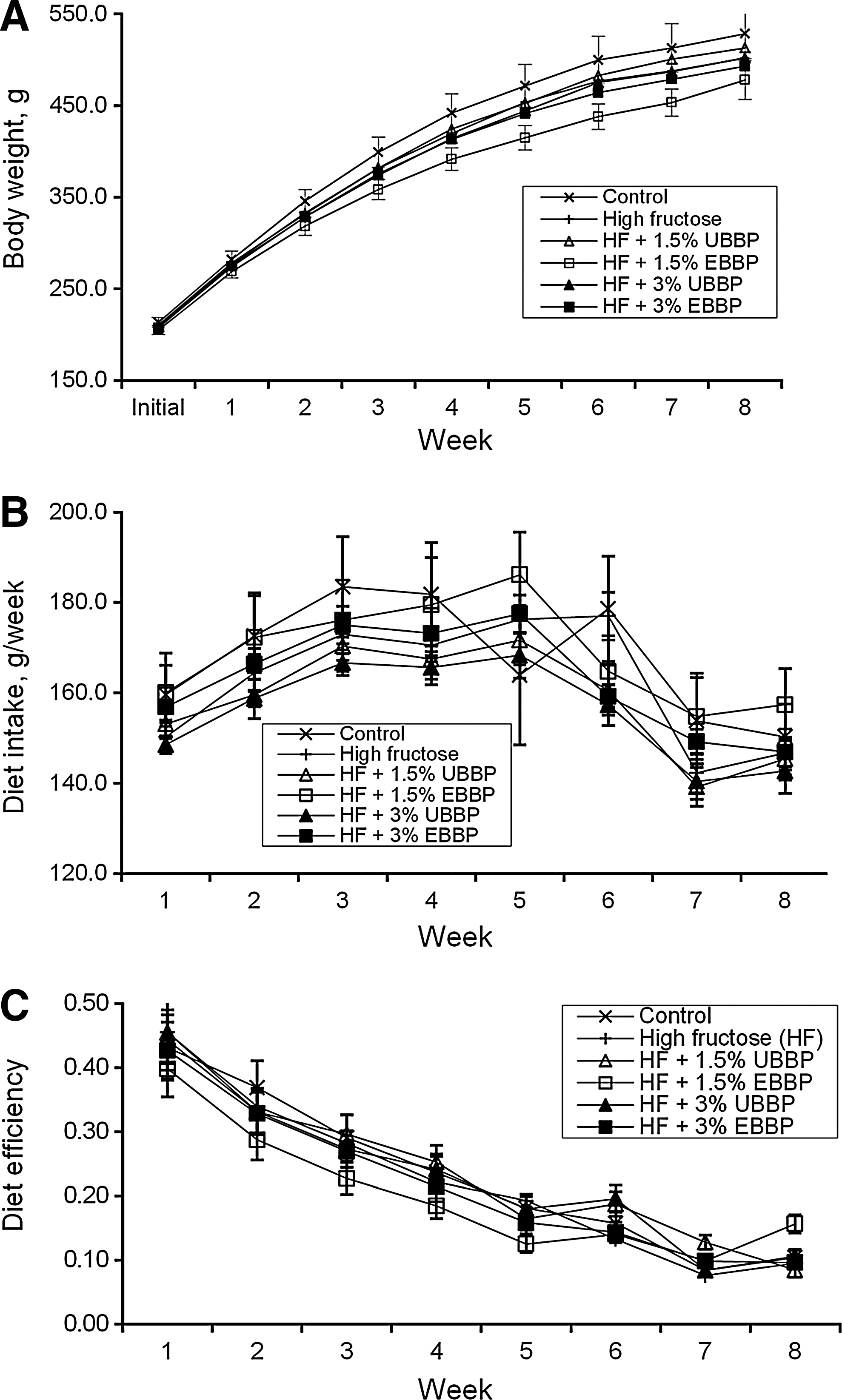
This is the weight used for randomization, which is different from the Week 1 weight at the beginning of the measurement period that occurred after 10 days of adaptation to diet and management conditions.
SEM, standard error of the mean.
Plasma metabolic profiles
Fasting plasma glucose, cholesterol, and triglyceride levels in animals fed different diets are presented in Table 4. There was no significant (P>.1) change in fasting plasma glucose levels across treatments, and all animals had normal fasting plasma glucose levels. However, HF diet significantly increased (P<.05) fasting plasma triglycerides, which was prevented by the inclusion of 3%, but not 1.5%, BBP, with no difference due to extrusion. There was no statistical difference (P>.1) in fasting plasma cholesterol level due to HF feeding compared with the control group, but animals fed 3% BBP had significantly lower fasting plasma cholesterol (P=.05) compared with those on HF diet.
Values with different letters were significantly different (P<.1)
Postprandial plasma glucose (Fig. 2A) and cholesterol (Fig. 2B) levels were within the range for the fed state; neither compound was affected by dietary treatments. However, compared with animals fed the control diet, postprandial plasma triglycerides (Fig. 2C) were considerably higher when HF was included in the diet with no significant effect (P>.1) due to BBP.
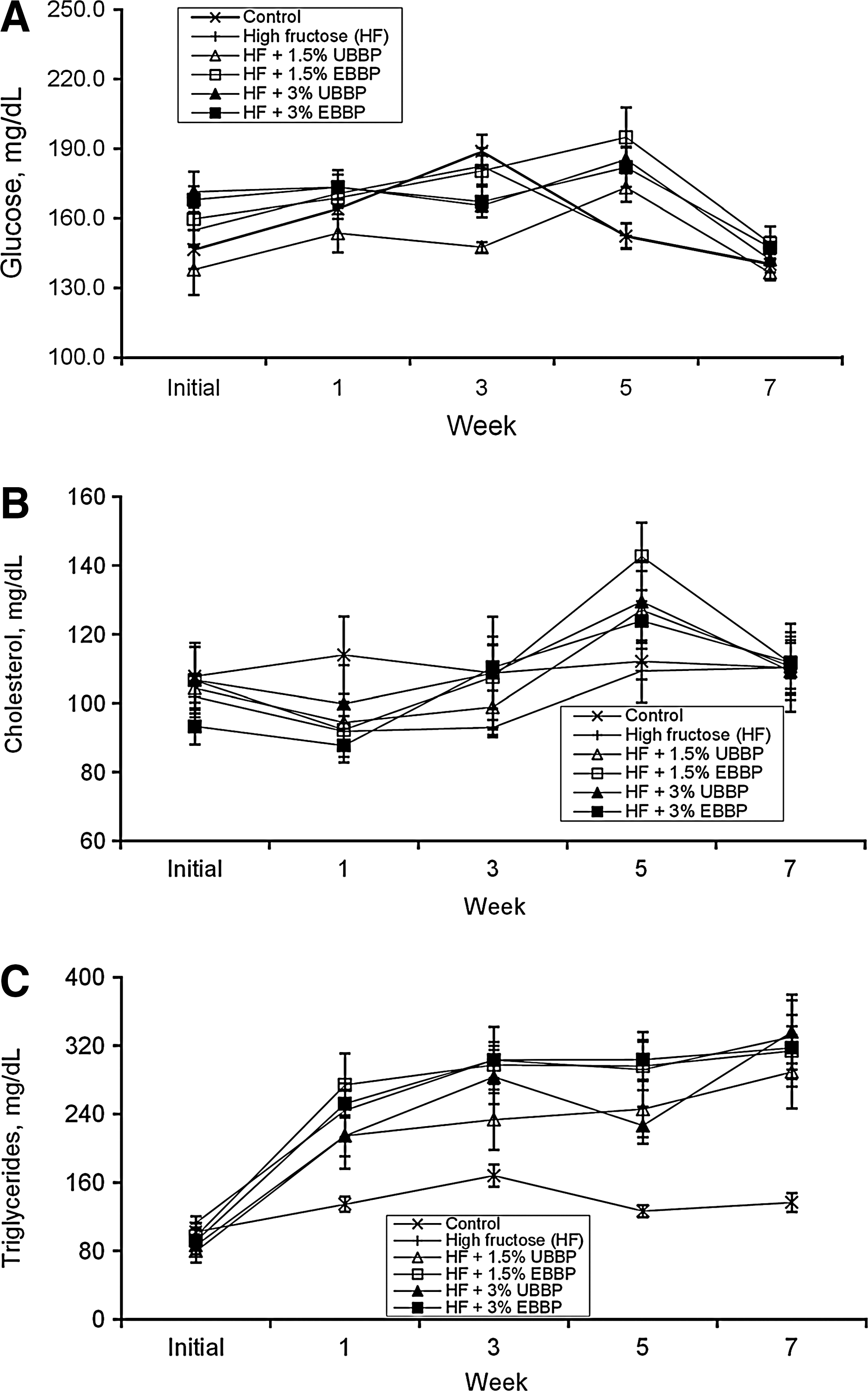
Postprandial plasma
No significant dietary effect (P>.1) was observed on postprandial plasma insulin levels (Fig. 3). However, fasting plasma insulin was considerably higher (P<.05) for animals fed HF diet relative to control. Although inclusion of BBP was able to bring down the fasting plasma insulin to values comparable to those in animals fed a control diet, it was more effective when included at 3% of the diet. Extrusion had no effect (P>.1) on fasting plasma glucose levels. Similarly, plasma leptin levels were lowest (P<.05) in animals in both the fed and fasted states when BBP was included at 3% of the diet (Fig. 4), while it was highest for animals fed the control diet.

Plasma insulin levels in rats fed a control diet, HF diet, and HF diet with 1.5% and 3% UBPP and EBBP:
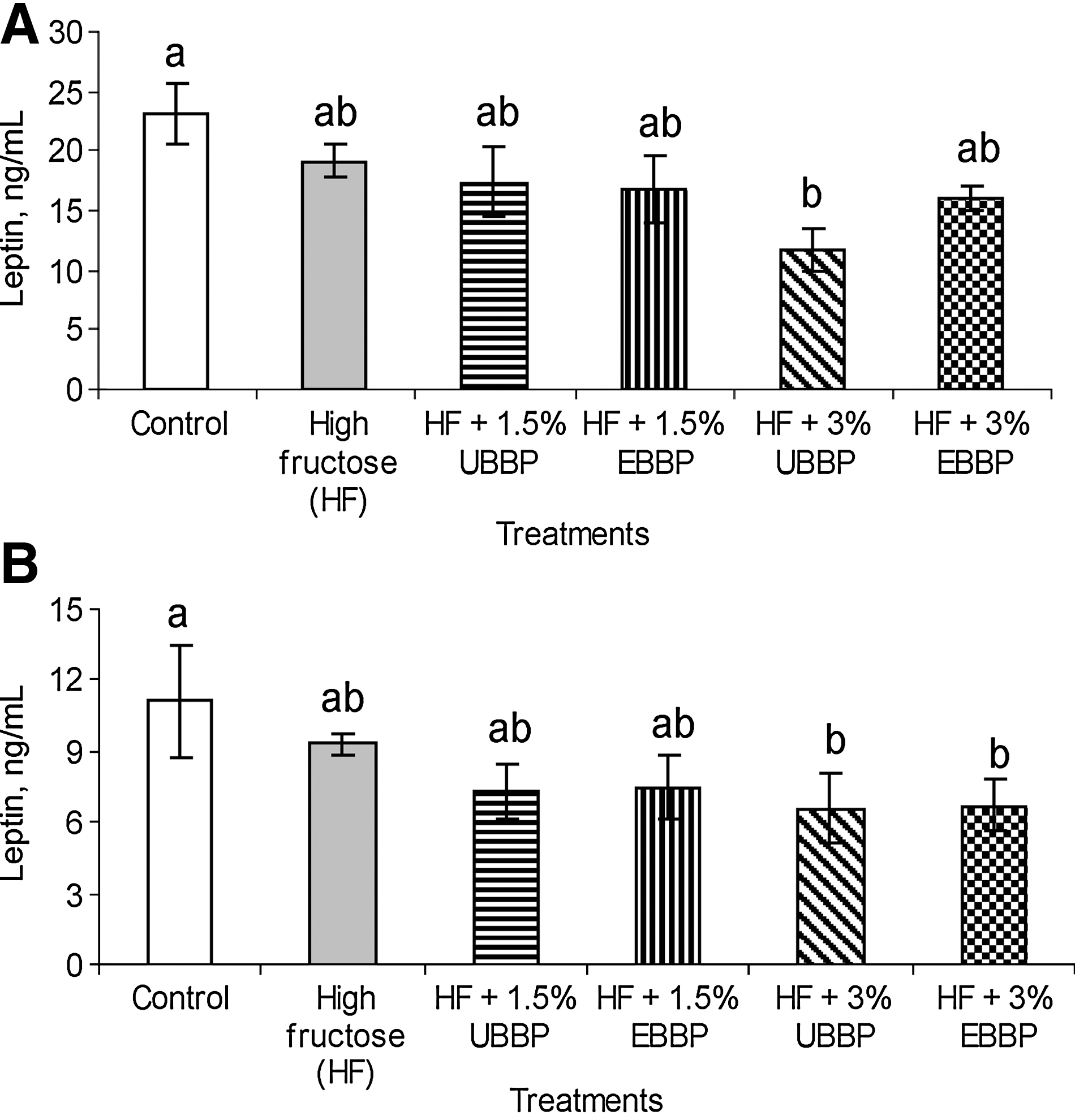
Plasma leptin levels in rats fed a control diet, HF diet, and HF diet with 1.5% and 3% UBPP and EBBP:
Homeostatic assessment scores and OGTT
Compared with animals in control diet, HOMA-IR and HOMA-BCF were affected in animals fed HF diet (Fig. 5). The HOMA-IR was reduced to control levels when BBP was included at 3% of the diet, but not when it was included at 1.5% of the diet. However, there was no dietary effect (P>.1) on the OGTT as evidenced by similar fasting plasma glucose level or area under the curve after the oral glucose load (Fig. 6). There was no significant (P>.1) improvement in HOMA-IR when EBBP was compared with UBBP.
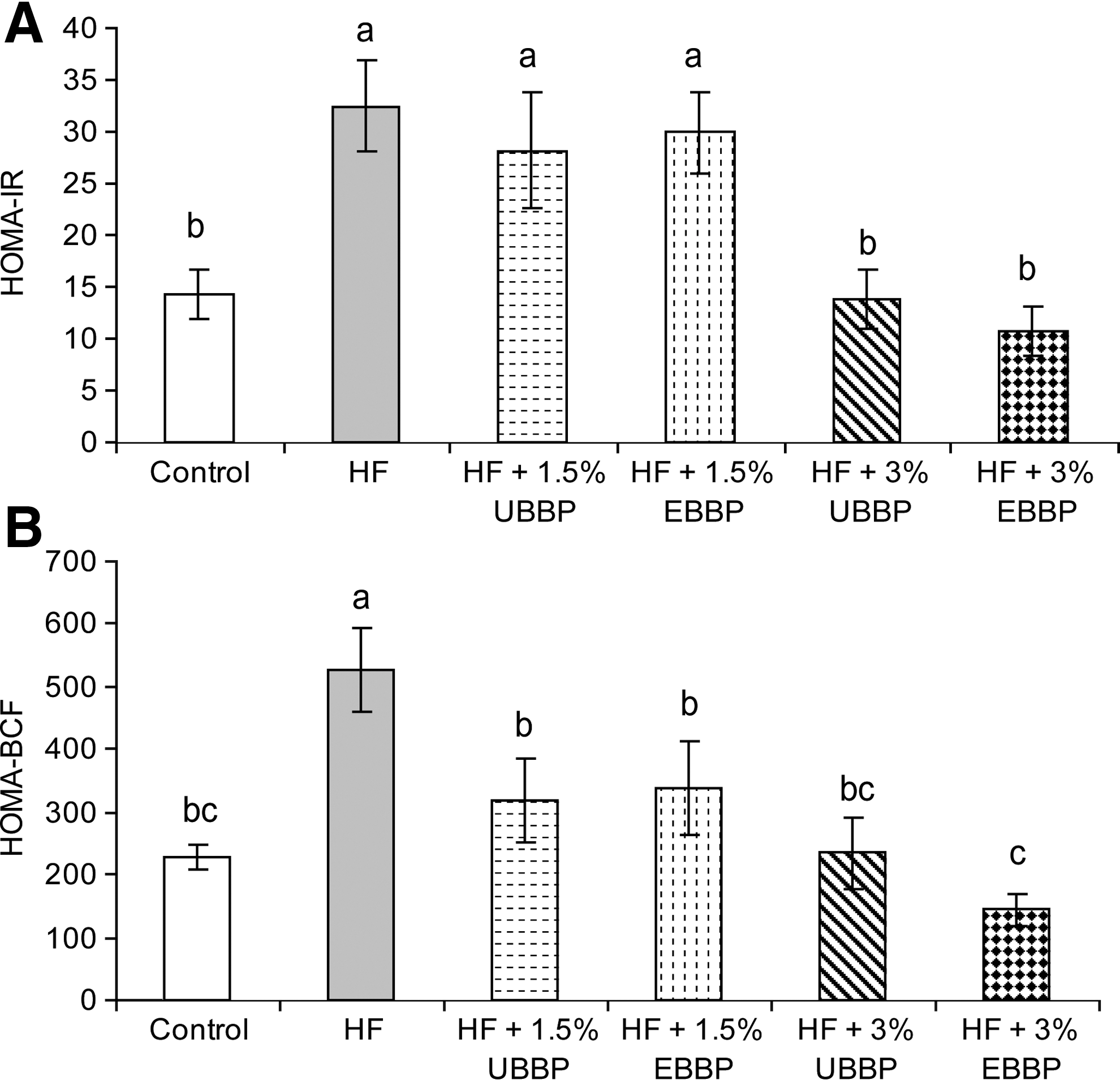
Homeostasis model assessment for
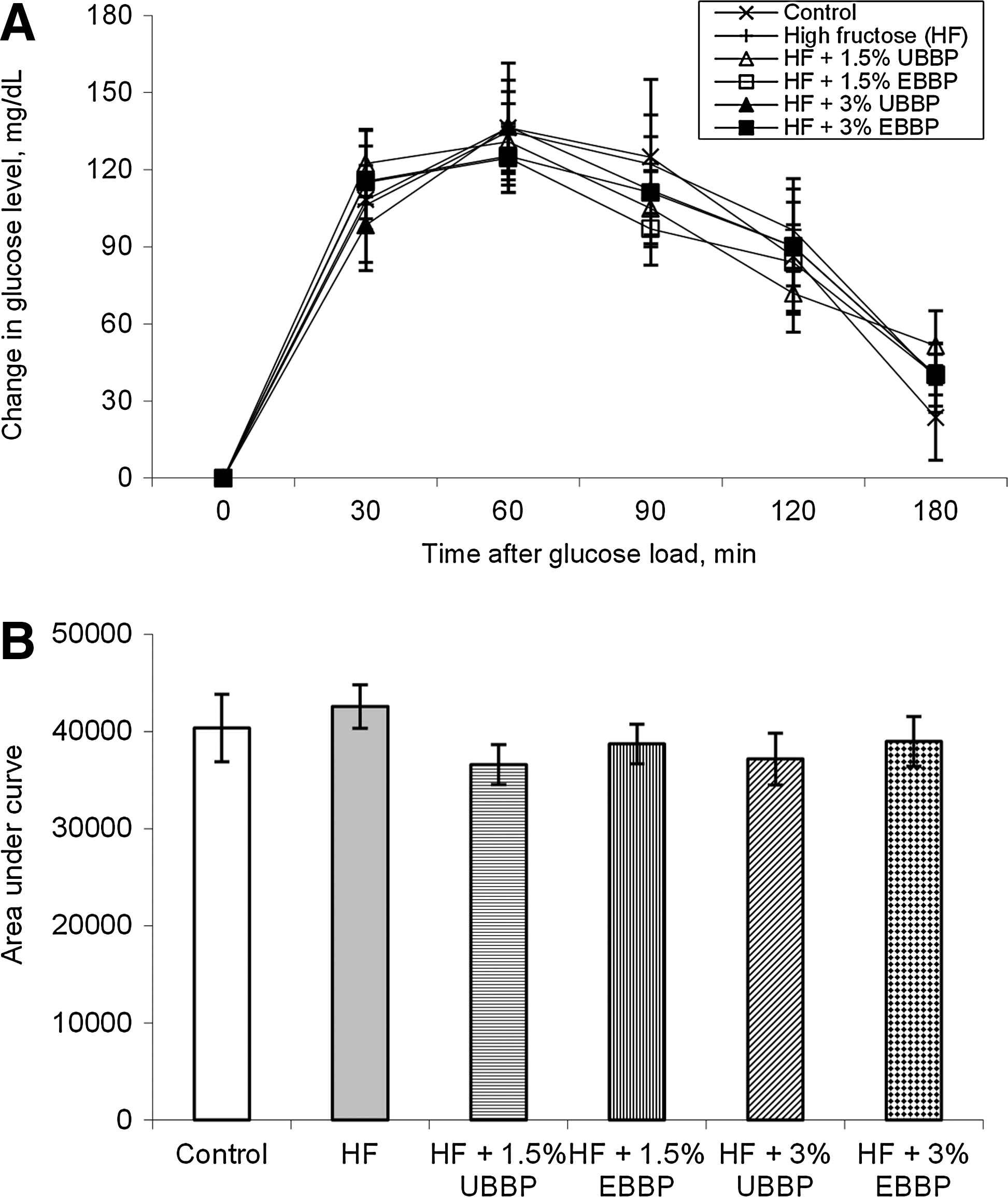
Change in
Plasma antioxidant status
Animals receiving HF diet had a fasting plasma antioxidant status similar to that of their counterparts receiving control diet (Fig. 7). Fasting plasma antioxidant status was numerically higher for animals fed BBP, but it was not significantly different among diets (P>.1).
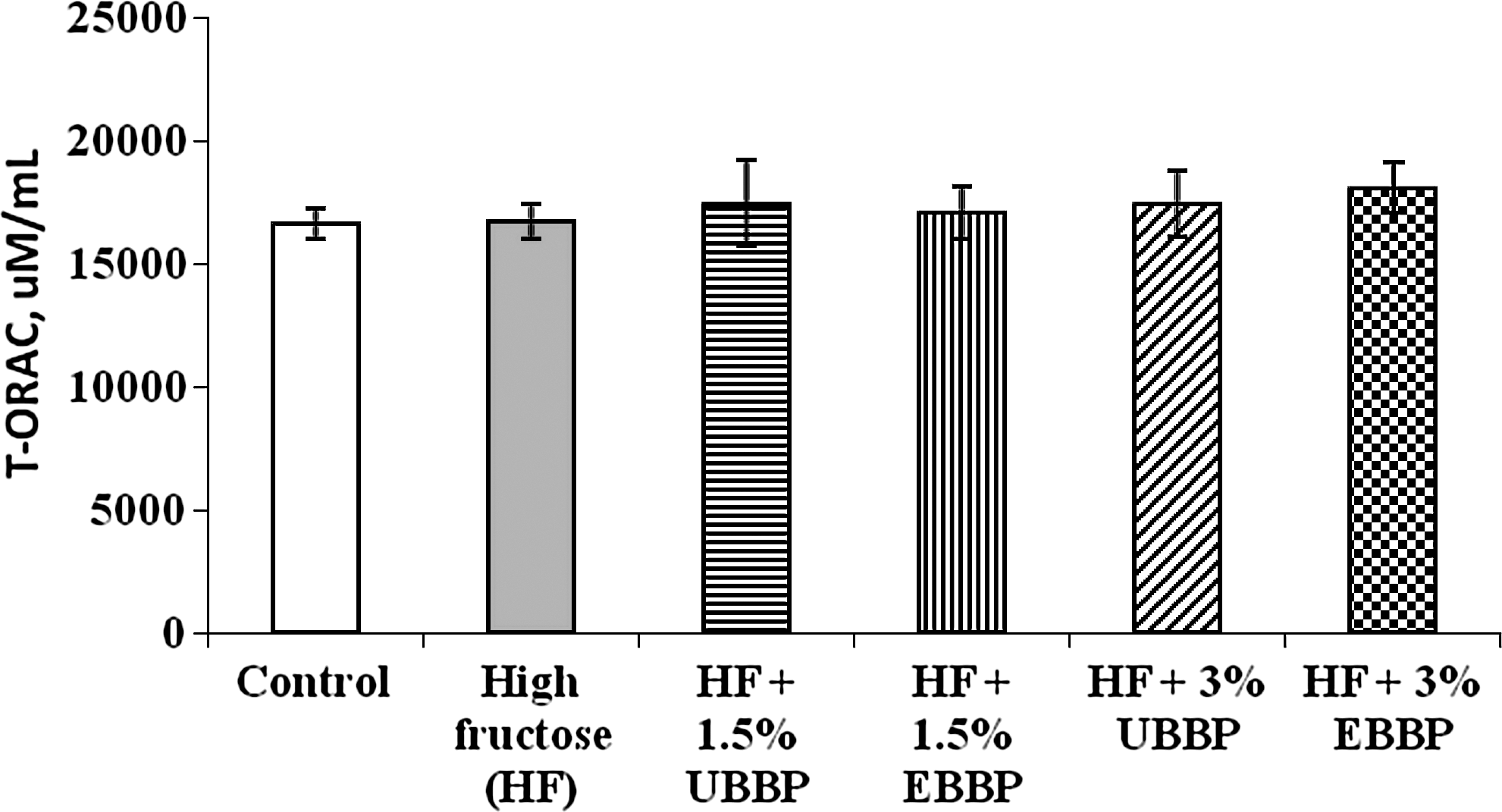
Total fasting plasma antioxidant status as the oxygen radical absorbance capacity (T-ORAC) (in μM of Trolox equivalents/mL) in rats fed a control diet, HF diet, and HF diet with 1.5% and 3% UBPP and EBBP.
Discussion
We have recently reported several studies demonstrating an increase in the relative quantities of monomeric and lower-molecular-weight procyanidins, up to tetramers, after extrusion of grape seed and grape pomace, 8 cranberry pomace, 9 and BBP. 17 This was further confirmed in the current study with an increase of >103%, 110%, and 90% in the monomeric, dimeric, and trimeric procyandins when BBP was extruded under similar conditions using the same extruder. Anthocyanin content is reduced considerably after heat treatment, including extrusion, and this was no exception in the current study. The increase in ferulic acid after extrusion may have resulted from ruptured matrix of the cell wall where it is present in higher amounts, whereas that of 3,4-dihydroxybenzoic acid may be the result of some degradation of catechins during the conversion of polymers or higher-order oligomers into monomers.
Metabolic syndrome is an epidemic of global importance directly related to diabetes 18,19 and cardiovascular disease 20,21 and more recently to cancer 22,23 and non-alcoholic fatty liver disease. 24 –27 Investigating dietary means to minimize the impact or prevent its occurrence is a critical challenge. Numerous health benefits, primarily against chronic diseases, have been attributed to different fruits and fruit products owing to their rich polyphenolic contents. While blueberry is one such fruit that has been investigated in depth for its many health benefits, this is the first study demonstrating the protective effects of its pomace on certain metabolic parameters associated with metabolic syndrome.
Feeding HF diet was able to accentuate some of the metabolic parameters, such as fasting and postprandial plasma triglycerides, fasting plasma insulin, and reduced insulin sensitivity, that are associated with metabolic syndrome and other chronic anomalies. HF-induced hypertriglyceridemia, probably a result of impaired removal of triglycerides, 27,28 may be central to the development of increased insulin resistance in diabetes or metabolic syndrome. Previous studies have suggested that relatively mild, short-term elevation of triglycerides is sufficient to produce a marked reduction in peripheral insulin action. 29 Moreover, a significant negative correlation between plasma triglycerides and insulin-mediated glucose uptake has been observed in people with normal glucose tolerance. 30 Demonstration that BBP, primarily at 3% of the diet, is beneficial in bringing at least the fasting—if not the postprandial—plasma triglycerides down to control levels may be helpful in mitigating hypertriglyceridemia to a certain degree. However, compared with cranberry or grape pomace, 12,31 BBP may be less effective in this regard. One possible reason for this difference may lie in the amount and composition of procyanidins present. Relative to BBP, grape pomace contains considerably higher amounts of monomeric and dimeric procyanidins that have greater bioavailability. Similarly, cranberry pomace has A-type procyanidins as opposed to B-type in BBP. Additionally, BBP, particularly at 3%, was effective in reducing the fasting, but not postprandial, plasma cholesterol level relative to control animals. This may potentially impart further health benefits in mitigating some of these chronic anomalies.
Increased fasting plasma insulin in HF-fed animals suggested that higher amounts of insulin were needed to utilize the available glucose circulating in the fasting state, a result demonstrated by increased insulin resistance as measured by HOMA-IR. Moreover, this increased insulin may have resulted from the pancreatic β-cells acting beyond their capacity, which may jeopardize their survival or capacity to produce insulin in the future. While inclusion of BBP in the diet was able to reduce insulin resistance, thereby requiring lower levels of insulin for utilizing similar amounts of circulating glucose, it also reduced the demand on β-cells as evidenced by a decrease in HOMA-BCF. BBP was more effective when included at 3% of the diet. There was no additional benefit of including EBBP in homeostatic assessment scores.
Because there was no change (P>.1) in the plasma antioxidant status as measured by the total ORAC, both HF-initiated accentuation of some of the metabolic profiles or those improved by the inclusion of BBP may not likely be the result of a change in the oxidative status due to HF feeding or improved antioxidant capability due to increased polyphenol consumption. It has been suggested that plant polyphenolic-derived change in plasma antioxidant content may not be more than 5–10% and may be only transient in nature. 32 However, this is in contrast to our previous finding in which inclusion of cranberry pomace at either 1.5% or 3% of the diet showed improved total ORAC in the plasma compared with HF but not control animals. This discrepancy may be due to differences in polyphenolic profile among cranberry pomace and BBP.
HF feeding, however, was not effective in increasing the diet intake, weight gain, and abdominal fat accretion. Although this is in contrast to many previous studies that indicate at least abdominal obesity as one of the primary indicators of metabolic syndrome, 33 –36 other investigators have observed similar results. For instance, no effect of HF feeding on weight gain was observed in one experiment, 37 while no difference in body weight or total fat as a percentage of body composition was observed in another. 29 Some other findings have also failed to demonstrate abdominal obesity or increased weight gain in HF-fed growing Sprague–Dawley or Wistar rats, while still developing other metabolic profiles associated with metabolic syndrome. 38 –41 It has been demonstrated that fructose-induced insulin resistance may be independent of visceral obesity, 41 which was what we observed in the current study. Our own observations in growing rats have been along the same line. 12,31 Moreover, there is no conclusive experimental evidence that dietary fructose actually increases energy intake, 42 the primary factor responsible for weight gain and abdominal adiposity.
The effect of fructose on leptin appears conflicting. A decrease in circulating leptin concentration in response to HF feeding is considered a known phenomenon, 43,44 which usually results in the increased diet intake that was absent in the current study. However, it is also shown to increase circulating leptin in healthy humans without affecting the body weight or body fat. 45 No increase in food intake, body weight, or abdominal obesity was observed in several other experiments where the researchers have used HF-fed growing Wistar or Sprague–Dawley rats. 12,31 Further investigation may be needed to fully explain such discrepancies. Nonetheless, BBP, primarily at 3% of the diet, which prevented increase in plasma leptin levels induced by HF feeding without stimulating food intake, weight gain, or adiposity, may be beneficial in weight management or obesity.
Although the crude fiber content of the experimental diets was balanced across treatments, we did not analyze the individual components, such as soluble and insoluble fibers. As a result, it may not be reasonable to speculate if any of the observed effects on metabolic profiles resulted from the differences in their contents.
In summary, feeding HF diets to growing Sprague–Dawley rats accentuated certain metabolic parameters, such as fasting and postprandial plasma triglycerides, fasting plasma insulin, and reduced insulin sensitivity, that are associated with metabolic syndrome and other chronic anomalies. However, HF diet was not as effective in augmenting other metabolic parameters, such as glucose intolerance, weight gain, abdominal obesity, and fasting glucose associated with metabolic syndrome, as has been suggested in the literature. The current study further confirmed our earlier findings that HF feeding may possibly augment some of the factors associated with metabolic syndrome without stimulating energy intake and/or promoting weight gain and obesity. Inclusion of BBP, particularly at 3% of the diet, was effective in ameliorating metabolic anomalies associated with HF feeding, except postprandial plasma triglycerides. In addition, inclusion of 3% BBP also improved fasting plasma cholesterol, which can potentially bring additional health benefits. Extrusion of BBP did not impart any additional benefits. Earlier studies focused on health benefits of blueberry fruit, juice, powder, or their polyphenolic extracts, but this is the first study demonstrating the beneficial effects of feeding BBP, a by-product of the juice processing industry, on health.
Footnotes
Acknowledgments
We thank FruitSmart, Prosser, WA, for providing blueberry pomace and B.L. White for help with extrusion. This work was funded by an Arkansas Biosciences Institute grant.
Author Disclosure Statement
The authors declare that they have no competing financial interests.