
Abstract
The pepper fruit of Capsicum annuum L. is used as a food, spice, and topical medicine. Here, we investigated the effect of a methanolic C. annuum L. extract (CAE) in a mouse model of ovalbumin-induced allergic airway inflammation. Animals were treated with CAE by oral gavage before ovalbumin challenge. After ovalbumin challenge, airway responsiveness to methacholine, influx of inflammatory cells into the lung, cytokine levels in bronchoalveolar lavage fluid and lung, nuclear factor-κB (NF-κB) activity in lungs, and lung histopathology were assessed. Oral treatment with CAE significantly reduced the pathophysiological signs of allergic airway disease, including increased inflammatory cell recruitment to the airways, airway hyperresponsiveness, and increased levels of T-helper type 2 cytokines. Reactive oxygen species were also decreased in cells from bronchoalveolar lavage fluid. In addition, we found that administration of CAE attenuated ovalbumin-induced increases in NF-κB activity in lungs. Collectively, these results suggest that CAE may be an effective oral treatment for allergic airway inflammation by virtue of its antioxidant activity.
Introduction
A
Pepper fruits of Capsicum annuum L. are widely used as food and as a spice. Many previous studies have demonstrated that peppers contain a wide array of phytochemicals, mainly vitamins C, A, and E, as well as phenolic and carotenoid compounds with well-known antioxidant properties. 3 –5 Hot cultivars are rich in capsaicinoids (alkaloids with pharmacological properties), giving the pepper fruit its characteristic taste. 6 Peppers are also a good source of provitamin A carotenoids (β-carotene, α-carotene, and β-cryptoxanthine) and oxygenated carotenoids or xanthophylls. These compounds have been shown to be effective free radical scavengers and may be effective for preventing various diseases associated with free radical oxidation, such as cardiovascular disease, cancer, and neurological disorders. 5,7,8 Several previous studies have reported on the antioxidant activity of C. annuum L. extract (CAE), but the effects of CAE in allergic airway disease have not yet been determined. In this study, we described the ability of CAE to reduce asthmatic airway oxidative stress and airway inflammation in an ovalbumin (OVA)-sensitized mouse model of allergic airway disease.
Materials and Methods
Preparation of CAE
This study was carried out on fruits of C. annuum L. collected and verified by Dr. Manuel Sandoval (Center for Research on Amazonian Natural Products, Loreto, Peru). Voucher specimens have been deposited in the International Biological Material Research Center at the Korea Research Institute of Bioscience and Biotechnology, Daejeon, Korea. Methanolic CAEs were prepared by exhaustively extracting dried and powdered pepper fruits with methanol three times at room temperature (yield, 15%). Concentrated, methanolic CAEs were freshly dissolved in phosphate-buffered saline before use.
Animals
Female BALB/c mice, 6–8 weeks of age (Daehan Biolink Co., Seoul, Korea), were used in compliance with protocols approved by the Institutional Animal Care and Use Committee of the Korea Research Institute of Bioscience and Biotechnology.
Experimental protocol
Specific-pathogen-free female BALB/c mice were sensitized by intraperitoneal injection of 20 μg of OVA (catalog number A5503, Sigma-Aldrich, St. Louis, MO, USA), emulsified in 2 mg of aluminum hydroxide (Sigma-Aldrich) in 200 μL, on days 1 and 14. The mice underwent aerosol challenges for 20 minutes with 1% OVA in saline using an ultrasonic nebulizer (model NE-U12, Omron Corp., Tokyo, Japan) on days 28, 29, and 30 after initial sensitization. 9,10 One hour before each challenge, one group of asthma-induced mice was treated with saline, a second group was treated with 3 mg/kg dexamethasone (Sigma-Aldrich), a third group was treated with 10 mg/kg CAE, and a fourth group was treated with 30 mg/kg CAE. Mice in the control group were sensitized and challenged with saline without drug treatment. Saline, dexamethasone, and CAE were administered by oral gavage. Airway hyperresponsiveness was measured 24 hours after the last challenge, and mice were sacrificed 48 hours after the last challenge for harvest of bronchoalveolar lavage (BAL) fluid, serum, and lungs.
Measurement of airway responsiveness
Airway responsiveness to methacholine (Sigma-Aldrich) was measured indirectly using single-chamber, whole-body plethysmography (Allmedicus, Seoul). 9 –12 Conscious mice were challenged with aerosolized saline and increasing concentrations of methacholine (6.25–50 mg/mL) for 3 minutes; following each challenge, enhanced pause values were calculated for 3 minutes.
Inflammatory cell counts in BAL fluid
BAL was performed as described previously. 13,14 BAL fluid was obtained from mice anesthetized with 50 mg/kg pentobarbital (Hanlim Pharm. Co. Ltd., Seoul) by lavaging lungs with saline delivered via a tracheal cannula. BAL fluid was deposited onto Cytospin slides and stained with Diff-Quik (Dade Behring Inc., Deerfield, IL, USA). Differential cell counts were performed by two independent investigators.
Measurement of intracellular reactive oxygen species
After BAL fluid cells was washed with phosphate-buffered saline, oxidative stress was assessed by incubating BAL cells for 10 minutes at 37°C with 10 μM 2′,7′-dichlorofluorescein diacetate (Sigma-Aldrich). Intracellular reactive oxygen species (ROS) activity was determined by measuring fluorescence at excitation of 488 nm and emission of 525 nm with a spectrofluorometer (Perkin-Elmer Ltd., Beaconsfield, United Kingdom).
Measurement of total immunoglobulin E and OVA-specific immunoglobulin E in serum
Serum levels of total and OVA-specific immunoglobulin E (IgE) were measured by enzyme-linked immunosorbent assay as described previously. 9 In brief, microtiter plates were coated with 100 μL per well of IgE (2 μg/mL) (Serotec, Oxford, United Kingdom) in phosphate-buffered saline containing Tween-20. Antibodies in BAL fluid were detected using isotype-specific secondary antibodies (anti-mouse IgE, Serotec). After four washes, 200 μL of o-phenylenediamine dihydrochloride (Sigma-Aldrich) was added to each well. The plate was incubated for 10 minutes in the dark, and absorbance was measured at 450 nm. Total and OVA-specific IgE concentrations were calculated from a standard curve generated from a 250 ng/mL solution of recombinant IgE (Serotec).
Measurement of T-helper type 2 cytokines in BAL fluid
Levels of T-helper type 2 (Th2) cytokines interleukin (IL)-4, IL-5, and IL-13 in BAL fluid were measured using an enzyme-linked immunosorbent assay kit (BioSource International, Camarillo, CA, USA) according to the manufacturer's instructions.
Western blot analysis
Both nuclear and cytoplasmic extracts from mouse lung were prepared with an NE-PER® nuclear and cytoplasmic extraction reagent kit (Pierce, Rockford, IL, USA) according to the manufacturer's instructions. Nuclear and cytoplasmic fractions were resolved on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride membrane (Amersham Life Sciences, Piscataway, NJ, USA). The blots were blocked with 5% fat-free dry milk in phosphate-buffered saline containing 0.1% Tween-20 for 1 hour at room temperature. Nuclear factor-κB (NF-κB) p65 protein was detected with an anti-p65 antibody (Cell Signaling Technology, Beverly, MA, USA) as previously described. 10 Horseradish peroxidase–conjugated secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used as a secondary antibody. The transferred proteins were visualized with an enhanced chemiluminescence detection kit (Amersham Pharmacia Biotech, Uppsala, Sweden). Densitometric band values were determined using UN-SCAN-IT Gel™ image analysis software (Silk Scientific, Orem, UT, USA) and statistically analyzed.
Histological analysis
Lungs were fixed with 4% paraformaldehyde prior to embedding in paraffin. Sections (4 μm thick) were stained with hematoxylin and eosin to assess inflammatory cell infiltration. Peribronchial and perivascular inflammation was evaluated using a semiquantitative scoring system with a grading scale ranging from 0 (no inflammatory cells) to 3 (very severe inflammation). 10,11 Samples were scored in a blinded fashion. Goblet cell hyperplasia was assessed by periodic acid–Schiff (PAS) staining. Mucus levels in the airway were quantified by counting PAS-positive and PAS-negative epithelial cells in each bronchiole, as described previously. 9,12,14 Results were expressed as the percentage of PAS-positive cells per bronchiole, calculated as the number of PAS-positive epithelial cells per bronchiole divided by the total number of epithelial cells in each bronchiole.
Measurement of Th2 cytokine production
Spleens were removed from naive BALB/c mice under aseptic conditions and homogenized in RPMI 1640 medium. The suspensions were then treated with 0.1 M Tris HCl (pH 7.2) containing 8 g/L Tris ammonium chloride to lyse red blood cells and centrifuged at 200 g for 5 minutes at 4°C. Cell pellets were then washed three times in RPMI 1640 medium and resuspended in complete RPMI 1640 medium (containing 2 mM
Statistical analysis
Data are expressed as mean±SE values. SPSS version 12.0 software (SPSS, Inc., Chicago, IL, USA) was used for statistical analyses. Differences between groups were determined by one-way analysis of variance. A value of P<.05 was considered statistically significant.
Results
Effect of CAE on airway responsiveness
OVA sensitization and airway allergen challenges significantly enhanced airway hyperresponsiveness compared with control mice at methacholine concentration of 25–50 mg/mL. These increases in airway hyperresponsiveness were significantly suppressed in a dose-dependent manner by CAE. Treatment with 30 mg/kg CAE once daily was more potent than treatment with dexamethasone at any concentration of methacholine (Fig. 1).
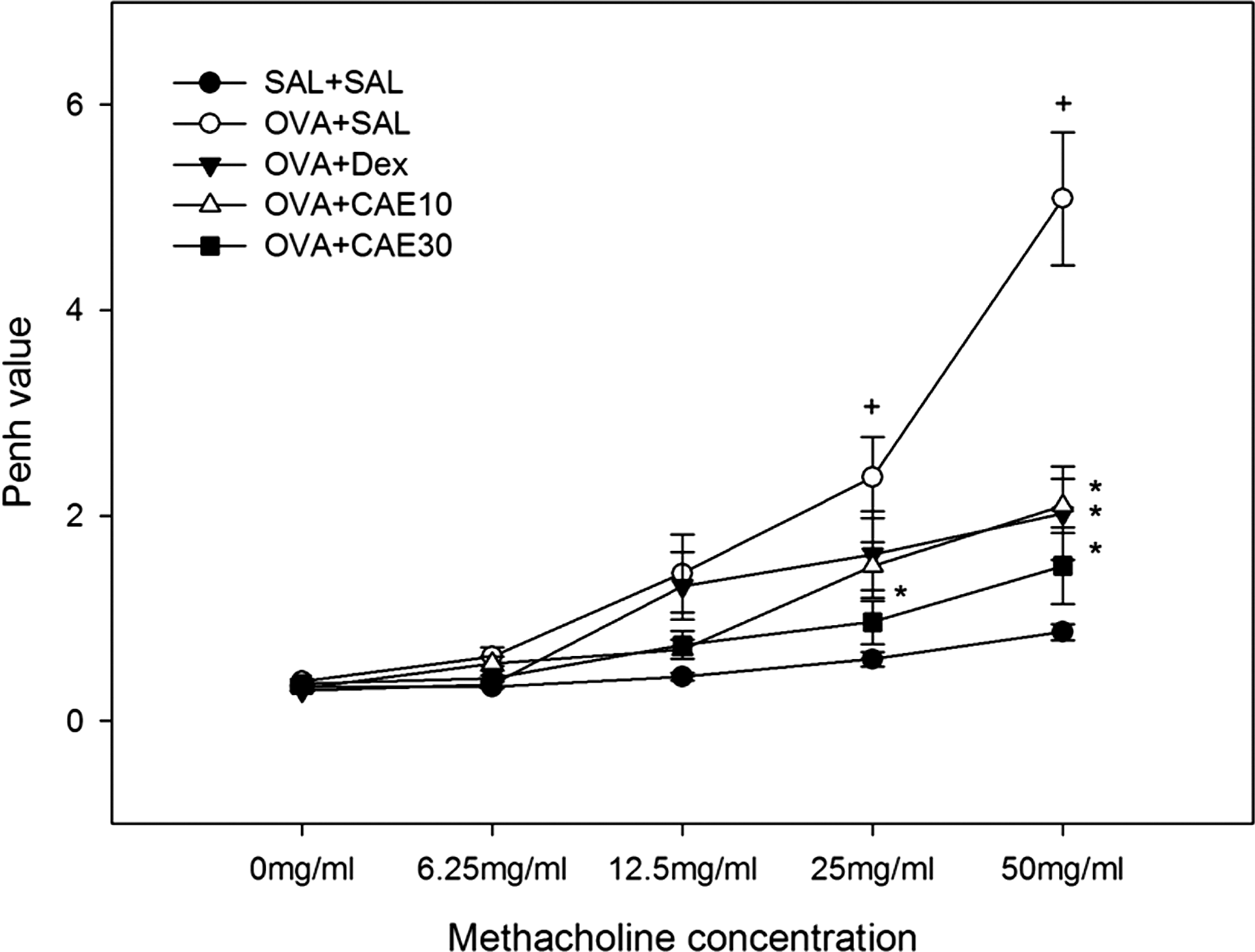
C. annuum L. extract (CAE) treatment attenuates development of airway hyperresponsiveness in ovalbumin (OVA)-sensitized and challenged mice. Airway responsiveness was measured 24 hours after the last challenge in control mice (SAL+SAL), OVA-sensitized and challenged mice treated with saline (OVA+SAL), OVA-sensitized and challenged mice treated with 3 mg/kg dexamethasone (OVA+Dex), and OVA-sensitized and challenged mice treated with CAE at 10 mg/kg (OVA+CAE10) or 30 mg/kg (OVA+CAE30). Enhanced pause (Penh) values were obtained in response to increasing doses of methacholine. Data are mean±SE values. + P<.05 versus SAL+SAL; *P<.05 versus OVA+SAL.
Effect of CAE on infiltration of eosinophils and other leukocytes into BAL fluid
To obtain evidence for inflammatory cell infiltration into lung tissue, we counted the number of total cells, lymphocytes, neutrophils, and eosinophils in BAL fluid in microscopic images. Infiltrated cells were increased significantly in OVA-sensitized and challenged mice compared with control mice (Fig. 2). The increases in the numbers of total cells, lymphocytes, neutrophils, and eosinophils were significantly reduced by oral administration of CAE. The inhibitory effect of CAE on the numbers of total cells, lymphocytes, neutrophils, and eosinophils in BAL fluid was greater at 30 mg/kg than at 10 mg/kg. The number of eosinophils infiltrating BAL fluid was significantly reduced in OVA-sensitized and challenged mice treated with 10 mg of CAE (95.92±7.43×103) or 30 mg/kg of CAE (86.07±10.11×103) compared with OVA-sensitized and challenged mice (330.55±58.22×103).
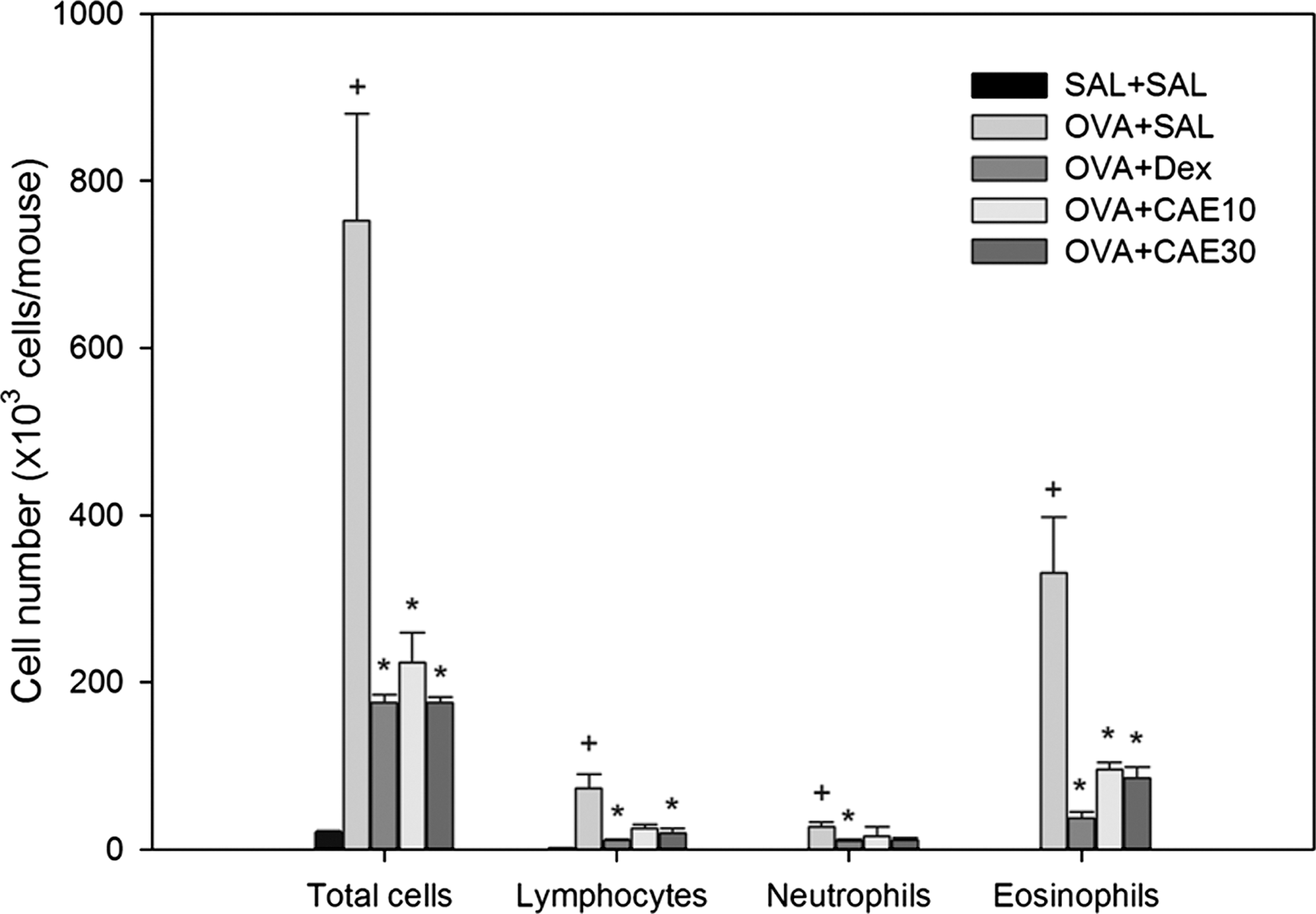
CAE reduces inflammatory cell infiltration in bronchoalveolar lavage fluid of OVA-sensitized and challenged mice. The numbers of total cells and differential cellular components of bronchoalveolar lavage fluid were determined in SAL+SAL, OVA+SAL, OVA+Dex, OVA+CAE10, and OVA+CAE30 mice 48 hours after the last challenge. Differential cell counts were performed in five different random locations to identify neutrophils, lymphocytes, and eosinophils. Data are mean±SE values. + P<.05 versus SAL+SAL; *P<.05 versus OVA+SAL.
Effect of CAE on ROS levels in BAL fluid cells
Recent evidence implicates oxidative stress as an important factor in aggravating lung diseases such as asthma and chronic obstructive pulmonary disease. OVA sensitization and challenge increased ROS levels in BAL fluid cells 2.18-fold compared with those in control mice. ROS levels were reduced by 55.08% in BAL fluid cells from OVA-sensitized and challenged mice treated with 30 mg/kg CAE compared with those from OVA-sensitized and challenged mice that received no drug treatment (Fig. 3).
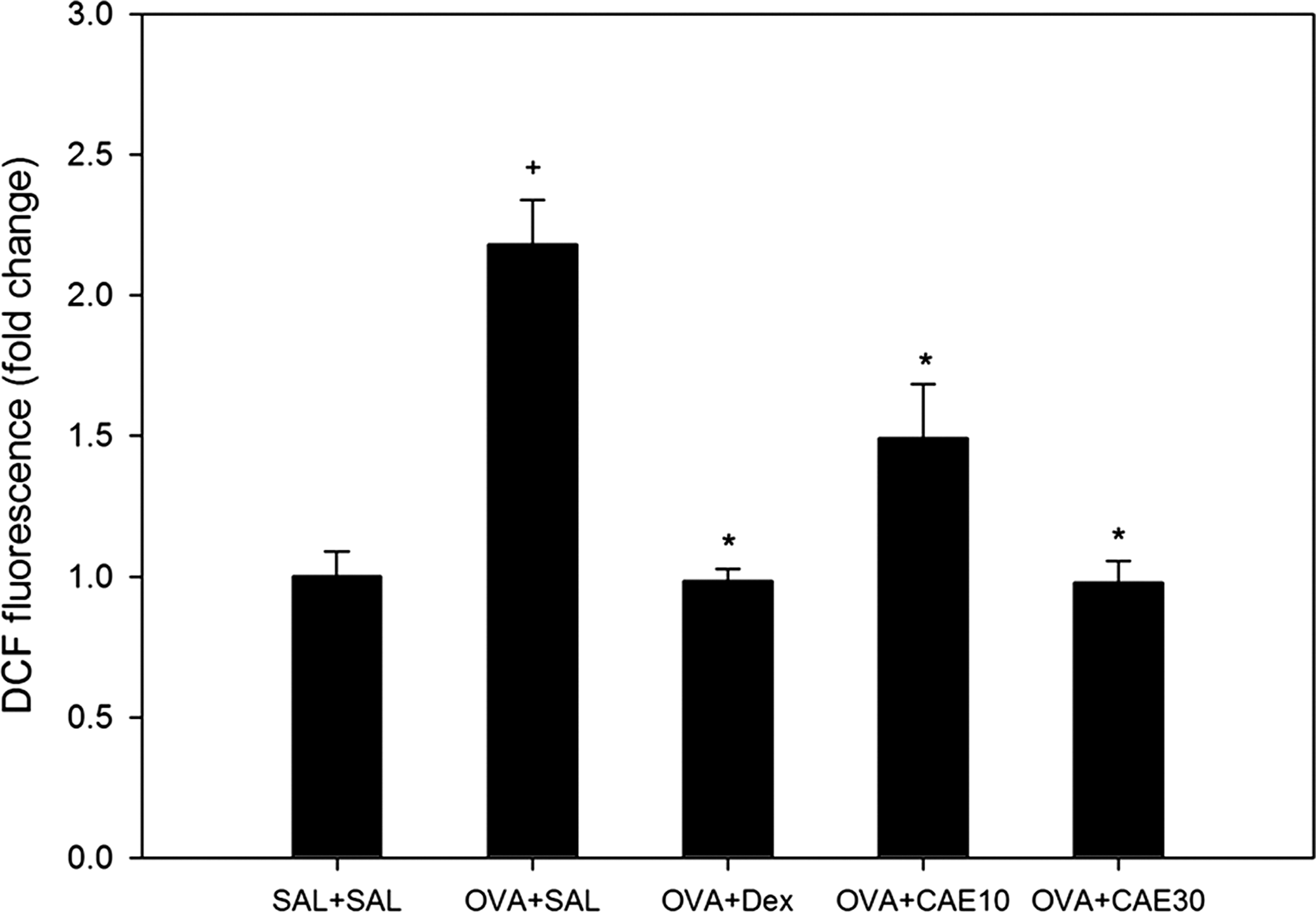
CAE reduces oxidative stress in bronchoalveolar lavage fluid cells of OVA-sensitized and challenged mice. Reactive oxygen species levels in bronchoalveolar lavage fluid cells were measured with 2′,7′-dichlorofluorescein (DCF) fluorescence in SAL+SAL, OVA+SAL, OVA+Dex, OVA+CAE10, and OVA+CAE30 mice. Data are mean±SE values. + P<.05 versus SAL+SAL; *P<.05 versus OVA+SAL.
Effect of CAE on total IgE and OVA-specific IgE levels in serum
Serum IgE and OVA-specific IgE levels, which were barely detectable in control mice, were significantly increased by OVA sensitization and challenges. Treatment with CAE reduced serum IgE to levels similar to dexamethasone treatment; however, there was no significant difference compared with those in OVA-sensitized and challenged mice. OVA-specific IgE levels in serum were significantly reduced by CAE treatment (Fig. 4).
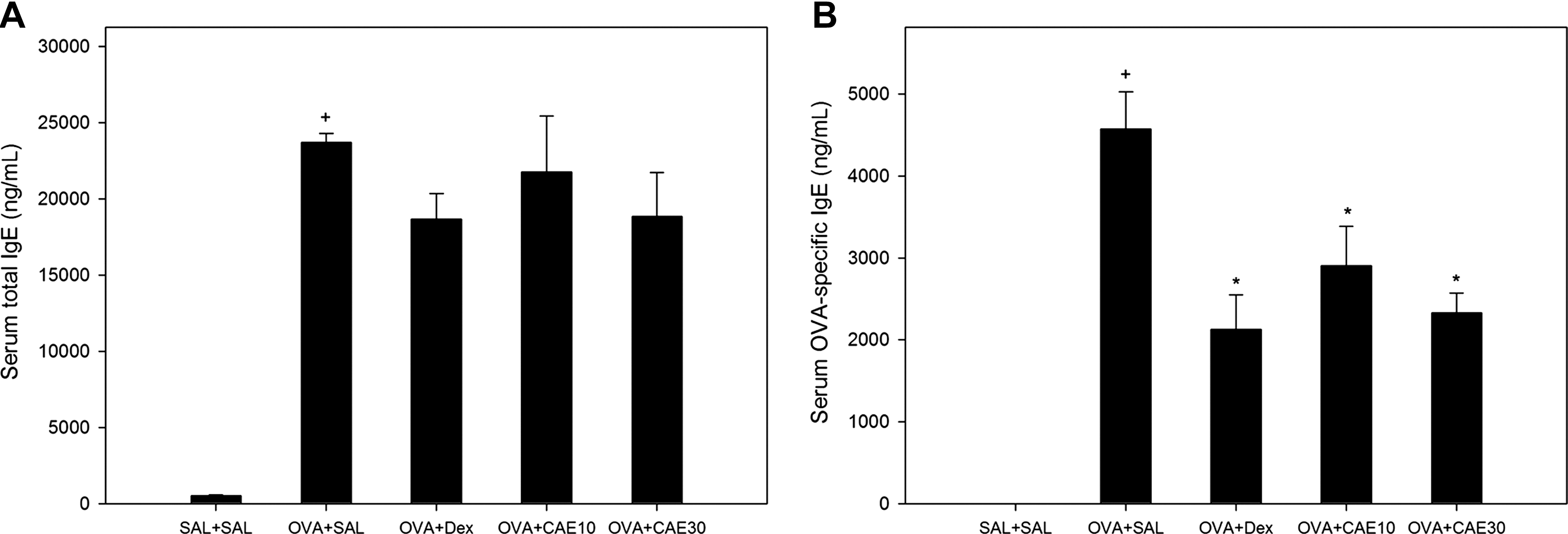
CAE treatment did not significantly reduce serum levels of total immunoglobulin E (IgE); however, levels of OVA-specific IgE in serum were significantly reduced in OVA-sensitized and challenged mice. (
Effect of CAE on Th2 cytokines
The levels of IL-4, IL-5, and IL-13 in BAL fluid were significantly increased in OVA-sensitized and challenged mice compared with those in control mice. Treatment with CAE significantly attenuated the increase in IL-4, IL-5, and IL-13 levels in BAL fluid compared with OVA-sensitized and challenged mice (Fig. 5A). Level of Th2 cytokines (IL-4, IL-5, and IL-13) were significantly increased following Con A stimulation of spleen cells from naive BALB/c mice. Con A–induced levels of IL-4, IL-5, and IL-13 were also significantly decreased in spleen cell cultures treated CAE. Treatment with CAE resulted in dose-dependent decrease of Th2 cytokines, with a maximum effect at 100 μg/mL treatment compared with control values (Fig. 5B).

CAE treatment significantly reduces the levels of T-helper type 2 (Th2) cytokines in bronchoalveolar lavage fluid and attenuates cytokine production from concanavalin A (Con A)–stimulated splenocytes. (
Effect of CAE on translocation of NF-κB p65 protein in lung tissues of OVA-sensitized and challenged mice
Western blot analyses revealed that the levels of NF-κB p65 protein in nuclear extracts from lung tissues were increased at 48 hours after OVA inhalation compared with the levels in control mice administered saline (Fig. 6). The OVA-induced increase in NF-κB p65 protein levels in nuclear protein extracts after inhalation was significantly attenuated by administration of CAE. In contrast, the levels of NF-κB p65 protein in cytosolic protein fractions from lung tissue were decreased after OVA inhalation compared with the levels in control mice (Fig. 6). NF-κB p65 protein levels in cytosolic fractions were substantially increased by administration of CAE. However, no significant changes were observed in OVA-sensitized and challenged mice treated with CAE.

CAE treatment reduced the levels of nuclear factor-κB (NF-κB) p65 protein in lung tissues of OVA-sensitized and challenged mice. NF-κB p65 protein levels were measured 48 hours after the last challenge. (
Effect of CAE on pathological changes and airway mucus production in OVA-induced asthma
Histological analyses revealed typical pathological features of asthma in the OVA-sensitized and challenged mice. Numerous inflammatory cells, including eosinophils, had infiltrated around the bronchioles, and mucus and debris had accumulated in the lumens of bronchioles (Fig. 7B and G) compared with control mice (Fig. 7A). Mice treated with CAE showed marked reductions in the infiltration of inflammatory cells in the peribronchial region (Fig. 7K). To ascertain effects on goblet cell hyperplasia and mucus secretion, we performed PAS staining. The percentage of airway epithelium that stained positively with PAS in OVA-sensitized and challenged mice (Fig. 7G) was significantly greater than that in control mice (Fig. 7F). Oral administration of CAE to OVA-sensitized and challenged mice significantly reduced the percentage of airway epithelium staining positively with PAS and the amount of debris in the airway lumens compared with that in OVA-sensitized and challenged mice (Fig. 7L).
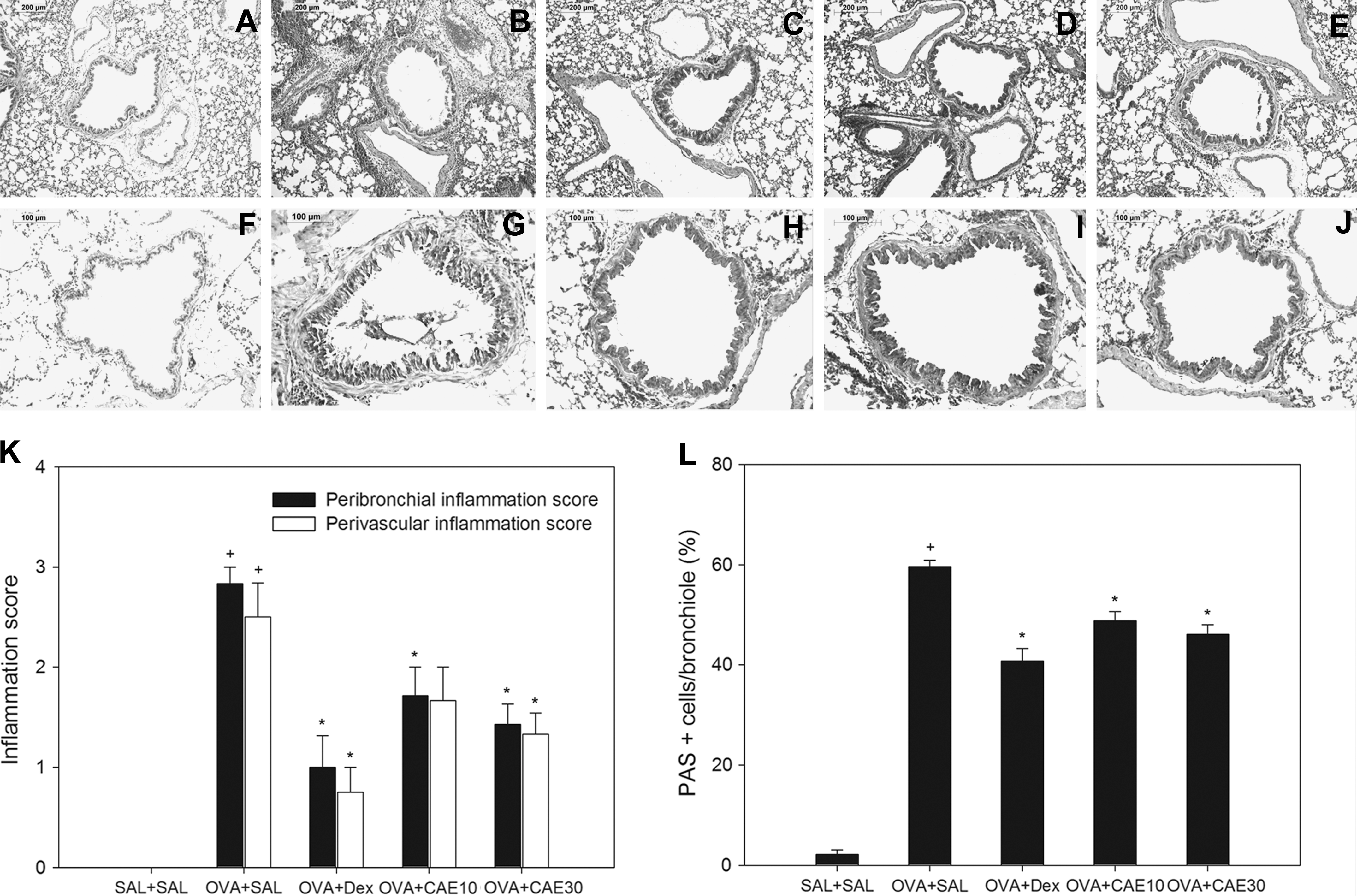
CAE treatment reduces lung tissue eosinophilia and mucus secretion in OVA-sensitized and challenged mice. Lung tissue was stained with (
Discussion
Previous studies have described a role for oxidative stress in asthma. Patients with asthma exhibit increased production of ROS, including superoxide anion (O2 −), hydrogen peroxide, and hydroxyl radicals (OH−). 1 Inflammatory cells recruited to the asthmatic airways may be a major contributor to the production of ROS. Activation of eosinophils, neutrophils, monocytes, and macrophages by many causes, including allergen exposure, can generate superoxide radical (O2 −) via the membrane-associated NADPH-dependent complex. Thereafter, dismutation of O2 − gives hydrogen peroxide, which can interact with various intracellular compounds to produce ROS. 2,15 ROS are chemically very active and can cause oxidative damage to cellular components. It has been shown that ROS disturb the cellular redox homeostasis and induce inflammatory signals that activate transcription factors, such as NF-κB, leading to the expression of cytokines, chemokines, and other inflammatory markers. 16,17 Consistent with these findings, our results showed that ROS generation in BAL cells, which mainly consist of recruited inflammatory cells, was increased significantly in our murine OVA-induced asthma model. Administration of CAE significantly attenuated both ROS production and inflammatory cell recruitment. Our results also have shown that CAE down-regulated the activity of the transcription factor, NF-κB. Protein levels of the p65 subunit of NF-κB in nuclear extracts were substantially decreased in OVA-sensitized and challenged mice treated with CAE, indicating that NF-κB activity was significantly reduced.
Th2 cytokines are also known to play an essential role in the pathogenesis of the allergic airway inflammation, and NF-κB is a critical transcription factor for Th2 cell differentiation. 15 IL-4, IL-5, and IL-13 can be produced by various lung resident cells, such as bronchial epithelial cells, tissue mast cells, and alveolar macrophages, as well as inflammatory cells, such as lymphocytes and eosinophils. Our results show that CAE significantly reduced the levels of IL-4, IL-5, and IL-13 in BAL fluids from OVA-sensitized and challenged mice. Similar findings were observed in Con A–stimulated splenocytes. These data show that the anti-inflammatory effect of CAE is at least in part mediated through a suppressive action on T lymphocytes. Therefore, the observed decreased IL-4, IL-5, and IL-13 levels in BAL fluid from CAE-treated mice may be due to inhibition of NF-κB activation in the infiltrated airway inflammatory cells. Elevated serum IgE levels are a hallmark of the Th2 immune response. IgE plays a significant role in the recruitment of eosinophils to the lungs of OVA-sensitized and challenged mice. 18 Our results showed that serum IgE levels were reduced by CAE treatment without significance. However, levels of OVA-specific IgE were significantly reduced by CAE treatment in OVA-sensitized and challenged mice.
It is believed that inflammation play a critical role in development of airway hyperresponsiveness. 19 It has been established that IL-5 is important in some animal models and almost certainly participates in the events leading to the eosinophilic inflammation and airway hyperresponsiveness in asthma. In addition, there is a close relationship among eosinophils, IL-4, IL-5, and IL-13 in the mechanism regulating airway hyperresponsiveness in the asthmatic lung. 19 Moreover, IgE-mediated mast cell activation may contribute to airway hyperresponsiveness by producing a wide array of inflammatory mediators and cytokines. 20 Thus the observed reduction of airway hyperresponsiveness by CAE may be associated with the reduction in Th2 cytokine production, tissue eosinophilia, and serum OVA-specific IgE level.
IL-4, IL-5, and IL-13 contribute to goblet cell hyperplasia and inflammatory cell infiltration into the airway. 21 Our findings showed that CAE prevented inflammatory cell infiltration into the airway as shown by a significant decrease in total cell counts and eosinophil and lymphocyte counts in BAL fluid and in tissue eosinophila in lung sections. We have also demonstrated a reduction in airway mucus production in CAE-treated mice. Selective ablation of NF-κB function in airway epithelium has been shown to reduce OVA-induced airway inflammation and mucus production in OVA-sensitized and challenged mice. 22
Our results are in agreement with a previous study that reported that capsaicin was effective against oxidative stress by countering the depletion of antioxidant molecules and antioxidant enzymes. 15,23 It is well documented that capsaicin, a main component of capsaicinoids in chili peppers (C. annuum L.) with potential anti-inflammatory and anti-tumor properties, also suppressed 12-O-tetradecanoylphorbol 13-acetate–induced activation of NF-κB in mouse skin, 24 in human promyelocytic leukemia HL-60 cells, 25 and human myeloid ML-1a cells. 26
On the basis of these observations, we have concluded that the inhibitory effect of CAE on oxidative stress in our murine model of allergic asthma was likely due, at least in part, to blocking the translocation of NF-κB into nuclei. We also found that treatment with CAE significantly attenuated major characteristics of allergic asthma, including airway hyperresponsiveness, airway inflammation, and elevated levels of Th2 cytokines in BAL fluid and lung tissues. Hence, these results indicate that CAE may provide a useful therapeutic approach for the treatment of allergic airway disease.
Footnotes
Acknowledgments
This work was supported by grant FGS1010924 from the Ministry of Education, Science and Technology and by grant KGM5231011 from the Korea Research Institute of Bioscience and Biotechnology Research Initiative Program of Korea.
Author Disclosure Statement
No competing financial interests exist.