
Abstract
Consumption of polyphenol-rich fruits, for example, tart cherries, is associated with a lower risk of cardiovascular disease and cancer. This is due, in large part, to the diverse myriad bioactive agents, that is, polyphenol anthocyanins, present in fruits. Anthocyanin-rich tart cherries purportedly modulate numerous cellular processes associated with oncogenesis such as apoptosis, cellular proliferation (CP), and cell cycle progression, although the effective concentrations eliciting these effects are unclear. We hypothesized that several dose-dependent effects over a large concentration range of 100% tart cherry juice (TCJ) would exist and affect these processes differentially with the potential for cellular protection and cellular death either by apoptosis or by necrosis. In this in vitro study, we tested the dose response of TCJ on CP and cell death in MCF-7 human breast cancer cells. TCJ was added at 0.03–30% (v/v) to cells and incubated overnight with the medium alone or with increasing TCJ. Bromodeoxyuridine incorporation was significantly reduced by 20% at ≥10% (v/v) TCJ and associated with necrosis, but was not different between the control and treatment groups at <10% TCJ. MTT reduction was also significantly reduced by 27% and 80% at 10% and 30% TCJ, respectively, and associated with necrosis. Apoptosis, but not necrosis, was increased ∼63% at 3% TCJ (∼307 nM monomeric anthocyanins), yet significantly decreased (P<.05) by 20% at 1% TCJ (920 nM) both of which were physiologically relevant concentrations of anthocyanins. The data support a biphasic effect on apoptosis and no effect on proliferation.
Introduction
D
Anthocyanins are the most abundant flavonoid polyphenols in plants with >300 different anthocyanidins (aglycone) and their glycosides (aglycone plus sugar) occurring in fruits such as berries, grapes, and cherries. 4,5 These water-soluble plant pigments confer intense coloration to fruits and flowers and are especially rich in tart cherries with up to 400 mg per kg fresh weight. 1,4,6 Although dietary intake of anthocyanins is significant and estimated to be 180–215 mg/day in the United States, numerous dietary supplements have been manufactured from tart cherries and marketed, with widely varying doses, based on accumulating evidence suggesting efficacy against inflammatory conditions such as arthritis and gout, muscle and back pain, diabetes, neurodegenerative diseases, and chronic diseases such as cancer. 3,7,8
Tart cherries have been shown to exhibit anticancer activity. Anthocyans (anthocyanidins and anthocyanins) can inhibit malignant cell survival and interrupt secondary cell signaling in oncogenesis. 5 Anthocyans have also been shown to inhibit intestinal tumorigenesis in APCmin mice, a model prone to multiple intestinal neoplasia, with fewer tumors, lower tumor burden, and smaller total tumor volume and area. 9,10 It is postulated that anthocyans exert antioncogenic activity through antiproliferation, induction of apoptosis and/or cytotoxicity, inhibition of oncogenic transcription factors and protein tyrosine kinases, disruption of the cell cycle, reduced mutagenic activation, and deactivation of various enzyme systems, although the doses needed to elicit these beneficial effects are unclear. 11 –13
The objective of these experiments was to identify effective concentration ranges where 100% tart cherry juice (TCJ) would influence the balance between CP and cell death, or apoptosis, in a routinely used in vitro model of cancer, viz, MCF-7 estrogen-responsive human breast cancer cells. We also sought to identify the relative dose needed to elicit effects on the cell number balance without induction of destructive necrosis and compare this to actual attainable plasma concentrations of anthocyanins. We hypothesized that beneficial effects on apoptosis would occur at TCJ concentrations approximating the physiological range of plasma anthocyanins, and that concentrations far exceeding or far below would result in either no effect or adverse effects on cells. The data gleaned from this in vitro dose–response study may contribute to a better understanding of the concentration-dependent effects of absorbed anthocyanins on cellular processes associated with oncogenesis and permit recommendations.
Methods
Chemicals
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), nicotinamide adenine dinucleotide, trypan blue, sodium pyruvate, trypsin, gallic acid, sodium carbonate, sodium pyruvate, potassium chloride, and sodium acetate were obtained from Sigma Chemical Company (St. Louis, MO, USA). Plastics ware (plates and flasks) was purchased from Corning, Inc. (Corning, NY, USA). All other chemicals and cell culture reagents were obtained from Sigma Chemical Company unless otherwise indicated.
Maintenance of cell line
MCF-7 human breast cancer cells were purchased from American Type Culture Collection (ATCC) (Manassas, VA) and cultured in flasks at 37°C in a humidified 5% CO2 atmosphere. Cells were grown in minimum essential medium (MEM) with 1 mM sodium pyruvate, 25 mM sodium bicarbonate, and 10% (v/v) fetal bovine serum. MEM was further supplemented with 0.01 mg/mL bovine insulin, 1% penicillin (100 U/mL), and 1% streptomycin (100 μg/mL; Sigma Chemical Company). Stock flasks of MCF-7 cells were grown to >75% confluency and subcultured routinely by trypsinization. The medium was renewed two to three times weekly.
For experiments, cells were grown in 96-well plates and T25 flasks. At 70% confluency, the medium was removed, and cells were treated with MEM alone or MEM containing increasing concentrations (0.03–30.0% v/v) of filter-sterilized (0.2-μm pore size) TCJ. After incubating for 20 h, the medium was removed, and cells were washed twice with phosphate-buffered saline (PBS), observed, harvested, and assayed as described below.
Preparation of TCJ
TCJ (100%) was obtained from a local supermarket in the greater Phoenix area. The 32-ounce bottle was inverted five times to mix, then a 50-mL aliquot was transferred to a centrifuge tube followed by low-speed centrifugation (1500 g) for 10 min to pellet debris and sediment. The supernatant was transferred to microfuge tubes and centrifuged at high speed (14,000 g) for 10 min to precipitate fine particles. The supernatants were combined and filtered through a 0.22-μm filter disk into a sterile tube. The juice was subsequently diluted into sterile MEM cell culture media using aseptic techniques under standard cell culture conditions to produce concentrations of 0.03–30.0% v/v. Total polyphenols and total monomeric anthocyanins were analyzed on samples before centrifugation and filtration steps.
Analysis of total polyphenols
The Folin-Ciocalteu method was used to quantify total polyphenols in samples of TCJ as described previously. 14 Three milliliters of water, 200 μL Folin-Ciocalteu reagent, and 200 μL of diluted sample or standard (gallic acid) were added to test tubes and mixed well by gentle vortexing (3 sec; low speed). Tubes were incubated at 25°C for 10 min followed by addition of 600 μL of 20% sodium carbonate. Tubes were then incubated in a water bath at 40°C for 20 min, followed by cooling for 30 min to 25°C. Samples were then analyzed at 755 nm using purified gallic acid as the external standard.
Analysis of total monomeric anthocyanins by the pH differential method
Total monomeric anthocyanins in 100% TCJ were quantified by the pH differential method. 15 Briefly, aliquots of TCJ were diluted 1:10 into both 0.025 M potassium chloride solution (pH 1.0) and 0.4 M sodium acetate buffer (pH 4.5). After equilibrating for 15 min at 25°C, all samples were measured at both 510 and 700 nm. Total anthocyanins were expressed as cyanidin-3-glucoside equivalents per liter using a molar absorptivity of 26,900 L/mol and a molecular weight of 449.2 g/mol.
HPLC analysis of anthocyanin profile in TCJ
The anthocyanin profile of TCJ was analyzed by the HPLC method of Spanos and Wrolstad. 16 Briefly, single-strength TCJ (4 mL) was passed through an activated C18 cartridge (Sep-Pak; Waters, Milford, MA, USA), followed by 0.01% HCl (5 mL). Pigments were eluted with acidified methanol (3 mL) and then passed through a 0.45-μm filter disk. The anthocyanin pigment profile was determined by binary-gradient HPLC using a 5-μm (2.5-cm×4.6-mm) C-18 column and a flow rate of 1.5 mL/min with detection at 515 nm. The mobile phases were (A) formic acid:water (90:10) and (B) formic acid:methanol:water (10:50:40). Profiles from samples (shown in Fig. 1) were compared with authentic pigment profiles of tart cherries, and purified cyanidin-3-glucoside was used to generate a reference chromatogram in accordance with authentication protocols used commercially. The most frequent anthocyanins in tart cherries are cyanidin 3-glucosylrutinoside, cyanidin 3-glucoside, cyanidin 3-sophoroside, and cyanidin 3-rutinoside. 17
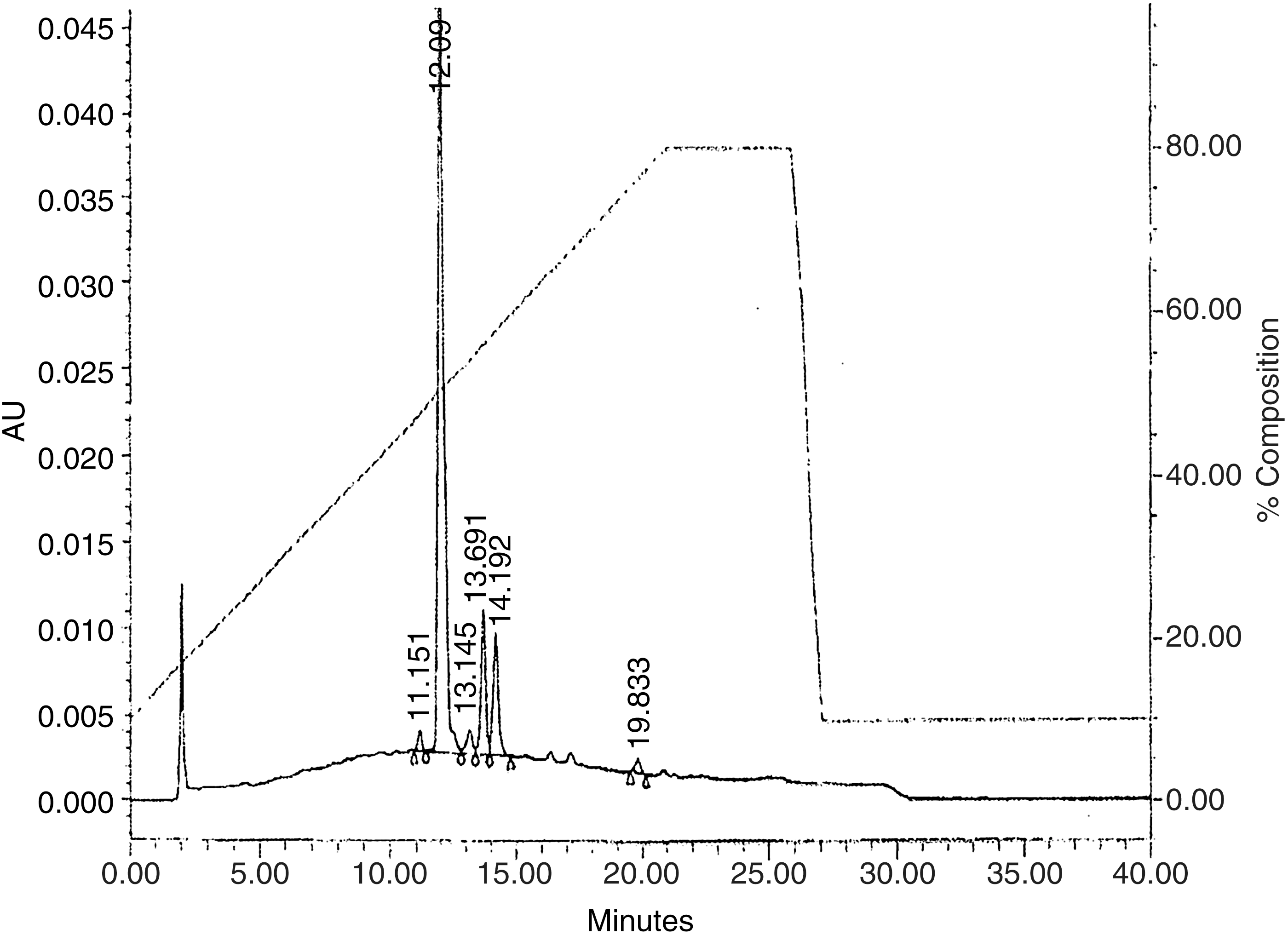
HPLC chromatographic profile of tart cherry juice (TCJ) anthocyanins. Authenticity of commercially obtained 100% TCJ was verified by the peak of cyanidin-3-2G-glucosylrutinoside at 12.008 min, and anthocyanin profile shown in remaining peaks as follows: cyanidin-3-glucoside, 13.516 min; cyanidin-3-rutinoside, 13.985 min; peonidin-3-rutinoside, 16.946 min; acylated anthocyanin, cyanidin aglycone, or a nonanthocyanin compound, 19.595 min. The right y-axis indicates relative percentages of the binary gradient used for separation. The mobile phases were formic acid:water (90:10) and formic acid:methanol:water (10:50:40).
Necrosis analyzed by lactate dehydrogenase release
We opted to use single-strength TCJ in our experiments instead of an extract, and as a result, corrected for the effects of the water vehicle by measuring the cytoplasmic release of lactate dehydrogenase (LDH) to the cell culture supernatant. We incubated MCF-7 cells for 20 h in 96-well plates with medium alone or with increasing concentrations of water (0.03–30%). Monolayers were processed, and LDH was analyzed as described previously. 18 LDH activity in medium and cells was determined and expressed as % release=([LDH activity in media]/[LDH activity in media+LDH activity in cells])×100.
Mitochondrial activity by the MTT assay
Monolayers were treated as described above, then washed, and a phenol red-free medium with MTT (5 mg/mL) was added. Cultures were incubated for 4 h at 37°C, with subsequent centrifugation of the plate for 10 min (1500 g at 25°C) to pellet-reduced MTT. The medium was aspirated; isopropyl alcohol (0.1 mL per well) was added; samples were mixed (15 min at 25°C) to solubilize MTT, and solutions were analyzed at 560 nm.
CP by bromodeoxyuridine incorporation
CP was determined by both a bromodeoxyuridine (BrdU) incorporation assay and hemocytometry (cell counts). For the former, stock BrdU was diluted 1:2000 in MEM, and added to supernatant in wells of control and treated cells 2 h before harvest. After washing monolayers and fixing cells with 37% buffered formalin, an enzyme-linked immunosorbent assay was performed as described by the manufacturer (Trevigen, Gaithersburg, MD, USA) to measure DNA incorporation of BrdU. After addition of stop solution (2 normality [N] HCl), samples were analyzed at 450 nm by spectroscopy.
Cell number and viability by hemocytometry and trypan blue exclusion
After overnight incubation with MEM alone or containing TCJ, monolayers were washed twice with PBS, trypsinized, transferred to culture tubes, centrifuged, and resuspended in fresh medium. An aliquot (50 μL) of cell suspension was added to an equal volume of trypan blue, and cells were counted by light microscopy using a hemocytometer. Dead cells, but not live cells, stained intensely with trypan blue and indicated cytotoxicity. Viability was determined using the following formula: % viability=[(live cells)/(dead cells+live cells)]×100.
Apoptosis by terminal deoxynucleotidyl end labeling
MCF-7 cells were incubated for 20 h as described above. Apoptosis was determined by the terminal deoxynucleotidyl end-labeling (TUNEL) method as described by the manufacturer (Trevigen). Briefly, after treatments, cells were pelleted by low-speed centrifugation, fixed in 37% buffered formalin, and permeabilized. Endogenous peroxidase activity was quenched using 3% hydrogen peroxide in methanol, and cells were incubated for 1 h at 37°C with a labeling reaction mix (terminal deoxynucleotidyl transferase, Mn2+, and biotinylated dNTPs), followed by streptavidin horseradish peroxidase. Apoptosis was determined by color development using TACS-Sapphire substrate (Trevigen) and measured at 450 nm after addition of stop reagent (2 N HCl).
Agarose gel electrophoresis for DNA banding
Apoptosis was also analyzed by DNA fragmentation, or DNA banding, using agarose gel electrophoresis. DNA was extracted from control and treated cells using a DNA miniprep kit according to the manufacturer's instructions (Qiagen, Valencia, CA, USA). DNA (10 μg/well) was loaded onto a 1.2% agarose gel and electrophoresed for 1 h at 85 V using TBE-running buffer (89 mM boric acid, 89 mM Tris base, and 2 mM EDTA). DNA was visualized by ethidium bromide staining and photodocumented.
Statistical analysis
All data are reported as means±standard deviation. Statistically significant differences were determined by the Student's t-test comparing each treatment group to the respective control. For dose–response experiments, one-way analysis of variance was used with post hoc analyses using Tukey's range test. Significance was determined at P<.05. Experiments were independently performed ≥3 times with three to eight replicates for each assay.
Results
We initially determined and confirmed the authenticity of three TCJ samples with differing lot numbers based on analysis by a commercial food and beverage testing company (Krueger Food Labs, Billerica, MA, USA) using standardized tart cherry samples and fruit-specific HPLC chromatograms for reference. As shown in Figure 1, the TCJ displayed a typical anthocyanin profile for authentic TCJ with the appearance of cyanidin-3-2G-glucosylrutinoside as the first and predominant peak (12.008 min). We also noted peaks for cyanidin-3-glucoside, cyanidin-3-rutinoside, peonidin-3-rutinoside, and an acylated anthocyanin, cyanidin aglycone, or nonanthocyanin compound appearing as the last identifiable peak (19.833 min). Table 1 shows representative results of analysis and characterization of three different batches, or lot numbers, of TCJ. All parameters were consistent with 100% pure sour, or TCJ. Using the Folin-Ciocalteu method, we determined total polyphenols in single-strength juice to be 1827±113 mg gallic acid equivalents/L. Using the pH differential method, we determined monomeric anthocyanins to be 13.8±2.2 mg cyanidin-3-glucoside equivalents (CGE)/L. Both total polyphenols and total monomeric anthocyanins were analyzed on samples before centrifugation and filtration.
We next added increasing concentrations of TCJ to MCF-7 cells for 20 h to determine the effect on CP as indicated by conversion of a tetrazolium dye to its precipitable formazan product. At concentrations of 10% and 30%, MTT reduction was decreased significantly by 35% and 90%, respectively, suggesting decreased mitogenesis and subsequent CP, likely due to necrosis (Fig. 2). We corroborated these results using BrdU incorporation as an indicator of CP. We noted significant reductions of ∼20% for BrdU incorporation at concentrations of 10% and 30% TCJ, although there was no difference in the magnitude between the two (Fig. 3).
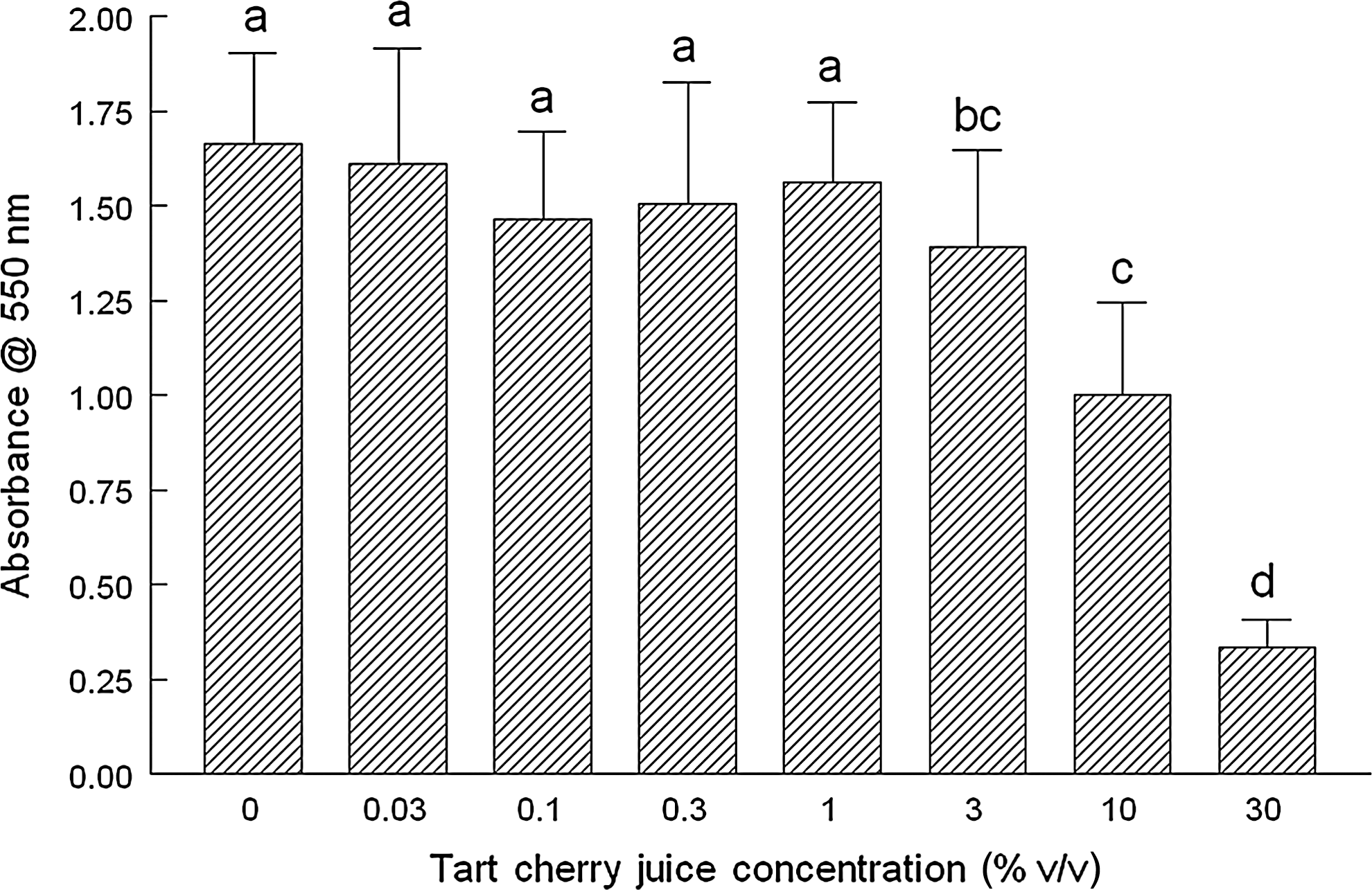
TCJ and its effects on cellular proliferation (CP) as determined by the MTT [(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay. The effect of increasing concentrations of TCJ (0.03–30% v/v) on the capacity of mitochondria to convert a tetrazolium salt to its formazan product as an indicator of CP was determined. Briefly, MCF-7 human breast cancer cells were incubated for 20 h with the indicated concentrations of TCJ. After addition of MTT, cultures were further incubated for 4 h at 37°C with subsequent centrifugation of the plate for 10 min (1500 g at 25°C) to pellet-reduced MTT. Medium was aspirated; isopropyl alcohol was added; samples were mixed (15 min at 25°C) to solubilize MTT, and solutions were analyzed at 560 nm. Data are expressed as the mean absorbance of three independent experiments±standard deviation. abcdDifferent letters indicate significant differences (P<.05) between groups.
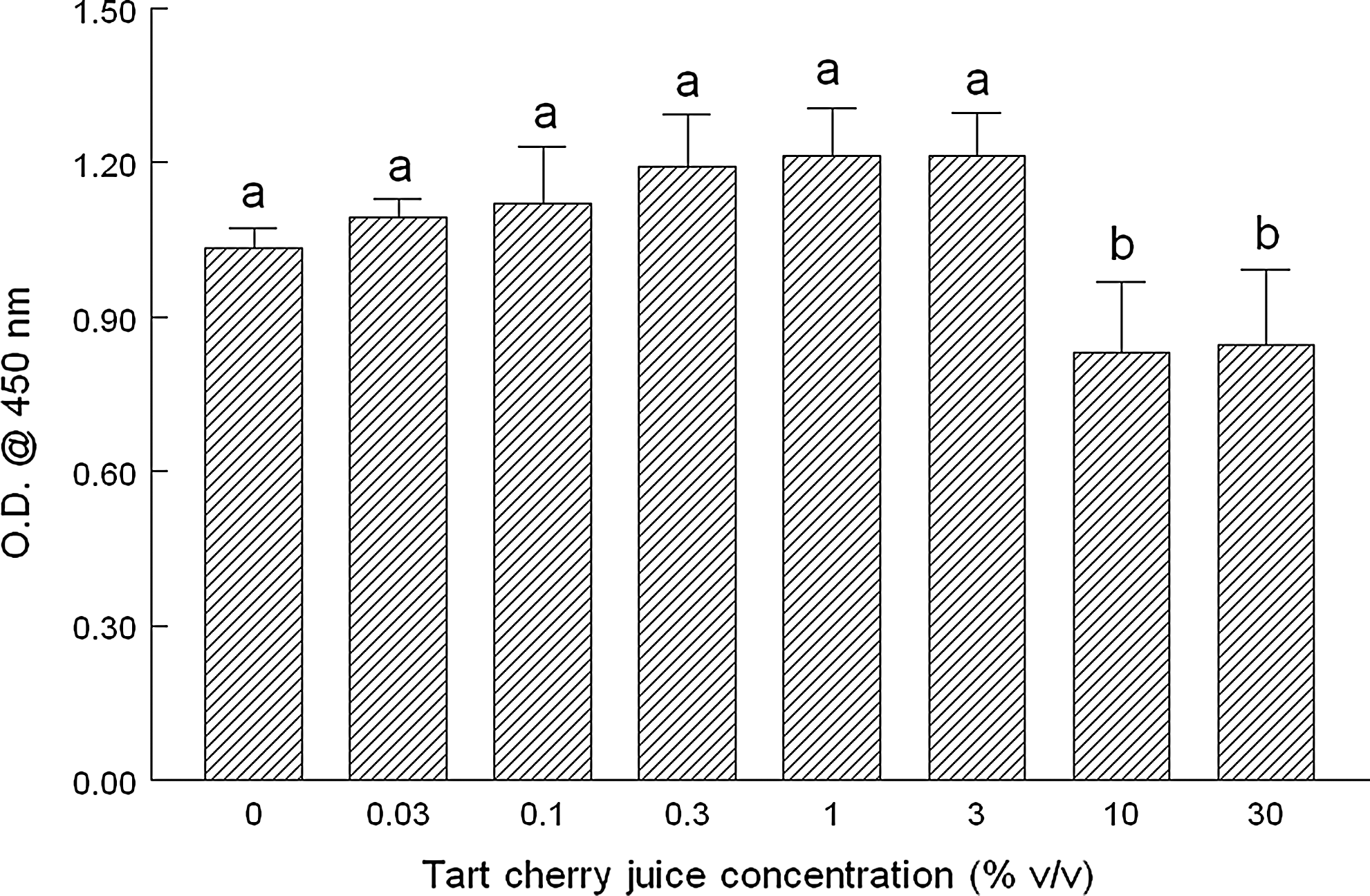
TCJ and its effects on bromodeoxyuridine (BrdU) incorporation as an indicator of mitogenesis and CP. The effect of increasing concentrations of TCJ (0.03–30% v/v) on the incorporation of BrdU into replicating DNA before cell division, viz., mitogenesis, was determined as an indirect indicator of CP. Briefly, MCF-7 human breast cancer cells were incubated for 20 h with the indicated concentrations of TCJ. BrdU was added during the last two hours of incubation. After washing monolayers and fixing cells, an enzyme-linked immunosorbent assay (ELISA) was performed as described by the manufacturer (Trevigen, Gaithersburg, MD, USA) to measure DNA incorporation of BrdU. After addition of the stop solution (2 N HCl), samples were analyzed at 450 nm by spectroscopy. Data are expressed as the mean absorbance of three independent experiments±standard deviation. abDifferent letters indicate significant differences (P<.05) between groups.
We next tested whether TCJ could induce apoptosis in MCF-7 cells using the TUNEL assay for internucleosomal DNA cleavage, which indicates nuclease activation associated with apoptosis. At concentrations of 3% and 10%, apoptosis increased by ∼34 and 44%, respectively, with vehicle-induced necrosis contributing at concentrations >10% (Fig. 4). Interestingly, we noted a significant reduction in apoptosis by 20% with the 1% TCJ treatment (Fig. 4). We corroborated these results by testing for DNA laddering or banding, using agarose gel electrophoresis, and noted nonspecific cleavage of DNA at 30% TCJ as shown by DNA smearing instead of laddering. We observed intermediate levels of faint banding between 3 and 10% TCJ and no overall discernable differences between these treatment groups and the control (Fig. 5). Cells incubated with 1% TCJ displayed banding that was less dense than the control, supporting the results from the TUNEL assay.
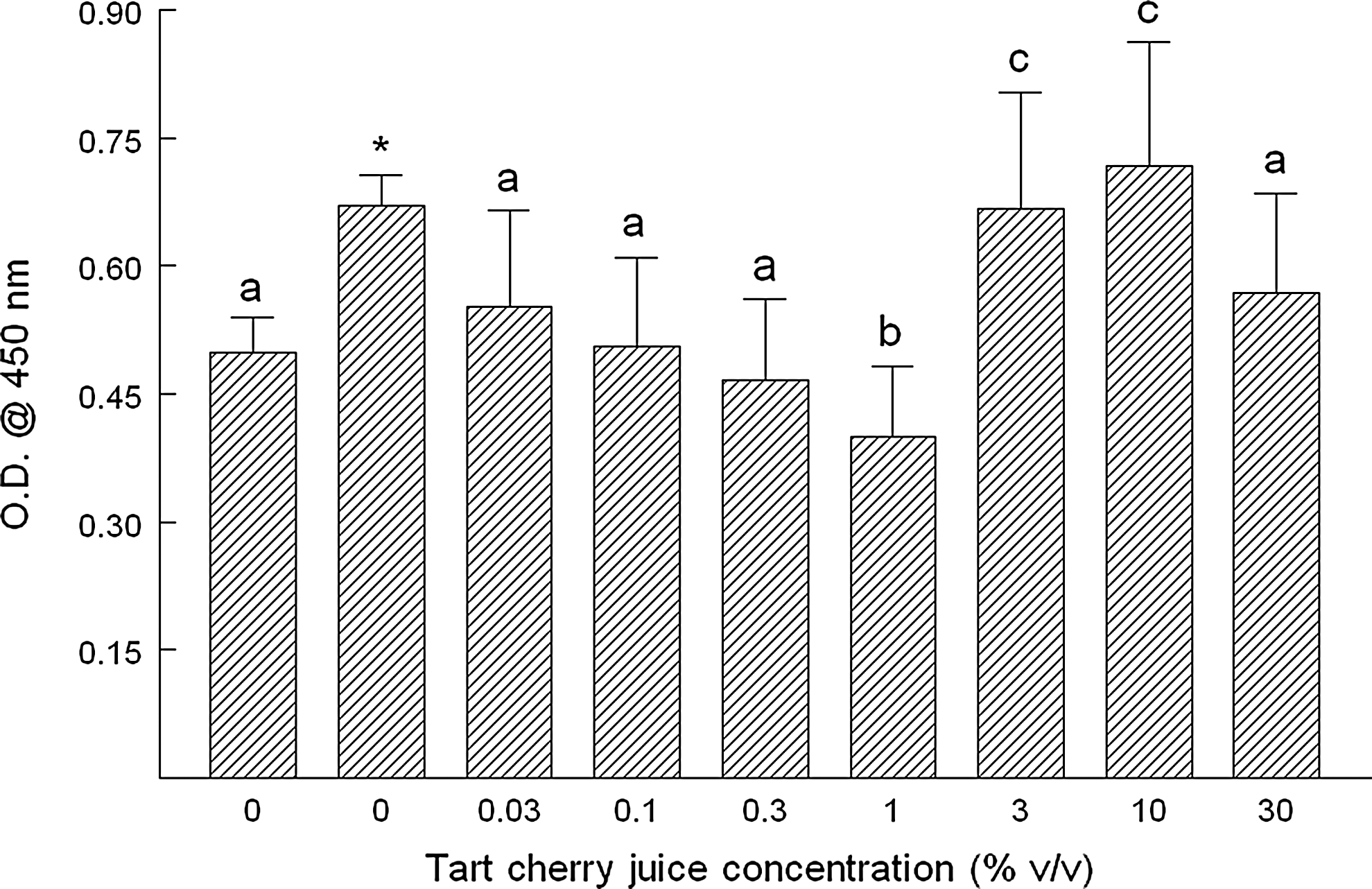
TCJ and its effects on internucleosomal cleavage of DNA indicative of apoptosis. MCF-7 human breast cancer cells were incubated for 20 h with the indicated concentrations of TCH. After washing monolayers and fixing cells, the terminal deoxynucleotidyl end-labeling (TUNEL) method was performed as described by the manufacturer (Trevigen). Briefly, after treatments, cells were pelleted by low-speed centrifugation, fixed in 37% buffered formaldehyde, and permeabilized. Endogenous peroxidase activity was quenched using 3% hydrogen peroxide in methanol, and cells were incubated for 1 h at 37°C with a labeling reaction mix (terminal deoxynucleotidyl transferase, Mn2+, and biotinylated dNTPs), followed by streptavidin horseradish peroxidase. Apoptosis was determined by color development using TACS-Sapphire substrate (Trevigen) and measured at 450 nm after addition of the stop reagent (2 N HCl). Data are expressed as the mean absorbance of three independent experiments±standard deviation. The nuclease-generated positive control is the second bar labeled as zero. Data are expressed as the mean absorbance of three independent experiments±standard deviation. *The asterisk indicates significant difference (P<.05) between the negative and positive controls. abcDifferent letters indicate significant differences (P<.05) between groups, including negative control.

Effect of TCJ on DNA fragmentation characteristic of apoptosis. Apoptosis was analyzed by DNA fragmentation, or DNA banding, using agarose gel electrophoresis. MCF-7 human breast cancer cells were incubated for 20 h with the 1% TCJ. At harvest, monolayers were washed twice with phosphate-buffered saline (PBS, then mechanically disrupted, and scraped into sterile tubes, followed by sonication to disrupt cells. Subsequently, DNA was extracted from control and treated cells using a DNA miniprep kit according to the manufacturer's instructions (Qiagen, Valencia, CA, USA). DNA (10 μg/well) was loaded onto a 1.2% agarose gel and electrophoresed for 1 h at 85 V using TBE running buffer (89 mM boric acid, 89 mM Tris base, and 2 mM EDTA). DNA was visualized by ethidium bromide intercalation into DNA and photodocumented. The lanes are identified as (1) 0% TCJ (control), (2) 1%, (3) 3%, (4) 10%, and (5) 30% TCJ.
We next determined if the aforementioned effects on MTT reduction and BrdU incorporation were correlated with a reduction in the total cell number and/or viability. We selected the 1% concentration of TCJ to further test the given differing effects between 1 and 3%. Cell numbers per T25 flask were 8.03±0.78×106 and 8.04±1.55×106 for control and TCJ flasks, respectively, and were not significantly different (Fig. 6). Viability as determined by trypan blue exclusion was 96.8±2.2% and 97.4±0.3% for control and TCJ-treated cultures, respectively, and not significantly different from one another (Fig. 7).

Effect of 1% TCJ on total cell number per flask as determined by hemocytometry. After overnight incubation with minimum essential medium (MEM) alone or containing 1% TCJ, monolayers were washed twice with PBS, trypsinized, transferred to culture tubes, centrifuged, and resuspended in fresh medium. An aliquot (50 μL) of cell suspension was added to an equal volume of trypan blue, and cells were counted by light microscopy using a hemocytometer. Data are expressed as the mean cell number per flask of three independent experiments±standard deviation.
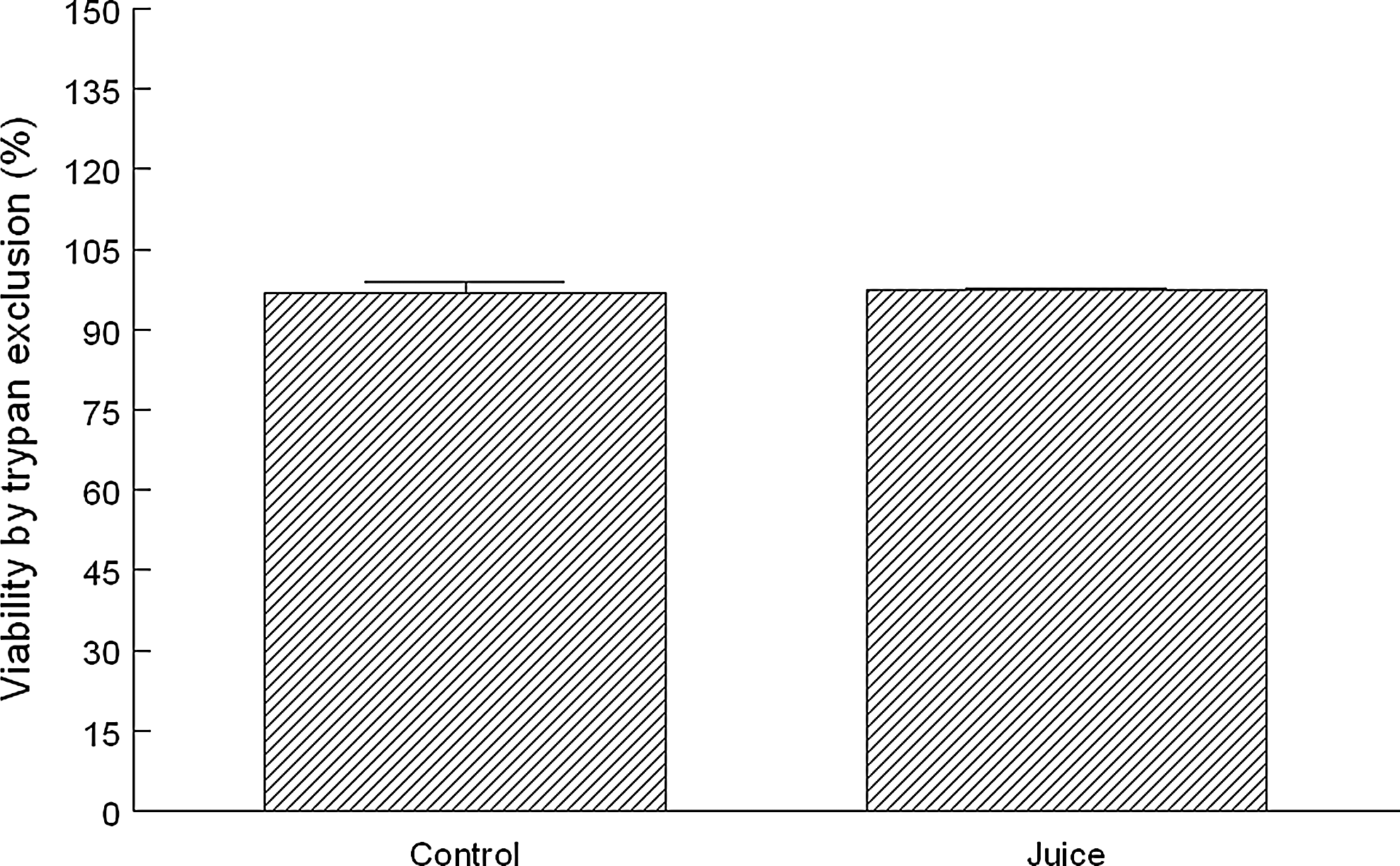
Effect of 1% TCJ on viability as determined by trypan blue exclusion. After overnight incubation with MEM alone or containing 1% TCJ, monolayers were washed twice with PBS, trypsinized, transferred to culture tubes, centrifuged, and resuspended in fresh medium as described before. An aliquot (50 μL) of cell suspension was added to an equal volume of trypan blue. Dead cells, but not live cells, stained intensely with trypan blue and indicated cytotoxicity. Viability was determined using the following formula: % viability=[(live cells)/(dead cells+live cells)]×100. Data are expressed as mean percentage of negative control and are from three independent experiments conducted in duplicate±standard deviation.
Discussion
The objective of this in vitro study was to test the capacity of increasing concentrations of 100% TCJ to influence the balance between CP and cell death, or apoptosis, and determine an optimal concentration using an in vitro model of cancer, viz., estrogen-responsive MCF-7 human breast cancer cells. We demonstrate, as expected, increased apoptosis and decreased CP at higher concentrations consistent with necrosis and due, in part, to the water vehicle. In pilot studies, we noted no adverse effects of the water vehicle at ≤5% on the parameters reported here, allowing concentrations ≥10% to serve as a positive control for markers of necrosis. We noted increased apoptosis without necrosis at 3% and decreased cell death at the 1% TCJ concentration, but not at lower concentrations and without apparent effects on CP. This suggests a transition in effects on cells between 1 and 3% consistent with the maximum concentration range of monomeric anthocyanins reported for human plasma (1.4–592.0 nM). 19
Initially, we determined the effect of TCJ on CP, since purified anthocyanins and anthocyanin-rich extracts from fruits and vegetables have been shown to exhibit antiproliferative activity toward multiple cancer cell types in vitro. 20,21 This may occur by induction of cell cycle arrest at the G2/M interface and ultimately induction of apoptosis. 22,23 At concentrations >10%, we observed significant reductions in BrdU incorporation (25%) and MTT reduction (35–90%), demonstrating decreased DNA synthesis and CP, respectively, in cells that were likely undergoing necrosis as well as apoptosis. At ≥10% TCJ, the water vehicle also significantly increased these two markers, making it impossible to discern the contribution of the TCJ to these effects. For lower TCJ concentrations (<10%), CP was not different from control, and the water vehicle did not adversely affect this marker. It is noteworthy that MCF-7 cells have a doubling time of 29 h, and it is arguable that changes in CP may not be detected if present. However, these cells are asynchronous, and at any one point in time, a subpopulation would be expected to be in the S phase and in the process of doubling, thus pulse-labeling with BrdU and subsequent DNA incorporation would permit antibody-specific detection by ELISA.
Anthocyanin-rich fruits have exhibited paradoxical effects on CP. Olsson et al. determined the effects of 10 different extracts of whole fruits and berries on CP of colon cancer and MCF-7 cells. Cherry extracts at 0.5% significantly inhibited CP of MCF-7 cells by 24–68% in a dose-dependent manner after 24 h. Purified cyanidin-3-O-glycoside (100 μg/mL) significantly inhibited the growth of MCF-7 cells by ∼30% as determined by the MTT assay with other cancer cell lines showing differing, lesser sensitivities. 24 Cooke et al. reported that purified anthocyans (such as cyanidin and cyanidin-3-glucoside), as well as many anthocyanin-rich extracts, inhibited growth of the myriad cancer cell lines by 47–85%. 5 Others have however shown proliferative activity in estrogen receptor-positive cell lines. 12 In particular, Schmitt and Stopper reported that the anthocyanidins pelargonidin chloride, cyanidin chloride, and delphinidin chloride induced CP in estrogen receptor-positive MCF-7 and BG19 human ovarian cancer cell lines, but not in the receptor-negative human breast cancer cell line MDA-MB-231. 25 Olsson et al. demonstrate using an MTT-type assay that the anthocyanin fractions of cherries and apples increased CP marginally, but significantly, to 105.9±6.8% compared to control cultures. 26 Incubation with a purified mixture of delphinidin, cyanidin, and malvidin (20 μg/mL) also significantly increased proliferation to 108.7%. Zhang et al. showed that anthocyanins (cyanidin 3-glucoside, cyanidin 3-galactoside, delphinidin 3-galactoside, and pelargonidin 3-galactoside) did not inhibit CP of numerous cell lines, including MCF-7 at 200 μg/mL, but anthocyanidins (cyanidin, delphinidin, pelargonidin, petunidin, and malvidin) inhibited proliferation using the MTT assay. 27 When considering our concentration range of 0.3–3%, we noted marginal, but not significant, increases in BrdU incorporation (Fig. 3), which is an indicator of DNA synthesis, but not specifically CP, as our hemacytometry results indicate. Noratto et al. determined that crude polyphenol extracts of yellow peach and red-fleshed plum inhibited 50% of CP at 42 mg chlorogenic acid equivalents/L for MDA-MB-435 estrogen-independent breast cancer cells compared to 515 mg/L for the estrogen-responsive MCF-7. 28
It is possible that any of the various diverse components (both nutritive and non-nutritive) of TCJ could contribute singly or in combination to the observed effects in this study. For example, TCJ is a good source of vitamins A, C, and E, minerals calcium, sodium, and potassium, melatonin, and carbohydrate. The role of carbohydrate is particularly interesting, since this macronutrient can contribute to the overall energy charge of the cell via ATP production to fuel energy-dependent processes such as apoptosis and DNA fragmentation, which could confound some assays such as MTT. MTT-type assays are ultimately dependent on functional mitochondrial succinate dehydrogenases and are largely used to demonstrate reduced CP, viz., cell number, but the results can also be interpreted as increased cytotoxicity, mitochondrial dysfunction characteristic of the intrinsic mitochondrial pathway of apoptosis, and/or altered mitochondrial energy status. Moreover, apoptosis and necrosis can occur simultaneously in cell cultures exposed to the same stimulus such as carbohydrate. 29 Nicotera et al. report that intracellular energy levels and mitochondrial function are rapidly compromised in necrosis, but not in apoptosis in neuronal cells. Repletion of the cytosolic ATP pool with glucose could in fact redirect cellular death toward apoptosis or necrosis. 29 Furthermore, Formigli et al. have determined using a xenograft model that MCF-7 cells can only undergo apoptosis when sufficient oxygen and energy supply are available. 30 Given the many components of TCJ and the difficulty in trying to correct for all of them, we did not include matched carbohydrate controls for the increasing concentrations of TCJ, although potentially a significant contributing factor.
Tart cherries also contain high levels of melatonin, an endocrine hormone, antioxidant, and mediator of circadian rhythm, compared with the melatonin concentrations in the blood of mammals with Montmorency and Balaton cherries, containing 13.46±1.10 ng/g and 2.06±0.17 ng/g, respectively. 31 Melatonin stimulates immune function and controls, in part, tumor promotion and growth due to interaction with specific melatonin receptors. Most research has centered on melatonin's role in inhibiting apoptosis in immune cells, preventing neuronal cell death, and increasing apoptotic cell death in cancer cells. 32 In fact, Eck et al. demonstrated that sequential treatment with melatonin and all trans-retinoic acid-induced apoptosis in MCF-7 human breast cancer cells. 33 Others have shown that melatonin suppresses CP in hormone-responsive MCF-7 cells in vitro, but does not affect apoptosis. 34 Melatonin clearly participates in apoptosis (increasing or decreasing) in a cell type-specific manner, although the mechanisms have not been clarified and perhaps exerts biphasic effects within the same cell line in a dose-dependent manner.
Anthocyanin-rich berry extracts exhibit proapoptotic effects in multiple human cancer cells types in vitro. We noted in this study that 3% TCJ significantly induced apoptosis in MCF-7 cells without necrosis, but at 30%, the TCJ DNA smearing characteristic of necrosis was observed in agreement with TUNEL data, suggesting concomitant apoptosis. Others have shown that the anthocyanidins delphinidin, petunidin, and cyanidin induced apoptosis (100 μM for 6 h) in HL-60 human promyelocytic leukemia cells as detected by morphology and DNA fragmentation. 35 Others have demonstrated structure-specific influences where, for example, pelargonidin, peonidin, and malvidin did not induce apoptosis. 36 In other studies, ellagic acid, a polyphenolic dimer of gallic acid, induced apoptosis (100 μM for 24 h) and reduced viability in the HUVEC cell line, normal human lung fibroblast cells HEL 299, Caco-2 colonic adenocarcinoma, MCF-7, and Hs 578T human breast cells, and DU-145 human prostatic cancer cells. 37 However, an ethanolic, anthocyanin-rich extract from black raspberry showed dose-dependent inhibition of cell growth without reducing cell viability. 20 It is likely that many other polyphenols, such as the flavonoids kaempferol and quercetin, can have a dose-dependent effect on cellular processes such as CP and apoptosis. 14,38,39
Although typically external chemical stimuli are used to induce apoptosis, commercial cell lines will undergo apoptosis in response to cultivation and the relatively stressful cell culture environment. 40,41 We noted significantly reduced apoptosis from basal levels with 1% TCJ using the TUNEL method, which was corroborated by electrophoresis, suggesting protection of MCF-7 against DNA strand breaks. Anthocyanidins such as delphinidin, cyanidin, and their glycosides inhibit tert-butylhydroperoxide-induced cytotoxicity and formation of single-strand beaks in human hepatoma cells. 12,22 A C3G-rich standardized extract from red oranges protected against DNA cleavage through dose-dependent free-radical scavenging. 42 Thus, it is not overly surprising that there could be both induction and suppression of apoptosis in the same model, given the broad range of concentrations (over three orders of magnitude) used in this study in agreement with others. 29
Both total polyphenols and total monomeric anthocyanins were analyzed on samples before centrifugation and filtration. This was based, in part, on a report by Vukosavljevic et al. showing that processing of fruit juice and subsequent microfiltration (0.2-μm pore size identical to cell culture) showed the best results for preservation of total anthocyanins, suggesting a marginal loss, if any, postfiltration. 43 Using the pH differential method, we determined monomeric anthocyanins in single-strength juice (before centrifugation and filtration steps) to be 13.8±2.2 mg CGE/L. McCune et al. report the CGE anthocyanin concentrations of harvested cherry juice to be 500–2300 mg/L, which is considerably higher than our values. 3 It is unclear why there would be such a disparity in values, although it is pointed out that many factors such as harvest year, time, temperature, and ultraviolet irradiation can markedly affect anthocyanin concentrations in cherries and presumably TCJ. 3 It is noteworthy that we used commercially processed single-strength 100% TCJ versus freshly harvested and pressed juice. This is relevant because Liu et al. evaluated different resins for commercial preparation of mulberry juice and its anthocyanins, and found that before purification with an X-5 resin, total anthocyanins were 384.1 mg/L, but were reduced considerably postfiltration to 11.5 mg/L, which is within the range of our findings. Another consideration is that although the pH differential method for total monomeric anthocyanins is a well-accepted, routinely used method validated by the Association of Analytical Communities (AOAC), it is possible that this method may be underestimating the anthocyanin concentration as suggested by others (personal communication). This argues for inclusion of a more sophisticated, corroborative analysis, and while we did analyze TCJ anthocyanins by HPLC, our data were qualitative, since the objective was simply to validate the anthocyanin pigment profile and confirm authenticity.
Identification of specific bioactive components is considerably complicated by the lack of information regarding biotransformation and metabolism by cells and inherent instability and degradation of anthocyanins under standard cell culture conditions, which may produce bioactive degradation products. 44 For example, Kern et al. report substantial degradation of anthocyanins in vitro as early as 30 min, forming phenolic acids derived from the B-ring of the anthocyanin skeleton. 45,46 In in-vitro experiments with Caco-2 cells, Kay et al. showed that after 4-h incubation in a cell-free culture medium, 57% of the initial cyanidin 3-glucoside and 96% of cyanidin had spontaneously degraded. Sadilova et al. have also demonstrated thermal degradation of acylated and nonacylated anthocyanins. 47 Kay et al. also report that in the presence of cells cultured with DMEM, there was a 31.2% greater loss of cyanidin 3-glucoside compared to buffer, primarily with production of protocatechuic acid and phloroglucinaldehyde and their respective glucuronide and sulfate conjugates. 44 Seeram et al. have identified primarily production of protocatechuic acid as well as 2,4-dihydroxybenzoic acid and 2,4,6-trihydroxybenzoic acid in McCoy's 5A medium. 48
A clear limitation of cell culture studies is that anthocyanin concentrations needed to elicit effects in vitro typically far exceed the amounts observed in human plasma in vivo, and the overall profile of bioactive molecules may be significantly different. Overall, <1% of administered doses of berry anthocyanins are absorbed in humans, presumably metabolites. 8 Fleschhut et al. have reported in two in vitro models that glycosylated and acylated anthocyanins are rapidly metabolized, or degraded, by intestinal microflora and glucuronidated by rat liver microsomes to several products. 46 In a review by Kay, the maximum plasma concentration in humans is reported to be 1.4–592 nM with elimination half-lives of 1.5–3 h (doses 68–1300 mg) and an average urinary excretion of 0.03–4% of ingested dose. 19 In a summary of 97 studies, Manach et al. report plasma concentrations of total metabolites ranged 0–400 nM with an intake of 50 mg aglycone equivalents, and the relative urinary excretion ranged 0.3–43% of the ingested dose. 49 Although the plasma levels are relatively low, it has been reported that aglycones show greater biopotency than their glycosylated forms, so the nanomolar range may be optimal. With in vitro studies, aglycones reduce cell viability generally in the range of 10–100 μM; however, concentrations >100 μM are needed for anthocyanins to elicit effects. 5 Expressing our anthocyanin results on a molar basis for CGE (molecular weight 449.2 g/mol), the concentration at 3, 10, and 30% produced final concentrations of 0.92, 3.1, and 9.2 μM, respectively, which are at the low end of the range for bioactivity for aglycones. Interestingly, the 1% concentration where protection against apoptosis was noted was 307.2 nM CGE, which falls close to the generally considered maximum concentration in human plasma.
Plasma concentrations of anthocyanins may peak in the nanomolar range and be cleared within 1 h. However, cellular concentrations could conceivably reach higher concentrations and potentially be cell type specific, that is, cancer cells. Numerous studies support differential effects on CP and viability of cancer cells with minimal adverse effects on their normal, or less-tumorigenic, cell line counterparts. 37,50,51 Moreover, it has been demonstrated that the uptake of three anthocyanins (cyanidin-3-glucoside, cyanidin-3-rutinoside, and cyanidin-3-xylosylrutinsoide) into a highly tumorigenic rat esophageal cell line was over 100 times that of the weakly tumorigenic precursor line. Furthermore, cyanidin-3-O-glucoside and cyanidin-3-O-rutinoside remained at intracellular steady-state levels for up to 12 h, but were not detectable in precursor cells after 2 h. 51
In this in vitro study, we show a biphasic effect of TCJ on induction of apoptosis, without necrosis, with protection at 1% and induced apoptosis at 3%. The 1% concentration of TCJ corresponded to 307.2 nM CGE, which is within the frequently reported maximal concentrations noted in human plasma. The data gleaned from this in vitro dose–response study may contribute to a better understanding of the concentration-dependent effects of absorbed anthocyanins on cellular processes associated with oncogenesis. Nonetheless, a clear message, particularly for those with inadequate dietary intake of anthocyanins, is that one should be consuming colorful fruits and vegetables as a part of a healthy diet. Tart cherries are particularly high in polyphenol anthocyanins and could prove to be an invaluable means of reducing the risk of chronic diseases such as cancer.
Authors' Contribution
A.W. carried out cell culture and molecular biology work and participated in the design and coordination of the study; K.R.M. designed the study, performed cell culture assays, analyzed and interpreted the data, and drafted the manuscript. All authors read and approved the final manuscript.
Footnotes
Acknowledgments
The authors thank Ginger Hook for technical assistance with laboratory assays, and Kristin Ricklefs and Giselle Pereira Pignotti for assistance with statistical analyses.
Author Disclosure Statement
The authors declare that they have no competing interests and no disclosures.