
Abstract
Paecilomyces cicadae Miquel Samson is the anamorph of Cordyceps cicadae Shing and is used in functional foods for the prevention and treatment of various diseases. In the present study, we examined the effects of P. cicadae polysaccharide (PCP) on dendritic cell (DC) maturation. Phenotypic maturation of DCs by PCP was confirmed by the elevated expressions of CD80, CD86, major histocompatibility complex (MHC)-I, and MHC-II molecules and functional maturation by increased expression of interleukin-12, interleukin-1β, and tumor necrosis factor-α, enhanced allogenic T cell stimulation, and decreased endocytosis. PCP induced the maturation of DCs from C3H/HeN and C57BL/6 mice but not from Toll-like receptor (tlr) 4–/– knockout mice and TLR4-mutated C3H/HeJ mice, which suggests that TLR4 is the membrane receptor for PCP. PCP increased the degradation of inhibitor of nuclear factor-κB (NF-κB) α/β, which enhanced the nuclear translocation of NF-κB p50/p65 and induced the phosphorylation of extracellular signal-regulated kinase, c-Jun N-terminal kinase, and p38 mitogen-activated protein kinases, which are signaling molecules downstream of TLR4. These results indicate that PCP induces DC maturation through TLR4 signaling.
Introduction
C
Cordyceps cicadae Shing is a parasitic fungus on the larvae of Cicada flammata Dist and has diverse therapeutic applications, including treatment of malaria, relief of heart palpitations, enhancement of blood aggregation, antitumor activity, and general improvement of health. 5 –7 Aqueous methanol (50%) extract from C. cicadae ascocarps enhances mononuclear cell proliferation, whereas methanol (100%) extract from C. cicadae insect body suppresses it. 5 Ergosterol peroxide isolated from C. cicadae inhibits human T cell proliferation. 8 Paecilomyces cicadae Miquel Samson is the anamorph of C. cicadae Shing, which is within the Clavicipitaceae family. 9 P. cicadae has several biological activities, including increases in interferon (IFN)-γ production by Peyer's patch cells, phagocytosis by macrophages, and spleen cell proliferation. 2,10 However, the effects of P. cicadae on dendritic cells (DCs) are unknown. In this study, we examined the immunopharmacological activities of P. cicadae polysaccharide (PCP), especially focusing on DC matuation.
DCs are professional antigen-presenting cells that are crucial for the initiation of tumor-specific T cell responses. 11 DCs are scattered throughout the body as immature cells with potent antigen uptake functions. Upon receiving microenvironmental signals, immature DCs can mature, migrate to the T cell areas of secondary lymphoid organs, and sensitize CD8+ T cells through major histocompatibility complex (MHC)-I and co-stimulatory molecules. 12 This scenario suggests that the maturation step is essential for DCs to initiate antitumor T-cell responses. However, tumor microenvironments contain small numbers of DCs, and those cells usually have the phenotype of immature DCs. 11 These immature DCs cannot induce antitumor immune responses but can induce tumor immune tolerance or anergy. 13 Defects of DCs in tumor microenvironments are an important immunological problem that limits the success of cancer immunotherapy. Here, we examine whether PCP induces DC maturation, and the underlying mechanisms are examined with respect to the characterization of the membrane receptor of PCP in DCs.
Materials and Methods
Materials
Female C57BL/6, BALB/c, C3H/HeN, and C3H/HeJ mice (6–8 weeks old) were obtained from the Korea Research Institute of Bioscience and Biotechnology (Chungbuk, Korea). Mice were housed in specific pathogen-free conditions at 21–24°C and 40–60% relative humidity under a 12-hour light/dark cycle. All animals were acclimatized for at least 1 week prior to the experiments. All experimental procedures were approved by the Animal Experimentation Ethics Committee of Chungbuk National University. Antibodies against mouse CD11c, CD40, CD80, CD86, and MHC-I/II were purchased from BD Pharmingen (San Diego, CA, USA), and those against extracellular signal-regulated kinase (ERK), p38, and C-Jun N-terminal kinase (JNK) mitogen-activated protein kinases (MAPKs) were purchased from Cell Signaling Technology (Beverly, MA, USA). Lipopolysaccharide (LPS), polymyxin B (PMB), and propidium iodide (PI) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Isolation of PCP from P. cicadae
P. cicadae 3L was kindly offered by Dr. Aiying Liu from the Institute of Fungus Resource, Guizhou University (Gzuifr, China). All strains were maintained on YMA (Difco Laboratories, Detroit, MI, USA) slants and subcultured every month. After the slants were incubated at 27°C for 7 days, they were stored at 4°C for stock. YM broth was used as a seed and fermentation medium and contained, per liter, 10 g of glucose, 3 g of malt extract, 5 g of proteose peptone, and 3 g of yeast extract. For flask cultures, all strains were grown on YMA medium in a Petri dish for 10 days. Cells were then collected and homogenized at 8400 g for 1 minute using a high-speed blender (SMT Co., Chiryu, Japan). Ten milliliters of the homogenized cell suspension was inoculated into 500-mL baffled flasks containing 200 mL of YM medium and cultivated for 11 days on a shaking incubator (Vision Biotech, Incheon, Korea) at 27°C and 140 rpm. For 5-L jar cultures, 5% (vol/vol) of the homogenized culture solution was added to a 5-L fermenter (stirred type; Kobio Tech, Seoul, Korea) containing 4,750 L of fresh YM medium. The fermenter was equipped with four equally spaced Rushton disk turbines with six flat blades. The pH was initially set at 6.5 with 1 N HCl or NH4OH but was not controlled thereafter. The dissolved oxygen tension was initially set at 100% saturation with a constant air flow rate (1.5 vessel volumes per minute) and was maintained at 20% or higher during cultivation by controlling the agitation rate (200–800 rpm). Cultivations were then performed for 120 hours. To obtain the heat-stable, nonpolymeric extract of cultured mycelia, mycelial masses were obtained by continuous centrifugation (Hanil Science Industrial, Incheon) at 10,447 g. The mycelia were washed three times with distilled water, suspended in distilled water, heated for 6 hours at 121°C, filtered, and lyophilized. Water-soluble extracts mainly contained polysaccharide and were termed PCP. No endotoxin was detected at concentrations of up to 100 μg/mL PCP as determined by the Limulus amoebocyte lysate test (catalog number 291-53101, Wako Pure Chemicals, Osaka, Japan).
Generation of bone marrow-derived DCs
DCs were generated from bone marrow (BM) cells obtained from 6–7-week-old female mice. 14 In brief, BM cells were flushed from femurs and tibias. After the red blood cells were lysed, whole BM cells (2×105 cells/mL) were cultured in 100-mm2 culture dishes in 10 mL per dish of complete medium containing 2 ng/mL granulocyte-macrophage colony-stimulating factor (GM-CSF) (R&D Systems, Minneapolis, MN, USA). On day 3, another 10 mL of fresh complete medium containing 2 ng/mL GM-CSF was added, and half of the medium was changed on day 6. On day 8, nonadherent and loosely adherent DCs were harvested by vigorous pipetting and used as immature DCs. DCs recovered from these cultures were generally >85% CD11c+ but not CD3+ and B220+.
Phenotype analysis
Phenotypic maturation of DCs was analyzed by flow cytometry. 15 Cells were stained using a combination of fluorescein isothiocyanate (FITC)-conjugated anti-CD40, anti-CD80, anti-CD86, or anti-MHC plus phycoerythrin (PE)-conjugated CD11c antibodies. Cells were analyzed using a FACSCanto™ flow cytometer (BD Biosciences, San Jose, CA, USA), and data were analyzed using WinMdi software (Scripps, La Jolla, CA, USA). Forward and side scatter parameters were used to gate live cells. Cell viability was examined by the PI nuclear staining method. Cells were stained with 1 μg/mL PI and analyzed with a FACSCanto flow cytometer. Cells stained with PI were considered dead cells.
Endocytosis assay
To analyze the endocytosis of DCs, 4×105 DCs were incubated at 37°C for 1 hour with 0.7 mg/mL FITC-dextran (42,000 Da, Sigma-Aldrich). After incubation, the cells were washed twice with cold washing buffer (phosphate-buffered saline containing 0.5% bovine serum albumin) and stained using PE-conjugated anti-CD11c antibody. Double-stained DCs were analyzed by flow cytometry. Parallel experiments were performed at 4°C to determine the nonspecific binding of FITC-dextran to DCs. 16
Cytokine assay
Total RNA was isolated using TRI™ Reagent (Molecular Research Center, Cincinnati, OH, USA). For reverse transcription–polymerase chain reaction, single-strand cDNA was synthesized from 2 μg of total RNA. 17 The primer sequences used were as follows: interleukin (IL)-12, sense, 5′-AGA GGT GGA CTG GAC TCC CGA-3′, antisense, 5′-TTT GGT GCT TCA CAC TTC AG-3′; tumor necrosis factor-α (TNF-α), sense, 5′-AGG TTC TGT CCC TTT CAC TCA CTG-3′, antisense, 5′-AGA GAA CCT GGG AGT CAA GGT A-3′; IL-1β, sense, 5′-ATG GCA ATG TTC CTG AAC TCA ACT-3′, antisense, 5′-CAG GAC AGG TAT AGA TTC TTT CCT TT-3′; IFN-α, sense, 5′-TCT GAT GCA GCA GGT GGG-3′, antisense, 5′-AGG GCT CTC CAG AYT TCT GCT CTG-3′; and β-actin, sense, 5′- TGG AAT CCT GTG GCA TCC ATG AAA C-3′, antisense 5′-TAA AAC GCA GCT CAG TAACAG TCC G-3′. 12 Polymerase chain reaction products were fractionated on 1% agarose gels and stained with 5 μg/mL ethidium bromide. Cytokine levels of IL-2 and IFN-γ in culture supernatants were measured using commercial immunoassay kits (R&D Systems).
Western blots
Lysates were prepared from total cells or nuclear as previously described. 12 Detergent-insoluble materials were removed, and equal amounts of protein were fractionated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to pure nitrocellulose membranes. Membranes were blocked with 5% bovine serum albumin in Tween 20 plus Tris-buffered saline for 1 hour and then incubated with an appropriate dilution of primary antibody in 5% bovine serum albumin (in Tris-buffered saline containing Tween 20) for 2 hours. Blots were incubated with biotinylated antibody for 1 hour and further incubated with horseradish peroxidase–conjugated streptavidin for 1 hour. Signals were detected by enhanced chemiluminescence (Amersham Pharmacia Biotech, Piscataway, NJ, USA).
Mixed leukocyte reaction
Responder T cells were purified from the spleen of BALB/c mice by negative depletion using biotinylated antibodies against B220, GR-1, and CD11c (BD Pharmingen) and Dynabeads M-280 streptavidin (Invitrogen, Carlsbad, CA, USA), as previously described. 14 Purity was typically more than 90%. DCs generated from BM cells of C57BL/6 mice were treated with 40 μg/mL mitomycin C for 1 hour. One million mitomycin C-treated DCs were added to 1×105 T cells in U-bottom 96-well plates (activator:responder cell ratio=10). Allogenic T cells were pulsed with [ 3 H]thymidine (113 Ci/nmol, NEN, Boston, MA, USA) at a concentration of 1 μCi per well for the last 18 hours and then harvested on days 3 and 5 using an automated cell harvester (Innotech, Dottikon, Switzerland). The amount of [ 3 H]thymidine incorporated into the cells was measured using a Wallac Microbeta scintillation counter (Wallac, Turku, Finland).
Statistics
Data represent mean±SD values of more than three samples, and all experiments were performed more than three times. Differences were evaluated by analysis of variance (GraphPad Software, San Diego), and P<.05 was considered significant.
Results
Phenotypic maturation of DCs by PCP
DCs were generated from BM precursors by using 2 ng/mL GM-CSF. On day 8 of culture, nonadherent and loosely adherent cells (i.e., immature DCs) were harvested from cultures and further activated with PCP or LPS for 24 hours. PCP dose-dependently increased the expression of MHC-I (Fig. 1A), MHC-II (Fig. 1B), CD80 (Fig. 1C), and CD86 (Fig. 1D), known maturation markers of DCs. PCP and LPS did not affect cell viability during the incubation (data not shown). These results demonstrate that PCP induces the phenotypic maturation of DCs. To rule out the possibility of LPS contamination, PCP was treated with PMB, a specific inhibitor of LPS. PMB strongly inhibited expression of MHC-II (Fig. 2A) and CD86 (Fig. 2B) in LPS-treated DCs but not in PCP-treated DCs. This result suggests that the PCP used in this study is free of LPS contamination.
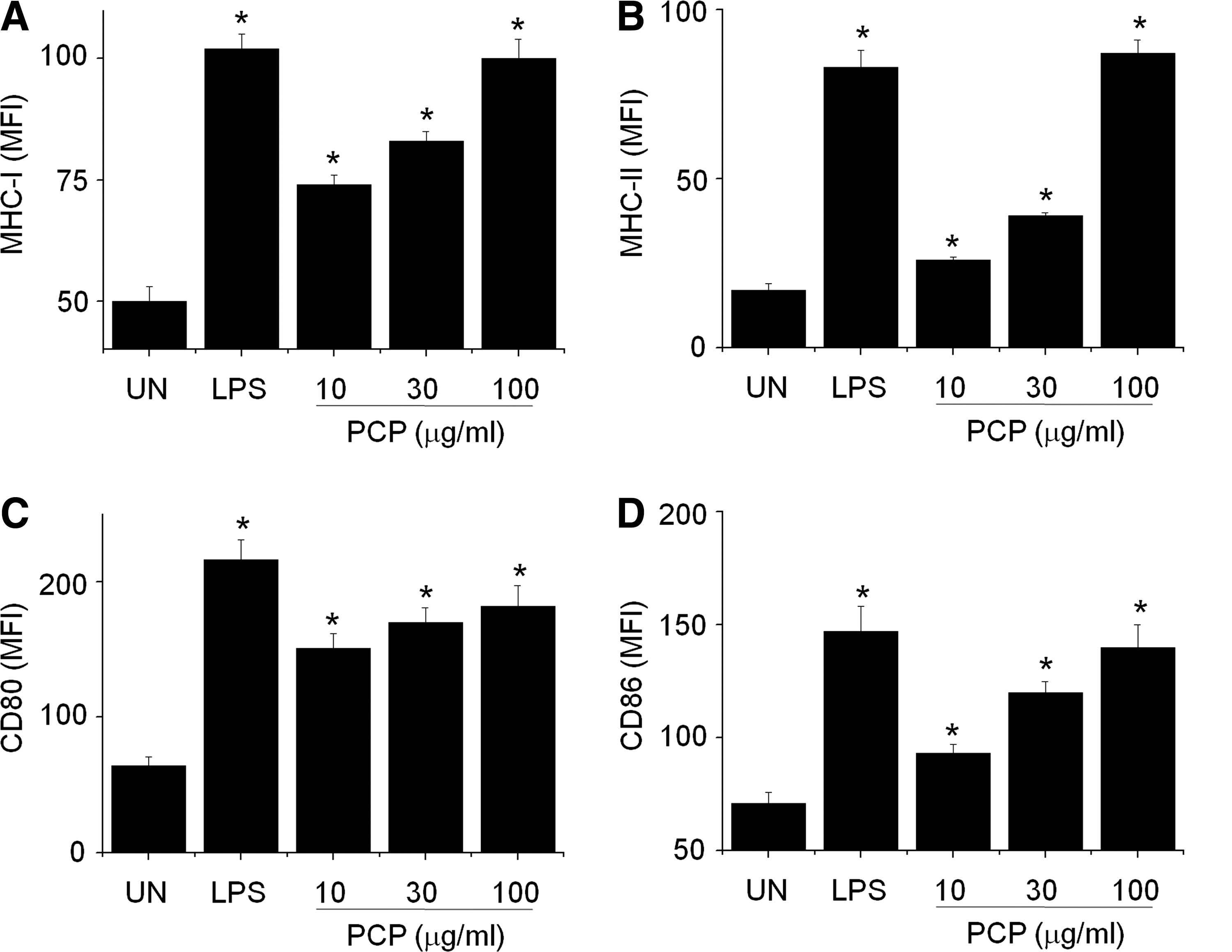
Phenotypic maturation of dendritic cells by P. cicadae polysaccharide (PCP). Dendritic cells were generated from mouse bone marrow cells by treating them with 2 ng/mL granulocyte-macrophage colony-stimulating factor for 8 days, and then cells were further activated with lipopolysaccharide (LPS) (1 μg/mL) or PCP (10–100 μg/mL) for 24 hours. Nonadherent and loosely adherent cells were harvested and stained with two antibodies (i.e., phycoerythrin-conjugated CD11c antibody plus fluorescein isothiocyanate–conjugated antibodies) to (

LPS contamination of PCP. PCP and LPS were pretreated with 1,000 U/mL polymyxin B (PMB) for 2 hours and then incubated with dendritic cells. Cells were stained with two antibodies: phycoerythrin-conjugated CD11c antibody plus fluorescein isothiocyanate–conjugated (
Functional maturation of DCs by PCP
Immature, but not mature, DCs efficiently capture antigens and show a high level of endocytosis. To determine the effect of PCP on DC endocytosis, we treated DCs with FITC-dextran and observed that antigen uptake was dose-dependently reduced in PCP-treated DCs (Fig. 3A). A similar result was obtained with LPS-treated DCs. Parallel experiments were performed at low temperature (4°C) to rule out nonspecific binding of FITC-dextran to DCs (Fig. 3B). Antigen uptake by DCs was inhibited at low temperature, which suggested that DC endocytosis was a specific active process.
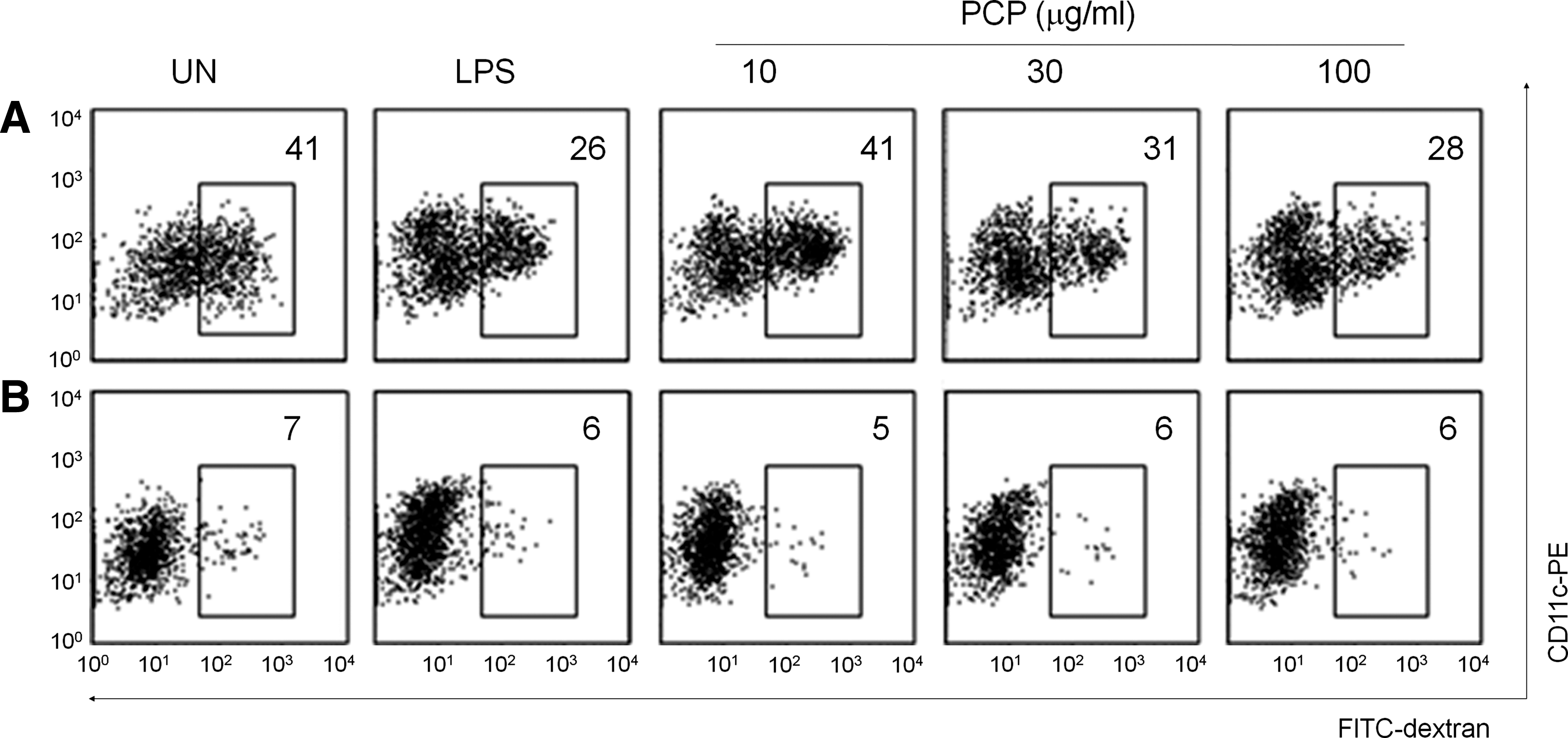
Effect of PCP on dendritic cell endocytosis. Dendritic cells were generated from mouse bone marrow cells by treating them with 2 ng/mL granulocyte-macrophage colony-stimulating factor for 8 days, and then cells were further activated with LPS (1 μg/mL) or PCP for 24 hours. (
Cytokine expression is another parameter of functional maturation of DCs. PCP strongly increased IL-12 gene expression by DCs, which is a major factor in the induction of the T-helper 1 immune response (Fig. 4A). In addition, PCP increased the gene expression of inflammatory cytokines, such as IL-1β and TNF-α, by DCs (Fig. 4A). IL-2, IL-4, and IFN-γ were not detected in DCs (data not shown). Protein production of IL-12, IL-1β, and TNF-α by PCP-treated DCs was quantitatively measured by enzyme-linked immunosorbent assay (Fig. 4B). These results demonstrate that PCP induces the functional maturation of DCs.
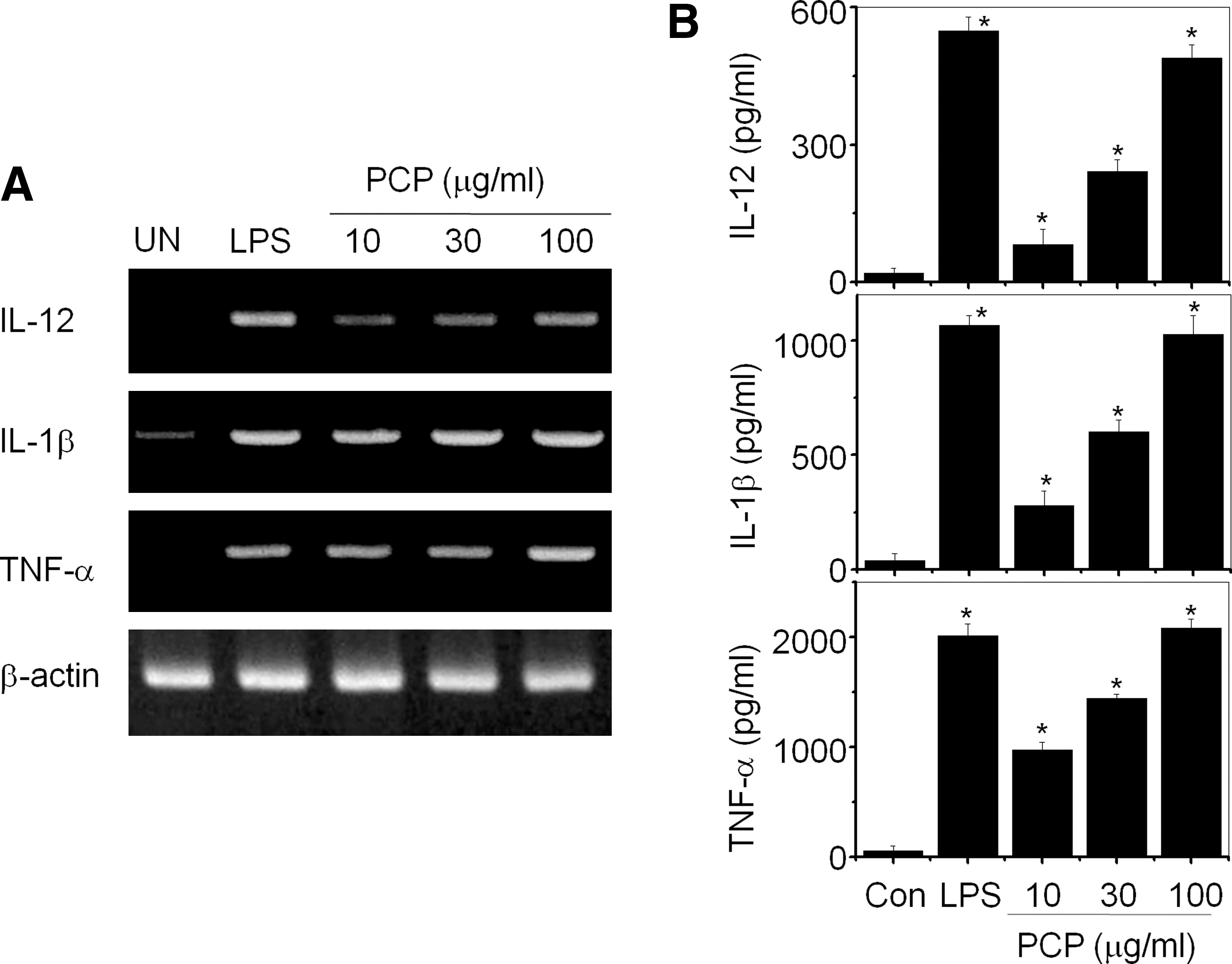
Effect of PCP on cytokine production of dendritic cells. Dendritic cells were generated from mouse bone marrow cells by treating them with 2 ng/mL granulocyte-macrophage colony-stimulating factor for 8 days, and then cells were further activated with LPS (1 μg/mL) or PCP for 4 hours. Nonadherent and loosely adherent cells were harvested. Total RNA was isolated, and (
Allogenic T-cell activation by PCP-treated DCs
The phenotypic and functional maturations of DCs by PCP suggest that PCP-treated DCs might act as stimulator cells during allogenic T cell responses. To test this hypothesis, we induced a mixed leukocyte response using C57BL/6 mouse-derived DCs (H-2b) and BALB/c mouse-derived T cells (H-2d). At 3 (Fig. 5A) or 5 (Fig. 5B) days after mixing T cells and DCs, we examined T-cell proliferation. PCP-treated DCs strongly enhanced allogenic T-cell proliferation, but PCP-untreated DCs did not. T cells or DCs alone could not proliferate. We also examined the production of T cell–derived cytokines in culture medium 2 days after mixing DCs and T cells. PCP-treated DCs could strongly activate T cells to produce IFN-γ (Fig. 5C) and IL-2 (Fig. 5D). However, PCP-untreated DCs showed weak stimulating activity. These results suggest that PCP-treated DCs have high capacity to stimulate allogenic T cells.
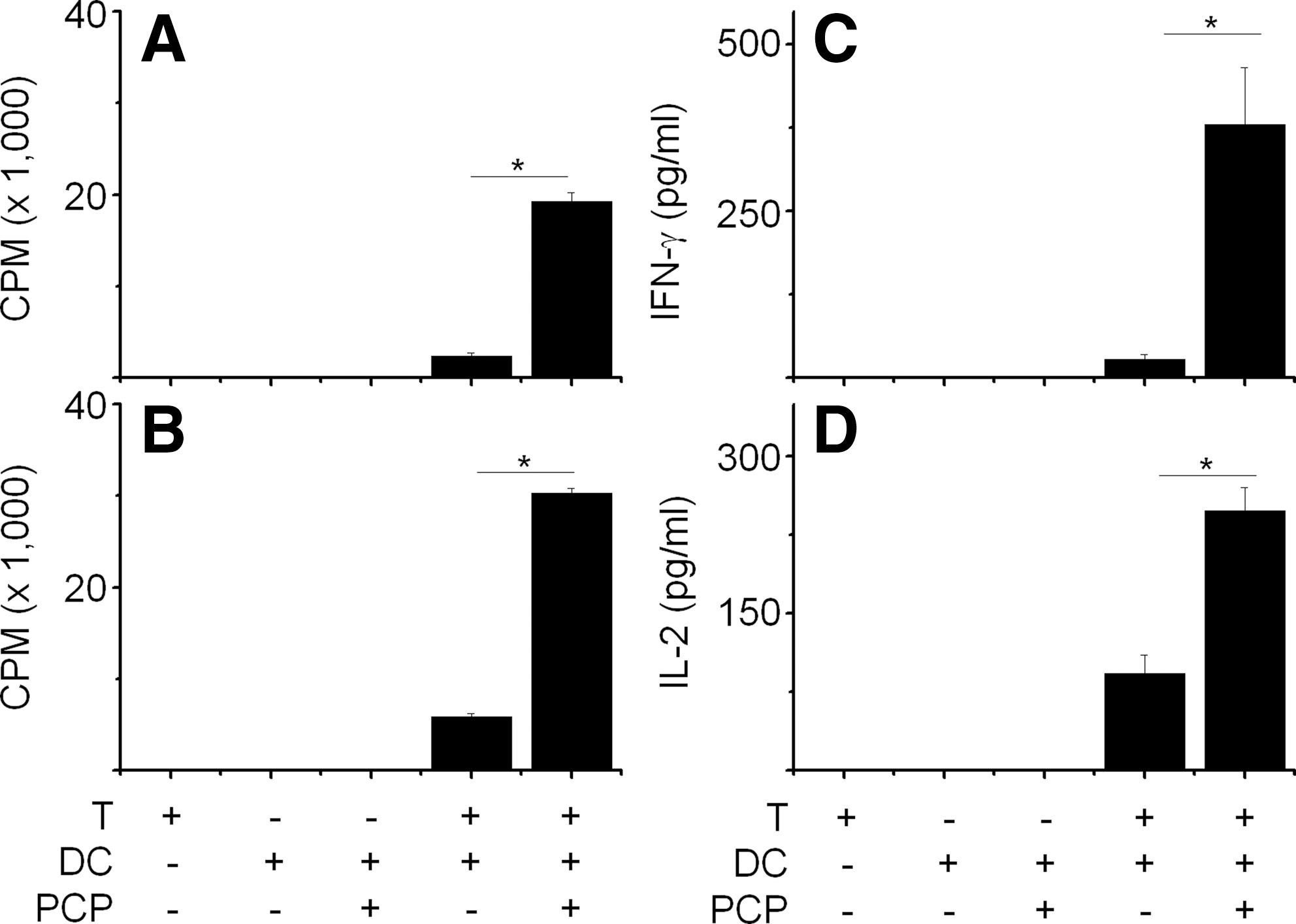
Allogenic stimulation of T cells using PCP-treated dendritic cells (DCs). DCs were generated from bone marrow cells of C57BL/6 mice by treating them with 2 ng/mL granulocyte-macrophage colony-stimulating factor for 8 days and further activated with PCP for 24 hours. DCs were harvested and treated with 40 μg/mL mitomycin C for 1 hour to prevent proliferation. Mitomycin C-treated DCs (1×104) and T cells (1×105) were mixed and incubated for (
Identification of membrane receptor of PCP
Because PCPs cannot penetrate DCs due to their size, DC maturation may be caused by surface binding of PCP to membrane receptors. We presumed that PCP activates DCs through Toll-like receptor (TLR) 4, an LPS receptor, because PCP and LPS influenced DC maturation in a similar manner. In this experiment, we generated DCs from two different mouse strains (i.e., C3H/HeN having normal TLR4 and C3H/HeJ having mutated TLR4). PCP increased the surface expressions of MHC-II (Fig. 6A), CD80 (Fig. 6B), and CD86 (Fig. 6C) in DCs generated from C3H/HeN mice but not C3H/HeJ mice. We also evaluated the effect of PCP on DC endocytosis, and, as expected, PCP decreased endocytosis of DCs from C3H/HeN mice but not TLR4-mutated C3H/HeJ mice (Fig. 6D).
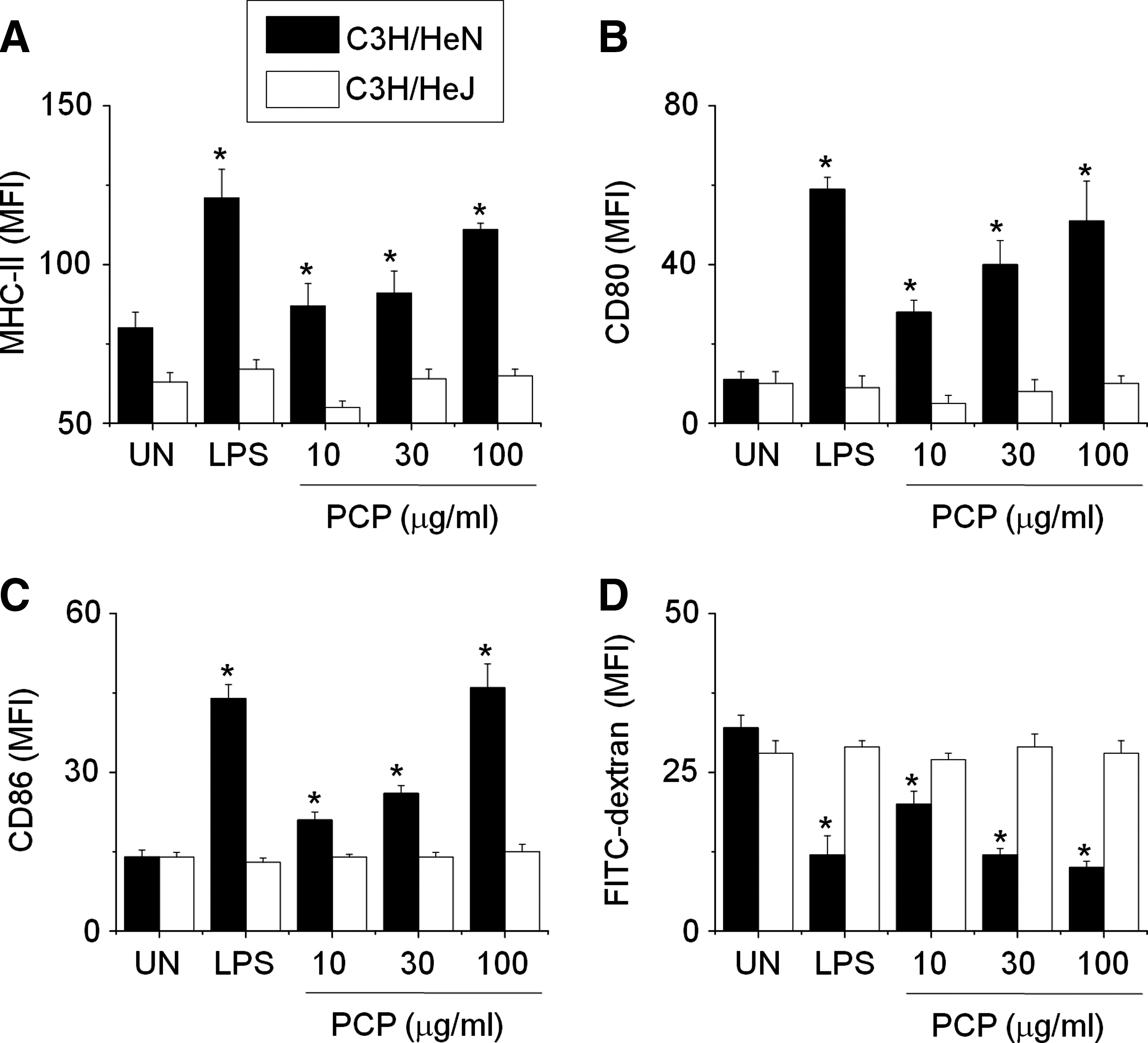
Effect of PCP on maturation of Toll-like receptor-mutated DCs. To study membrane receptors of PCP, DCs were generated from bone marrow cells of C3H/HeN mice (normal Toll-like receptor 4) and C3H/HeJ mice (mutated Toll-like receptor 4) by treating them with 2 ng/mL granulocyte-macrophage colony-stimulating factor for 8 days and then further activating cells with LPS (1 μg/mL) or PCP for 24 hours. Nonadherent and loosely adherent cells were harvested and stained with two antibodies: PE-conjugated CD11c antibody plus FITC-conjugated antibodies to (
To evaluate the direct involvement of TLR4 in DC activation, we examined the effect of PCP on DCs generated from tlr4–/– mice. PCP increased the surface expression of CD86 (Fig. 7A) and MHC-II (Fig. 7B) in DCs generated from tlr4+/+ wild-type mice, but not from tlr4–/– knockout mice. In addition, PCP also did not affect endocytosis of DCs from tlr4–/– knockout mice (Fig. 7C). PCP induced gene expression of IL-12, IL-1β, TNF-α, and IFN-α of tlr4+/+ DCs but not those of tlr4–/– DCs (Fig. 7D). The TLR3 ligand poly(I:C) could induced cytokine gene expression of tlr4–/– DCs. Overall, these data suggest that PCP increases DC maturation and function through TLR4.
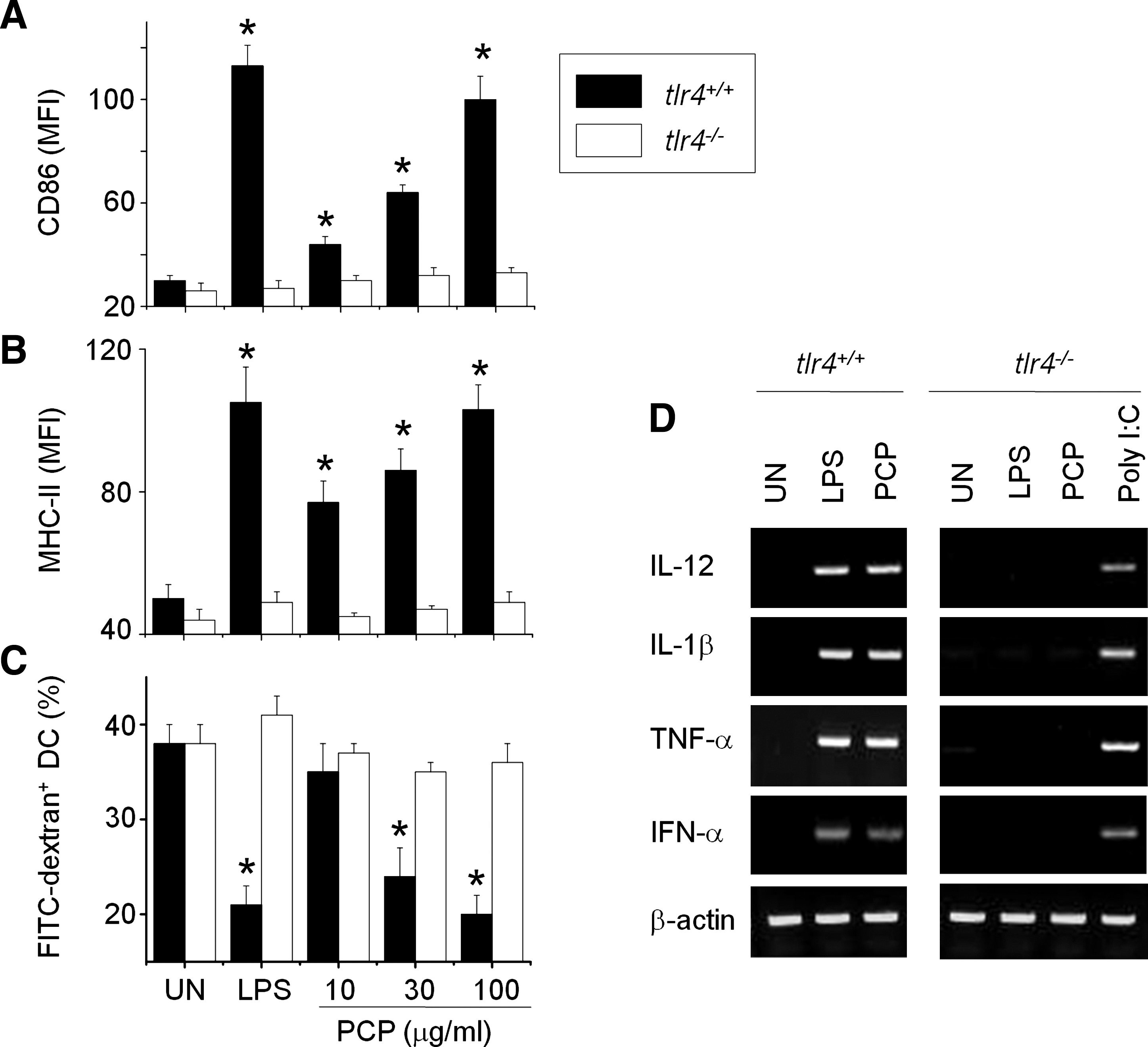
Effect of PCP on maturation of Toll-like receptor (tlr) 4–/–
DCs. DCs were generated from tbone marrow cells of tlr4+/+
or tlr4–/–
mice by treating them with 2 ng/mL granulocyte-macrophage colony-stimulating factor for 8 days and then further activating cells with LPS (0.5 μg/mL) or PCP for 24 hours. DCs were stained with CD11c-PE plus (
Effect of PCP on MAPK and nuclear factor-κB signalings
Upon binding with LPS, TLR4 activates both MyD88-dependent and TRIF-dependent signaling pathways. Although the detailed events downstream of MyD88 and TRIF differ, MAPKs and nuclear factor-κB (NF-κB) signaling are commonly activated. Furthermore, signaling pathways involving NF-κB and MAPK play major roles in DC maturation. Thus, we investigated the activation of MAPKs and NF-κB in PCP-treated DCs. The majority of NF-κB subunits are sequestered in the cytoplasm by inhibitor of NF-κB (IκB) α/β and translocated into the nucleus after IκBα/β degradation. As shown in Figure 8A, total IκBα levels in PCP-treated DCs decreased within 15 minutes and gradually increased thereafter. Total IκBβ levels in PCP-treated DCs slowly decreased within 60 minutes and subsequently increased after 120 minutes. LPS produced a similar pattern as PCP (Fig. 8B). Finally, PCP induced the nuclear translocation of NF-κB p50 and p65, as demonstrated by the increased levels of nuclear p50/p65 proteins (Fig. 8C). As shown in Figure 8D, basal levels of phosphorylated ERK, JNK, and p38 MAPKs in immature DCs were low, whereas phosphorylation of MAPKs was profoundly increased over basal levels in DCs upon exposure to either LPS or PCP, indicating that MAPK pathways might be involved in PCP-induced DC maturation. These results suggested that PCP induced DC maturation through MAPK and NF-κB signalings that were downstream of TLR4.
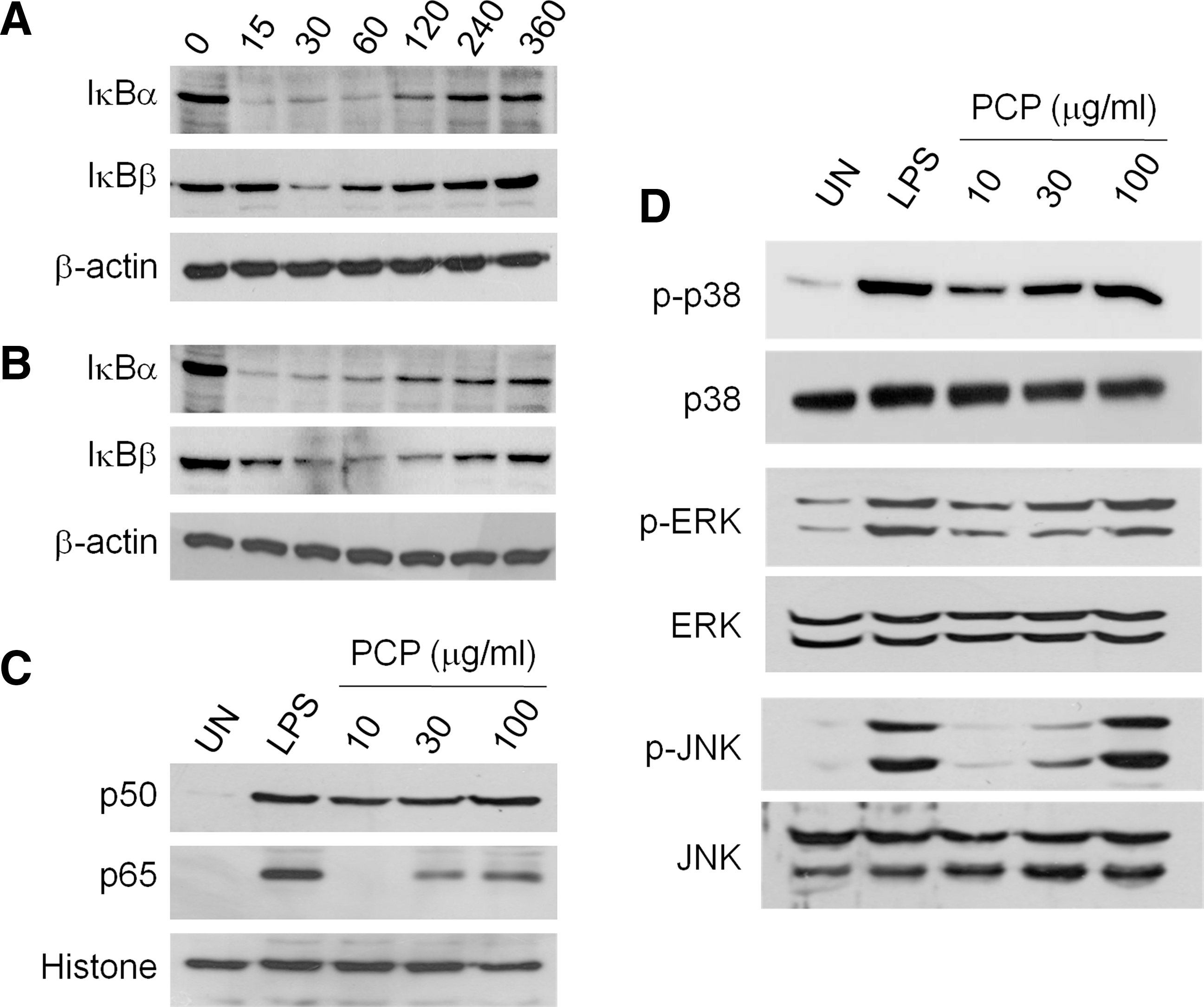
Activation of nuclear factor-κB (NF-κB) and mitogen-activated protein kinase signaling in DCs by PCP. DCs were generated from mouse bone marrow cells by treating them with 2 ng/mL granulocyte-macrophage colony-stimulating factor for 8 days, and the cells were then further activated with (
Discussion
In the present study, the immunostimulating effects of PCP on DCs were examined, and the mechanism of PCP action was partially elucidated with respect to the membrane receptor. PCP induced phenotypic maturation of DCs as demonstrated by the increased expressions of MHC-I, MHC-II, CD80, and CD86. Three functional changes were also demonstrated in PCP-treated DCs: decreased endocytosis, increased cytokine production, and an increased capacity to induce allogenic T-cell activation. Two biochemical changes in PCP-treated DCs were shown: increased phosphorylation of MAPKs (ERK, p38, and JNK) and enhanced nuclear translocation of NF-κB p50/p65. Of interest was that PCP might use TLR4 as a membrane receptor in DCs. PCP did not induce maturation and IL-12 expression in DCs generated from TLR4-mutated C3H/HeJ mice and tlr4–/– knock-out mice.
The induction of a successful antitumor immune response requires the use of immune-stimulatory mature DCs because of their enhanced capacity to induce antigen-specific T-cell responses. 18 In addition, mature DCs have been shown to be resistant to immunosuppressive factors produced by tumors and are phenotypically and functionally stable in the absence of cytokines. 19,20 It has been reported that tumors prevent DC maturation and function by releasing vascular endothelial growth factor, tumor growth factor-β, prostaglandin E2, IL-13, IL-10, and IL-6. 21 –26 Defective DC maturation in tumor microenvironments is an important immunological problem that limits the success of cancer immunotherapy. DC maturation can be achieved by several stimuli: chemokines, pro-inflammatory cytokines, and TLR ligands. 12,18 Zymosan and peptidoglycan have been used as TLR2 ligands, LPS and monophosphoryl lipid A (MPL) as TLR4 ligands, poly(I:C) as a TLR3 ligand, and CpG as a LR9 ligand. Currently, the most popular stimulus is a cocktail containing IL-1β, IL-6, TNF-α, and prostaglandin E2. 27 –30 The maturation process is associated with several coordinated events, such as loss of endocytic receptors, up-regulation of co-stimulatory molecules, changes in morphology, and changes in MHC compartment, all of which are shown in PCP-treated DCs. The different stimuli used to induce maturation give rise to subtle differences in the DC maturation stage. These differences can be observed at the phenotypical expression level of MHC-I/II and CD80/86, at the functional level of allogenic T-cell activation, and at the cytokine expression levels of IL-12, IL-1β, and TNF-α. Semimature DCs, which express high levels of MHC and co-stimulatory molecules and produce trace amounts of IL-12, have been implicated in the conversion of naive T cells into regulatory T cells. 31 From this point of view, full DC maturation might be important to induce optimal antitumor T cell response. In this study, we showed that PCP fully induced DC maturation by up-regulating phenotypic and functional parameters. PCP increased the expression level of MHC and co-stimulatory molecules and furthermore strongly increased IL-12 expression, followed by significant augmentation of allogenic T-cell activation.
We also partially characterized the PCP receptor in DCs. Because of its molecular mass, we reasoned that PCP cannot penetrate cells and that DC activation is therefore caused by PCP binding to receptors expressed on DCs. We first hypothesized that TLR4 was a possible PCP receptor because PCP and LPS, which was used as a reference compound in this study, have similar effects on DCs. The role of TLR4 as a PCP receptor was clearly demonstrated by the observation that DCs from C3H/HeJ mice, which have naturally mutated TLR4, are hyporesponsive to PCP compared with normal DCs from C3H/HeN mice. The role of TLR4 as the LPS receptor was also confirmed by the reduced responsiveness of C3H/HeJ DCs to LPS. One question raised in this study was whether PCP was free of LPS contamination. The field of PCP research has been confounded by the presence of the endotoxin LPS in sample preparations. In this study, to rule out the possibility of LPS contamination in PCP, we validated PCP purity in two ways. First, we treated PCP with PMB, a specific inhibitor of LPS. PMB strongly inhibited MHC-II expression by LPS-treated DCs but not by PCP-treated DCs. Second, we examined endotoxin levels in PCP by using the Limulus amebocyte lysate test. Although the traditional Limulus amebocyte lysate test is the standard and most sensitive assay for detecting bacterial endotoxin, it also responds to β-glucan or polysaccharide. 32 We also observed that most commercially available kits were positive to polysaccharides (data not shown). However, several kits, including the Limulus Color KY Test (Wako Pure Chemicals), were specific for LPS but not polysaccharide. No endotoxin was detected at concentrations of up to 100 μg/mL PCP, indicating that the PCP was free of LPS contamination and that DC maturation was PCP specific.
The involvement of TLR4 in DC activation is well established. It has been reported that in vitro and in vivo exposure to LPS causes DCs to undergo activation and maturation. 33 LPS-induced DC maturation involves at least two important signaling pathways: (a) ERK activation, which enables DC survival, and (b) NF-κB/Rel activation, which facilitates DC maturation. This coordinated process leads to a high T cell-stimulatory capacity and to IL-12 release, both of which result in the induction of a protective immune response. The present study provides evidence that PCP, like LPS, induces DC maturation through ERK and NF-κB activation. Many inducers of DC maturation are strong activators of NF-κB transcription factors, suggesting that these factors may play a key role in DC maturation. 34 Members of the NF-κB family of transcription factors exist as homodimers or heterodimers of five proteins (p50, p52, RelA, RelB, and cRel) and play an important role in regulating inflammatory gene expression. 35 NF-κB activation occurs by nuclear translocation following inducible phosphorylation of inhibitory IκB proteins by the IκB kinase complex. 36 Promoter studies have shown that NF-κB sites play an important role in regulating expression of MHC and co-stimulatory molecules. 35,37 These sites are also essential for IL-12 expression. 34
TLR4 signaling pathways may play a common role in immune cell activation by polysaccharide biological response modifiers. TLR4 is expressed on myeloid cells (monocytes, macrophages, and DCs) and B cells, which are the main stimulatory targets of polysaccharide biological response modifiers. 38 Polysaccharide from Phellinus linteus, Angelica gigas Nakai, and Cordyceps militaris interact with TLR4 in DCs. 12,14,39 Other polysaccharides from Ganoderma lucidum, Astragalus membraneceus, Acanthopanax koreanum, Acantopanax senticosus, Dioscore alata, Inonotus obliquus, Angelica gigas Nakai, and Platycodon grandiflorum have been reported to activate B cells and macrophages through TLR4. 12,40 –45
During neoplastic disease development, the body's natural defenses are usually attenuated. In particular, DCs that have infiltrated tumor tissues are known to exhibit reduced expressions of co-stimulatory molecules and defective cytokine production, which implies that tumor-derived factors impede DC maturation. 39 Defective DCs in tumor microenvironments are an important immunological problem that limits the success of cancer immunotherapy. In the present study, we found that PCP can induce phenotypic and functional maturation of DCs through TLR4. Thus, PCP might increase the efficiency of DC-based cancer immunotherapy.
Footnotes
Acknowledgments
This research was supported by the Medical Research Center program (grant 2010-0029480), the Happy Tech. program (grant 2010-0020806), and the Regional Core Research program through the National Research Foundation of Korea.
Author Disclosure Statement
No competing financial interests exist. All the authors have substantially participated in the investigation, data analysis, and the preparation of the manuscript and accept full responsibility for its content.