
Abstract
Gut-associated lymphoid tissue maintains mucosal homeostasis by combating pathogens and inducing a state of hyporesponsiveness to food antigens and commensal bacteria. Dietary modulation of the intestinal immune environment represents a novel approach for enhancing protective responses against pathogens and inflammatory diseases. Dietary rice bran consists of bioactive components with disease-fighting properties. Therefore, we conducted a study to determine the effects of whole dietary rice bran intake on mucosal immune responses and beneficial gut microbes. Mice were fed a 10% rice bran diet for 28 days. Serum and fecal samples were collected throughout the study to assess total immunoglobulin A (IgA) concentrations. Tissue samples were collected for cellular immune phenotype analysis, and concentrations of native gut Lactobacillus spp. were enumerated in the fecal samples. We found that dietary rice bran induced an increase in total IgA locally and systemically. In addition, B lymphocytes in the Peyer's patches of mice fed rice bran displayed increased surface IgA expression compared with lymphocytes from control mice. Antigen-presenting cells were also influenced by rice bran, with a significant increase in myeloid dendritic cells residing in the lamina propria and mesenteric lymph nodes. Increased colonization of native Lactobacillus was observed in rice bran–fed mice compared with control mice. These findings suggest that rice bran–induced microbial changes may contribute to enhanced mucosal IgA responses, and we conclude that increased rice bran consumption represents a promising dietary intervention to modulate mucosal immunity for protection against enteric infections and induction of beneficial gut bacteria.
Introduction
R
Prebiotics are defined as nondigestible food ingredients that have a beneficial effect through their selective metabolism in the intestinal tract. 12 Prebiotics promote the growth of beneficial bacteria, commonly referred to as probiotics. 13,14 In addition, the fermentation of prebiotic components via probiotic bacteria elicits the production of short-chain fatty acids. This results in the acidification of the colonic environment, 15 –17 which is detrimental to pathogenic bacteria and advantageous for colonic epithelial cells. Probiotic bacteria tend to be classified as noncolonizing members of the microbiota with the exception of Lactobacillus, found in low concentration in the intestines. 18 The mechanisms by which probiotics benefit the host include strengthening of the mucosal surface, 19 modulating the immune response, and antagonizing pathogens via competition or antimicrobial agents. 20
Changes in the gut microbiota have been shown to significantly affect the mucosal immune system, 21 most notably through the increased production and secretion of immunoglobulin A (IgA). 22,23 For example, mice administered oral Lactobacillus acidophilus were shown to have increased production of IgA by B cells in the Peyer's patches. 24 Also, probiotic bacteria have been associated with the enhancement of innate immunity through increased macrophage recruitment and phagocytic activity, 13,25,26 as well as pro-inflammatory cytokine production. 27
Emerging evidence supports the beneficial effects of single rice bran components, 28 enzyme-treated rice bran, 11 and fermented rice bran 3,10 on the immune system. Although these studies have uncovered important information, whole dietary rice bran demonstrates greater feasibility and may be more practical for public health intervention in the developing and developed world when compared with extract preparations. Given the strong potential for dietary inclusion of whole rice bran as a functional food ingredient for chronic disease control and prevention, 1,7,9,29 studies focused on the effects of dietary rice bran intake on the gut mucosal immune response and the microbiota are warranted. We investigated whether a 10% rice bran intake when compared with a control diet led to changes in cellular and humoral immune responses as well as alterations in the concentration of native gut Lactobacillus spp.
Materials and Methods
Animals and feeding schedule
Specific pathogen-free 4–6-week-old female ICR mice were purchased from Harlan Laboratories (Indianapolis, IN, USA). Specific pathogen-free mice have a normal composition of intestinal bacteria and are devoid of known mouse pathogens. 30 All mice were provided water ad libitum and fed a maintenance diet (AIN-93M; Harlan Teklad, Madison, WI, USA) for 1 week prior to randomization. Mice were placed on either the control diet (AIN-93M) or the 10% rice bran diet for 28 days. The Institutional Animal Care and Use Committee at Colorado State University approved all protocols involving the animal experiments described in this study.
Diet composition
The Neptune rice variety was chosen based on its availability for rice grown in the United States. Control (AIN-93M) and rice bran–containing diets were formulated to match macronutrients across diet groups, and differences in total starch and fiber contents were balanced using purified diet components. The composition of rice bran–containing diet was calculated based on published reports 31,32 that demonstrated chronic disease-fighting activity of rice bran and were stored at –20°C until fed to the mice. Diet formulations are shown in Table 1.
Control diet (AIN-93M) produced by Harlan.
10% rice bran diet produced by Harlan using AIN-93M in addition to rice bran from Neptune rice variety, adjusted to match control diet in total fat, protein, and carbohydrates.
TBHQ, tert-butylhydroquinone.
Sampling procedures
Mice were fed the experimental rice bran diet for 28 days. Fecal and serum samples were collected from all mice on day 0, 4, 7, 14, 18, 21, 25, and 28 following commencement of experimental diets. Mice were bled from the tail vein (50–75 μL per mouse), and serum samples were obtained using serum separator tubes (BD Biosciences, San Diego, CA, USA). Fecal extracts were made by using methods previously described. 33
Quantification of total IgA and Lactobacillus-specific IgA antibody
Total IgA antibody titers were assessed in mouse feces and serum by enzyme-linked immunosorbent assay (ELISA). Nunc-Immuno™ plates (Thermo Scientific, Waltham, MA, USA) were coated overnight at 4°C with purified rat anti-mouse IgA (C10-3; BD) at a concentration of 5 μg/mL in carbonate-bicarbonate buffer (pH 9.6). Nonspecific protein binding sites were blocked with phosphate-buffered saline (PBS) containing 3% bovine serum albumin (Sigma, St. Louis, MO, USA). Dilutions (1% bovine serum albumin) of the samples and standard were applied to the plate (50 μL per well) and incubated for 1.5 hours at room temperature. Plates were incubated with biotin rat anti-mouse IgA (C10-1; BD) at a dilution of 1:1,000 for 1 hour at room temperature. Next, plates were incubated with peroxidase-conjugated streptavidin (Jackson ImmunoResearch, West Grove, PA, USA) at a concentration of 1:1,500 for 20 minutes at room temperature. Plates were developed with 3,3′,5,5′-tetramethylbenzidine (Sigma). The reaction was stopped using 1 N HCl, and the optical densities were read at 450 nm. Purified mouse IgA (eBioscience, San Diego, CA, USA) was used to create the standard curve for the ELISA.
The Lactobacillus IgA ELISA used a similar protocol as the total IgA ELISA. The Lactobacillus spp. used to coat the plate were isolated from fecal samples collected on day 18 following commencement of dietary rice bran. The lactobacilli was expanded in MRS broth prior to undergoing heat-killing as described previously. 34 ELISA plates were coated overnight at 4°C with 108 colony-forming units/mL heat-killed lactobacilli in carbonate-bicarbonate buffer. Nonspecific binding sites were blocked with PBS containing 5% nonfat dried milk. A positive control sample was obtained by vaccinating intraperitoneally an ICR mouse with the heat-killed lactobacilli and collecting serum after 14 days.
Tissue harvest and immune cell preparation
After the 28-day feeding period, mice were killed following anesthetization by intraperitoneal injection of ketamine (100 mg/kg) with xylazine (10 mg/kg). The mesenteric lymph nodes and Peyer's patches were collected, mechanically disrupted through a mesh screen, and treated with NH4Cl. Next, a 10-cm section of the intestine (ileum) was collected, and the intestinal lamina propria lymphocytes were prepared as previously described.
35
The intestinal tissue was disrupted through a mesh screen prior to being run through a density gradient (Optiprep, Accurate Chemical & Scientific Corp., Westbury, NY, USA). All cell preparations were resuspended in complete RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) containing 10% fetal bovine serum (Gemini Bio-Products, West Sacramento, CA, USA), 2 mM
Flow cytometric analysis
Directly conjugated antibodies used for these analyses were purchased from eBioscience or BD Pharmingen (San Jose, CA, USA). The following antibodies were used in various combinations: anti-CD4 (GK1.5), anti-CD8b (H35-17.2), anti-CD5 (53-7.3), anti-CD27 (LG.7F9), anti-CD11c (N418), anti-CD11b (M1/70), anti-CD45R (clone RA3-6B2), and anti-mouse IgA (C10-1). Immunostaining was completed as described previously. 36 Analysis was performed with a Cyan ADP flow cytometer (Beckman Coulter, Fort Collins, CO, USA), and data were analyzed using FlowJo software (Tree Star, Ashland, OR, USA).
Fecal Lactobacillus levels
On day 0, 4, 7, 14, 18, 21, 25, and 28, following commencement of experimental diets, fecal samples were collected from all mice. Approximately five or six fresh fecal pellets (0.1 g) were collected per mouse and rehydrated in 1 mL of sterile PBS for 15 minutes. The samples were vortex-mixed, diluted in PBS, plated on MRS agar (enrichment agar for the genus Lactobacillus), and placed in a 37°C incubator containing 5% CO2 for 48 hours. All colonies grown on MRS agar were confirmed as Lactobacillus spp. using real-time polymerase chain reaction as previously described. 37
Statistical analyses
Statistical analysis was performed using Prism version 5.0 software (GraphPad, La Jolla, CA, USA). Two-tailed nonparametric (Mann–Whitney) t tests were performed for comparisons between the two groups. A repeated-measures (mixed model) two-way analysis of variance with a Bonferroni post hoc test was performed to compare the two groups over time. Differences were considered statistically significant for P<.05 for all comparisons.
Results
Dietary rice bran intake induces local and systemic IgA production
The effect of daily rice bran intake on the mucosal immune system was first investigated in mice for changes in serum, intestinal, and fecal IgA. Control and 10% rice bran diets were fed to mice for 28 days, and the total IgA antibody levels were measured throughout this time period. The concentration of total IgA in the feces of the rice bran–fed mice (Fig. 1A) was significantly increased on day 18 and remained high through day 21 in comparison with mice on the control diet. A similar difference was detected systemically in the serum (Fig. 1B) with the total IgA concentration peaking at day 18.
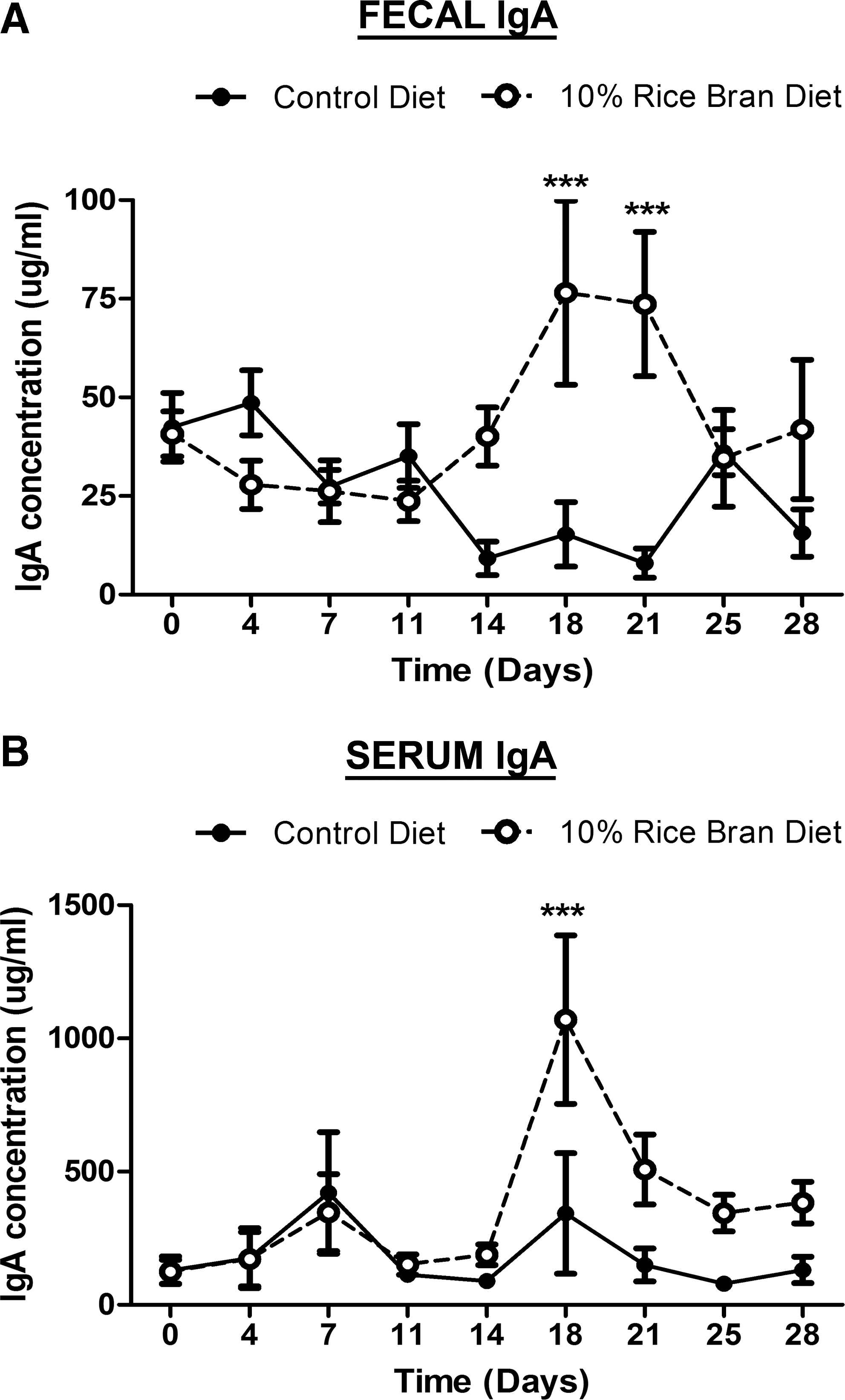
Effect of dietary rice bran on local and systemic total immunoglobulin A (IgA) responses. A total IgA enzyme-linked immunosorbent assay was performed on serial dilutions of
We assessed the expression of IgA on the surface of intestinal B cells to elucidate the modulating IgA response following dietary rice bran intake. Increased expression of surface IgA indicates activation of local B cells as well as an increased potential to respond to commensal and pathogenic bacteria in a T cell–independent fashion. 23 Mice were euthanized after 14 days of rice bran consumption, and the B220+ B cells were analyzed for surface IgA expression (Table 2). A significant difference was observed in the number of IgA molecules expressed on the B cell surface between control and rice bran–fed mice, determined by mean fluorescence intensity. These results are consistent with the idea that dietary rice bran induces mucosal immunity via systemic and local IgA production as well as increased activation of IgA+ B cells found in the Peyer's patches.
Data are mean±SEM values, pooled from two independent experiments (n=10).
P<.05 as determined by comparing control diet versus 10% rice bran (RB) diet in each cell population using a nonparametric Mann–Whitney t test.
MFI, mean fluorescence intensity.
Increased mucosal CD11c+CD11b+ dendritic cells induced by rice bran
In order to determine whether dietary rice bran intake influences antigen-presenting cells as a mechanism for IgA induction, we examined cellular immune phenotypes previously shown to be required for IgA class switching. 38,39 Following 14 days of daily 10% rice bran intake, mice were sacrificed, and the cellular responses were assessed in the lamina propria, Peyer's patches, and mesenteric lymph nodes (Table 3). The percentages of myeloid dendritic cells (CD11c+CD11b+) in both the lamina propria and the mesenteric lymph nodes were significantly increased in mice fed the 10% rice bran diet in comparison with the mice on control diet. No significant differences were detected in T or B cell populations (Table 3). These data suggested that rice bran–mediated induction of dendritic cell recruitment into the mucosal tissues resulted in increased antigen presentation and subsequent IgA production.
Data are mean±SEM values, pooled from two independent experiments (n=10).
P<.05, ** P<.01 as determined by comparing control diet versus 10% RB diet in each cell population using a nonparametric Mann–Whitney t test.
DC, dendritic cell.
Dietary rice bran increased Lactobacillus colonization
Given the associations of probiotic bacteria with the modulation of the mucosal immune system, 21 we next determined the titers of the native gut Lactobacillus spp. in the rice bran–fed mice compared with the control group. Lactobacillus was chosen for its aerobic growth conditions as well as its ability to protect against a variety of infectious diseases. 40 For these experiments, we processed and plated fresh fecal pellets for the presence of Lactobacillus spp. over a 28-day period. On days 11, 18, 21, and 25 the numbers of Lactobacillus spp. in the feces were significantly higher from the rice bran–fed mice in comparison with a more cyclical pattern seen in the mice fed control diet (Fig. 2). The nearly 500% increase in numbers of Lactobacillus spp. (in colony-forming units per gram of feces) became apparent on day 11 and remained significantly high for 14 days. Because of the ability of multiple Lactobacillus species to induce protection against intestinal pathogens, 40 –43 real-time polymerase chain reaction was performed to confirm a greater than 900% increase in DNA levels from the genus Lactobacillus between day 0 and day 14 in the mice fed the rice bran diet (data not shown). Both the control and rice bran diets did not contain any lactobacilli. Next, we determined if the increased IgA titers were Lactobacillus-specific. A Lactobacillus IgA ELISA was performed and revealed low to undetectable Lactobacillus-specific IgA (data not shown). A positive control was in place to ensure the reliability and sensitivity of the Lactobacillus IgA ELISA as described in Materials and Methods. Therefore, these results indicate a potential role for dietary rice bran in modulating the intestinal microbiota through eliciting increased concentrations of beneficial bacteria.
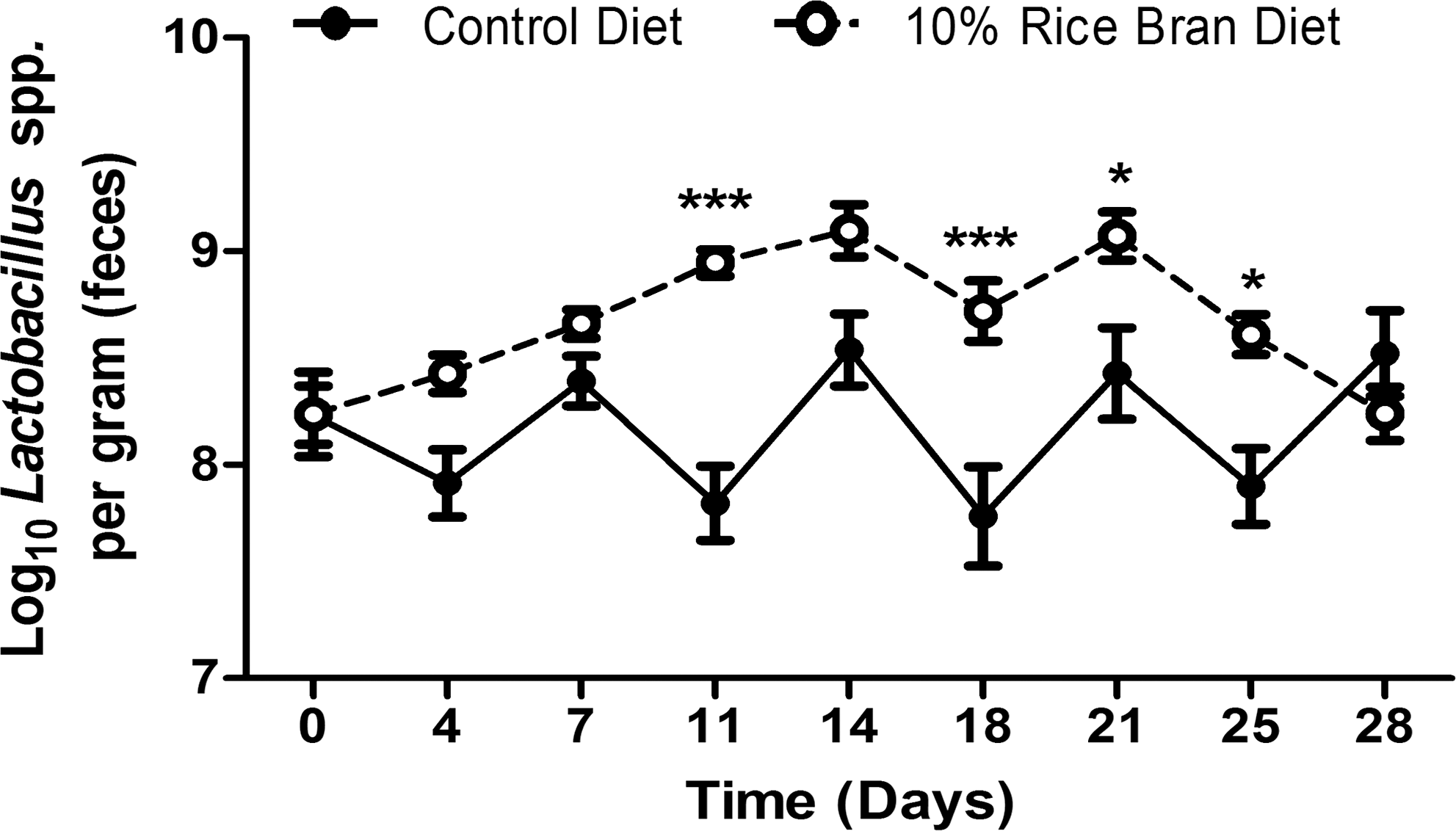
Effect of dietary RB on fecal titers of Lactobacillus. Fresh fecal pellets were collected, processed, and plated on MRS agar every 3–4 days in order to determine the Log10 Lactobacillus titer per gram of feces. Data are mean±SEM values (n=10 mice per group). Results were pooled from two independent experiments. Significant differences (*P<.05, ***P<.001) were determined by a repeated-measures (mixed model) two-way analysis of variance followed by a Bonferroni post hoc test.
Discussion
The unique health-promoting properties and chemical composition of rice bran make it a promising candidate for dietary supplementation, for nutritional therapy, and for prevention of chronic disease. 1,2,4 –9,28,44 –47 In this study we found the effects of whole dietary rice bran intake on the mucosal immune system to be the induction of IgA (Fig. 1) and the enhancement of the innate immune response (Table 3). In addition, the ability of dietary rice bran to promote increased intestinal colonization of native Lactobacillus spp. (Fig. 2) highlights a novel role for rice bran prebiotic components to influence the intestinal microbiota. 2,48
Understanding the effect of dietary rice bran on the innate immune system and antigen presentation is crucial to elucidating the mechanisms by which rice bran may induce protective responses at mucosal surfaces. The increased percentages of myeloid dendritic cells in both the lamina propria and the mesenteric lymph nodes following rice bran consumption speak to the targeted affect of rice bran on the key cells involved in shaping an immune response. Intestinal dendritic cells have the unique ability to induce IgA production from B cells through secretion of retinoic acid, interleukin-5, and interleukin-6. 49 Therefore, the enhanced dendritic cell population can help in the initiation of an adaptive immune response through the activation of B cells and subsequent IgA class-switching. Also, the increased presence of dendritic cells in the lamina propria enhances the mucosal innate immune response at the site where pathogens invade and reveals a potential protective mechanism induced by dietary rice bran.
Evidence for the modulation of the mucosal IgA response associated with the consumption of dietary components has been limited to fructooligosaccharides and pectin. 50,51 Consistent with these findings, this study demonstrated the ability of dietary rice bran to induce increased IgA concentrations systemically and locally. Various bioactive and prebiotic components present in rice bran are hypothesized to be involved in the enhancement of immunity. These phytochemicals include, but are not limited to, γ-oryzanol, polyphenols, and fatty acids, as well as some essential amino acids and micronutrients. 2 Understanding the mechanism of IgA induction is of considerable importance when evaluating the dietary capacity of rice bran to elicit protection against enteric infections. Current evidence suggests that the majority of IgA molecules in the gut are induced by commensal bacteria and that there is a role for this transient population to induce a specific IgA response. 23,52 –57 A recent study performed by Hapfelmeier et al. 54 confirms the role of commensal bacteria, but also describes the specific IgA response to be less robust and have a slower onset, consistent with our findings. Another relevant finding by Macpherson et al. 23 showed induction of commensal-specific IgA antibodies to be mostly T cell independent but dependent largely on the B1 peritoneal B cells, a potential angle that should be evaluated in rice bran–fed mice.
Based on the emerging evidence for commensal involvement in shaping the mucosal IgA population, the beneficial effect of dietary rice bran on the native gut Lactobacillus spp. was shown to be a potential mechanism of immune modulation (Fig. 2). The antibody-enhancing ability of Lactobacillus was described previously in a study showing increased production of rotavirus-specific antibodies when fermented milk was administered during the acute phase of a rotavirus infection. 58 The low Lactobacillus-specific IgA antibodies in our model may speak to the unique ability of rice bran to enhance antibody responses specific for other bacteria, or it may reflect the fact that dietary rice bran contains growth substrates for resident bacteria. As a result, the specificity of the increased IgA would be heterogeneous, making the Lactobacillus titer too low for detection by ELISA. We hypothesize that rice bran–induced modulation of multiple commensal bacteria resulted in increased luminal IgA concentrations in the intestine. Future research directions should include elucidating the effect of dietary rice bran on the gastrointestinal microbiota using 454 pyrosequencing or other high-throughput microbiome analytical approaches.
The ability of dietary rice bran to promote the growth of native gut Lactobacillus spp. and enhance mucosal immune cell populations offers numerous health-promoting and disease-fighting possibilities. For example, the potential use of rice bran as a dietary vaccine adjuvant is highlighted by the recent application of Lactobacillus fermentum in an intramuscular influenza vaccine. 59 Also, the beneficial effects of rice bran on inflammatory diseases such as inflammatory bowel disease 10,11 and type 1 diabetes 6,60 emphasize a unique research avenue to study the immune-enhancing effects of dietary rice bran on these nutritionally relevant diseases. In summary, the ability of whole dietary rice bran to modulate the immune system as well as promote the growth of native gut Lactobacillus spp. holds great promise in protection against enteric pathogens and modulation of chronic inflammatory diseases.
Footnotes
Acknowledgments
We would like to thank Dr. Anna McClung from the U.S. Department of Agriculture rice research unit for providing rice bran from the single Neptune variety. This work was supported by a Grand Explorations in Global Health grant (OPP1015267) from the Bill and Melinda Gates Foundation and the Shipley Foundation and a grant from the National Institutes of Health (R03CA150070).
Author Disclosure Statement
The authors disclose no conflicts of interest.