
Abstract
In an attempt to identify bioactive natural products with anti-inflammatory activity, we evaluated the anti-inflammatory potential of the indole-containing fraction from the roots of Brassica rapa (IBR) (Family Brassicaceae) and the underlying mechanisms. Initially, we examined the inhibitory effect of IBR on the production of pro-inflammatory mediators in vitro and then evaluated its in vivo anti-inflammatory effects. IBR was found to concentration-dependently reduce the productions of nitric oxide, tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6) in lipopolysaccharide (LPS)-induced macrophages. Consistent with these findings, IBR suppressed the LPS-induced expressions of inducible nitric oxide synthase (iNOS) at the protein level and of iNOS, TNF-α, and IL-6 at the mRNA level. Furthermore, IBR attenuated LPS-induced DNA-binding activities of nuclear factor-κB (NF-κB), and this was accompanied by a parallel reduction in the degradation and phosphorylation of inhibitory κBα and, consequently, by a reduction in the nuclear translocation of the p65 subunit of NF-κB. In addition, treatment with IBR inhibited carrageenan-induced paw edema in rats and acetic acid–induced writing response in mice. Taken together, our data suggest that the expressional inhibitions of iNOS, TNF-α, and IL-6 caused by an attenuation of NF-κB activation are responsible for the anti-inflammatory and antinociceptive activity of IBR.
Introduction
I
Cytokines like TNF-α, IL-1β, and IL-6 have been reported to be crucial mediators of inflammation during acute response to injury. 9 TNF-α is induced by a wide range of pathogenic stimuli and performs a key role during the orchestration of inflammatory responses. 10 On the other hand, IL-1β is a major pro-inflammatory cytokine and is secreted by macrophages exposed to LPS. Acute inflammation induces the up-regulation of IL-1β at sites of inflammation, and increased levels of IL-1β can provoke cell or tissue damage. 11 IL-6 is a pivotal pro-inflammatory cytokine and a crucial checkpoint regulator of neutrophil trafficking, which is achieved by orchestrating chemokine production and leukocyte apoptosis. 12 Macrophages are the major cellular source of TNF-α, IL-1β, and IL-6, which in turn participate in the mediation of acute-phase responses to injury. 9
Several pathogenic factors cause the activation of nuclear factor-κB (NF-κB), which in turn controls the production of pro-inflammatory mediators. 13,14 Under normal conditions, NF-κB is a dimer composed of p65 and p50 and is present in the cytoplasm as a NF-κB–inhibitory κB (IκB) complex. Activation of NF-κB–IκB involves the phosphorylation of IκB and its subsequent recognition by ubiquitinating enzymes. The resulting proteasomal degradation of IκB leads to the translocation of the freed p65 and p50 subunits of NF-κB to the nucleus, where they bind to the consensus DNA binding sites to regulate the transcriptions of a large number of genes, which include those for pro-inflammatory cytokines, adhesion molecules, chemokines, and inducible enzymes. 15
Plants in the Brassicaceae family are important foods and account for a substantial portion of the worldwide vegetable production. Brassica rapa includes many significant crops, such as the Chinese cabbage and the turnip. In South Korea, B. rapa is mainly represented by the Ganghwa turnip and is used to make kimchi. 16 Brassica vegetables contain many active phytochemicals, such as isothiocyanates and indoles, the latter of which include indole-3-acetonitrile, indole-3-carbinol, and 3,3'-diindolylmethane. 17 In particular, indole-3-carbinol has potent anti-inflammatory and anticancer activities in vitro and in vivo, 18 and indole-3-acetonitrile inhibits the growth of insects and fungi and acts as a plant growth hormone because of its conversion in vivo to indole-3-acetic acid. 19,20 However, almost no information is available on the anti-inflammatory effects of B. rapa. Therefore, as part of our ongoing screening project to evaluate the anti-inflammatory potentials of natural compounds, we investigated the anti-inflammatory effects of the indole-containing fraction from the roots of B. rapa (IBR) on LPS-stimulated RAW 264.7 macrophages and subsequently in animal models.
Materials and Methods
Preparation of IBR and analysis of indole composition
The roots of B. rapa (28 kg) were extracted twice with 95% aqueous ethanol (40 L×2) at room temperature for 24 hours. The concentrated extracts were partitioned using an n-hexane and ethyl acetate mixture (1:1, 2.5 L×2, 19.7 g), ethyl acetate (3 L×2, 2.5 g), n-butanol (2.4 L×2, 33.0 g), and water (2 L), successively. From the n-hexane and ethyl acetate mixture fraction (1:1 fraction, indole-containing-fraction), two indoles—arvelexin (4-methoxyindole-3-acetonitrile) and caulilexin C—were isolated through repeated silica gel and ODS column chromatographies and identified. 21 High-performance liquid chromatography analysis was achieved using a Shimadzu (Tokyo, Japan) LC-20 instrument equipped with a Shimadzu SPD-20AV ultraviolet detector (268 nm) and Waters (Milford, MA, USA) C18 (250 mm×4.6 mm; particle size, 5 μm) column. The mobile phase consisted of water (solvent A) and methanol (solvent B), which were eluted at a flow rate of 0.7 mL/minute and with the following gradient elution: 0–20 minutes, 45–70% solvent B; 20–25 min, 90% solvent B; and 25–40 minutes, 90% solvent B. A 50-mg indole-containing fraction was dissolved in a mixture of pyridine (0.25 mL) and methanol (0.75 mL). A 20-μL aliquot was injected into the high-performance liquid chromatography system after filtration with a membrane filter (pore size, 0.45 μm). The two indoles, arvelexin and caulilexin C, were eluted at 16.7 minutes and 20.1 minutes, respectively. The contents of arvelexin and caulilexin C in the indole-containing fraction were determined to be 6.15 and 6.32 mg/g, respectively.
Materials
Dulbecco's modified Eagle's medium, fetal bovine serum, penicillin, and streptomycin were obtained from Life Technologies Inc. (Grand Island, NY, USA). Monoclonal antibodies to iNOS, cyclooxygenase (COX)-2, p65, phospho-IκB α, poly(ADP-ribose)polymerase (PARP), and β-actin and peroxidase-conjugated secondary antibody were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). The enzyme immunoassay (EIA) kits for TNF-α and IL-6 were obtained from R&D Systems (Minneapolis, MN, USA). Random oligonucleotide primers and M-MLV reverse transcriptase were purchased from Promega (Madison, WI, USA). SYBR® Green Ex Taq™ was obtained from TaKaRa (Shiga, Japan). iNOS, TNF-α, IL-6, COX-2, and β-actin oligonucleotide primers were purchased from Bioneer (Seoul, Korea). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), phenylmethylsulfonyl fluoride,
Cell culture and sample treatment
The RAW 264.7 macrophage cell line was obtained from the Korea Cell Line Bank (Seoul). These cells were grown at 37°C in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, penicillin (100 units/mL), and streptomycin sulfate (100 μg/mL) in a humidified 5% CO2 atmosphere. Cells were incubated with IBR at a concentration of 25, 50, or 100 μg/mL or with positive controls and then stimulated with LPS at 1 μg/mL for the indicated time. Various concentrations of tested compounds dissolved in dimethyl sulfoxide were added to the medium; the final concentration of dimethyl sulfoxide did not exceed 0.1%.
MTT assays for cell viability
RAW 264.7 macrophages were incubated with various concentrations (0–200 μg/mL) of IBR in the presence or absence of LPS (1 μg/mL) for 24 hours. After exposure to MTT solution for 4 hours at 37°C, the medium was discarded, and the formazan blue that formed in the cells was dissolved in dimethyl sulfoxide. The optical density was measured at 540 nm.
Lactate dehydrogenase cytotoxicity assays
RAW 264.7 macrophages were incubated with various concentrations (0–200 μg/mL) of IBR or Triton X-100 (1%) for 24 hours. Lactate dehydrogenase (LDH) levels in culture medium were determined spectrophotometrically using LDH cytotoxicity assay kits (TaKaRa Bio).
Nitrite determination
RAW 264.7 macrophages were plated at 5×105 cells per well in 24-well plates and then incubated with or without LPS (1 μg/mL) in the absence or presence of various concentrations (25, 50, or 100 μg/mL) of IBR for 24 hours. Nitrite levels in culture medium were determined using the Griess reaction assay and presumed to reflect NO levels. 22
Prostaglandin E2, TNF-α, and IL-6 assay
RAW 264.7 macrophages were pretreated with IBR for 1 hour and then stimulated with LPS (1 μg/mL) for 24 hours. Levels of prostaglandin (PG) E2, TNF-α, and IL-6 in the culture medium were quantified using EIA kits (R&D Systems).
COX-2 enzyme activity assay
Human recombinant COX-2 proteins were incubated with various concentrations (25, 50, or 100 μg/mL) of IBR for 10 minutes at 37°C before initiating the reaction by the addition of arachidonic acid. The reaction was terminated with a mixture of 0.1 N HCl and stannous chloride to convert all the PGE2 to PGF2α. The prostanoid products were quantified via EIA using a broadly specific antibody that binds to all the major prostaglandin compounds using the COX inhibitor screening assay from Cayman (Ann Arbor, MI, USA).
Western blot analysis
RAW 264.7 macrophages were collected by centrifugation and washed once with phosphate-buffered saline. The washed cell pellets were resuspended in PRO-PREP™ protein extraction solution (Intron Biotechnology, Seoul) and incubated for 20 minutes at 4°C. Cell debris was removed by microcentrifugation, followed by quick freezing of the supernatants. The protein concentration was determined using the Bio-Rad (Hercules, CA, USA) protein assay reagent according to the manufacturer's instruction. Cellular protein from treated and untreated cell extracts was electroblotted onto a polyvinylidene difluoride membrane following separation on 10–12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis. The immunoblot was incubated overnight with blocking solution (5% skim milk) at 4°C, followed by incubation for 4 hours with a primary antibody. Blots were washed four times with Tween 20/Tris-buffered saline and incubated with a 1:1,000 dilution of horseradish peroxidase–conjugated secondary antibody for 2 hours at room temperature. Blots were again washed three times with Tween 20/Tris-buffered saline and then developed by enhanced chemiluminescence (Amersham Life Science, Piscataway, NJ, USA). Density ratios versus β-actin or PARP were determined by densitometric analysis (Quantity One® software, Bio-Rad).
Quantitative real-time reverse transcriptase polymerase chain reaction
Total cellular RNA was isolated by Easy Blue® kits (Intron Biotechnology). From each sample, 1 μg of RNA was reverse-transcribed using MuLV reverse transcriptase, 1 mM deoxyribonucleotide triphosphates, and 0.5 μg/μL oligo(dT12–18). Polymerase chain reaction (PCR) amplification was performed using the incorporation of SYBR Green. The oligonucleotide primers for iNOS designed from the mouse (NM_010927) were CATGCTACTGGAGGTGGGTG (forward) and CATTGATCTCCGTGACAGCC (reverse), those for TNF-α designed from the mouse (NM_013693.2) were AGCACAGAAAGCATGATCCG (forward) and CTGATGAGAGGGAGGCCATT (reverse), those for IL-6 (NM_031168) were GAGGATACCACTCCCAACAGACC (forward) and AAGTGCATCATCGTTGTTCAT ACA (reverse), and those for COX-2 designed from the mouse (NM_011198) were TGCTGTACAAG CAGTGGCAA (forward) and GCAGCCATTTCCTTCTCTCC (reverse); the appropriate sizes of synthesized cDNA were 209, 220, 142, and 149 bp, respectively. The oligonucleotide primers for β-actin, used as a housekeeping gene, designed from the mouse were (NM_007393) were ATCACTATTGGCAACGAGCG (forward) and TCAGCAATGCCTGGGTACAT (reverse); the appropriate size of the synthesized cDNA was 200 bp. Steady-state mRNA levels of iNOS, TNF-α, IL-6, COX-2, and β-actin were determined by real-time quantitative PCR using the TaKaRa Bio Dice® thermal cycler. The results were expressed as the ratio of optimal density to β-actin.
Nuclear extraction and electrophoretic mobility shift assay
RAW 264.7 macrophages were plated in 100-mm-diameter dishes (1×106 cells/mL), treated with IBR (25, 50, and 100 μg/mL), stimulated with LPS for 1 hour, washed once with phosphate-buffered saline, scraped into 1 mL of cold phosphate-buffered saline, and pelleted by centrifugation. Nuclear extracts were prepared as described previously.
22
Nuclear extracts (5 μg) were mixed with double-stranded NF-κB oligonucleotide. 5′-AGTTGAG
Transient transfection and luciferase assay
The mouse iNOS promoter plasmid (pGL3-iNOS; −1,592/+185) and COX-2 promoter plasmid (pGL3-COX-2; −965/+39) were prepared as described previously. 23,24 RAW 264.7 macrophages were co-transfected with pGL3-iNOS or pGL3-COX-2 plasmid plus the phRL-TK plasmid (Promega) using Lipofectamine LTX™ (Invitrogen, Carlsbad, CA, USA) as instructed by the manufacturers. After 4 hours of transfection, cells were pretreated with IBR for 1 hour and then stimulated with LPS (1 μg/mL) for 18 hours. Each well was washed with cold phosphate-buffered saline, cells were lysed, and the luciferase activity was determined using the Promega luciferase assay system.
Animals
All animal experiments followed the university guidelines of the Ethical Committee for Animal Care and Use of Kyung Hee University (Seoul) according to an animal protocol (KHP-2010-03-8). ICR male mice weighing 20–25 g and Sprague–Dawley male rats weighing 180–200 g were purchased from Orient Bio Inc. (Seongnam-si, Korea) and maintained under controlled conditions (temperature of 20±2°C, humidity of 40–60%, and a light/dark cycle of 12 hours). At 12 hours before the experiment, only water was provided.
Carrageenan-induced edema in rats
The initial hind paw volume of the Sprague–Dawley strain rats was determined volumetrically. A 1% solution of carrageenan in saline (0.1 mL per rat) was injected subcutaneously into right hind paws after the test sample had been administered orally. The test samples were first dissolved in 5% ethanol and 5% Cremophor® EL (Sigma Chemical Co., St. Louis, MO, USA) and diluted with saline. The control group received only vehicle. Paw volumes were measured for up to 5 hours after injections, and edema volumes were measured using a plethysmometer. Ibuprofen (100 mg/kg), an anti-inflammatory drug, was used as a positive control.
Acetic acid–induced abdominal constriction in mice
The acetic acid abdominal constriction test was performed using the Atta-ur-Rahman method 25 with minor modifications. Vehicle, IBR (25 or 100 mg/kg), or ibuprofen (100 mg/kg), a reference peripheral analgesic, was orally administered 1 hour before the experiment, and 0.7% acetic acid in saline at 10 mL/kg was then injected intraperitoneally. The frequencies of abdominal constrictions per animals were then counted over the next 10 minutes starting 10 minutes after the injection.
Statistical analysis
Results are expressed as mean±SD values of triplicate experiments. Statistically significant values were compared using analysis of variance and Dunnett's post hoc test, and P values of <.05 were considered statistically significant.
Results
Anti-inflammatory activity and cytotoxicity of solvent fraction prepared from B. rapa
In the present study, the ethanol extract of B. rapa was successively partitioned with n-hexane–ethyl acetate, ethyl acetate, and n-butanol. The n-hexane–ethyl acetate fraction (IBR) showed the most potent inhibitory effects against LPS-induced NO and TNF-α production, whereas it showed low cytotoxicity in RAW 264.7 macrophages (Table 1). Therefore, IBR was chosen to ensure the anti-inflammatory activity of B. rapa as the most active fraction. The potential cytotoxicity of IBR on RAW 264.7 macrophages was investigated by measurement of the cell viability using the MTT assay with increasing concentrations of IBR (0–200 μg/mL) during a 24-hour period in the presence or absence of LPS. No effect on cell viability was observed up to 200 μg/mL (Fig. 1A). In addition, we measured the total LDH release in cell culture medium after treatment with IBR for 24 hours. LDH content was slightly, but not significantly, increased at concentrations up to 200 μg/mL IBR (Fig. 1B), indicating that its suppressive effects on NO and TNF-α production could not be attributable to nonspecific cytotoxicity.
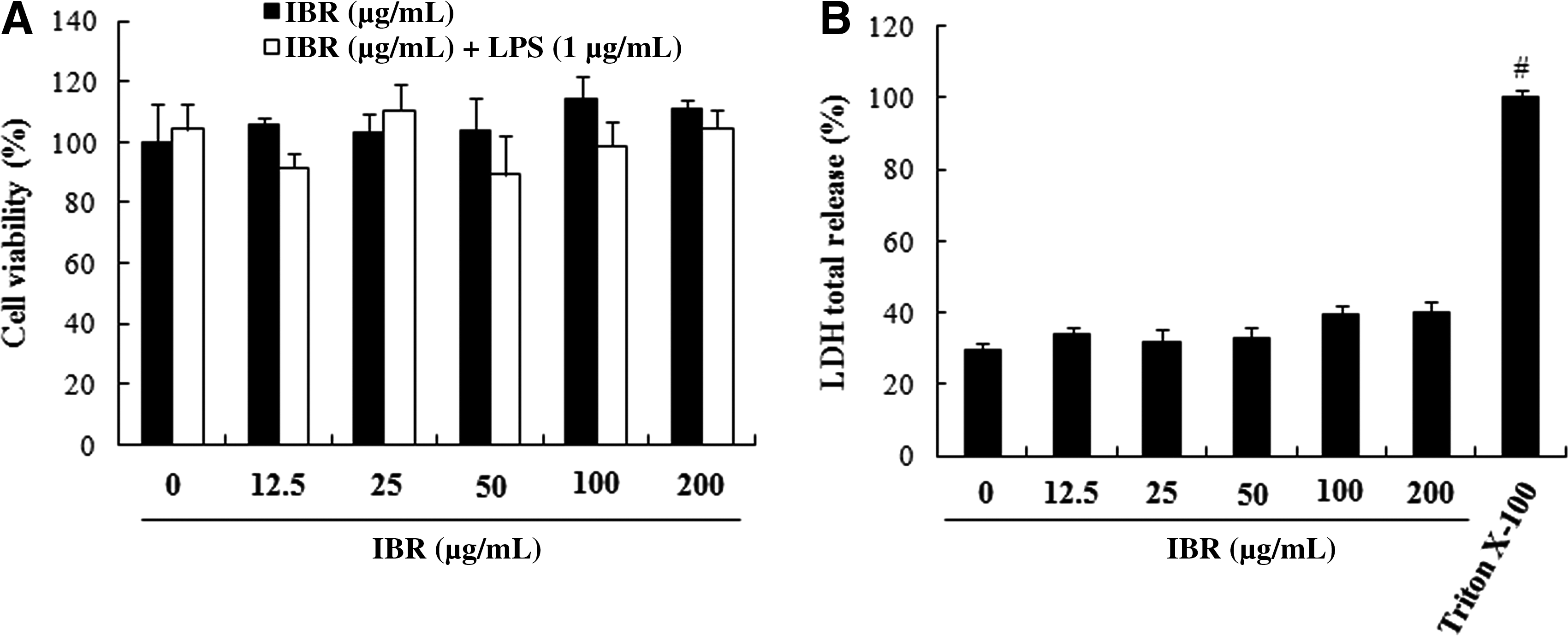
Effects of the indole-containing fraction from the roots of B. rapa (IBR) on (
IC50, the concentration required for 50% inhibition; NO, nitric oxide; TNF-α, tumor necrosis factor-α.
IBR inhibited LPS-induced NO production and iNOS expression
To investigate the effects of IBR on LPS-induced NO production in RAW 264.7 macrophages, culture media were harvested, and nitrite levels were measured. IBR (25, 50, or 100 μg/mL) potently and dose-dependently inhibited LPS-induced NO production (50% inhibitory concentration, 37.59±4.25 μg/mL) (Fig. 2A).
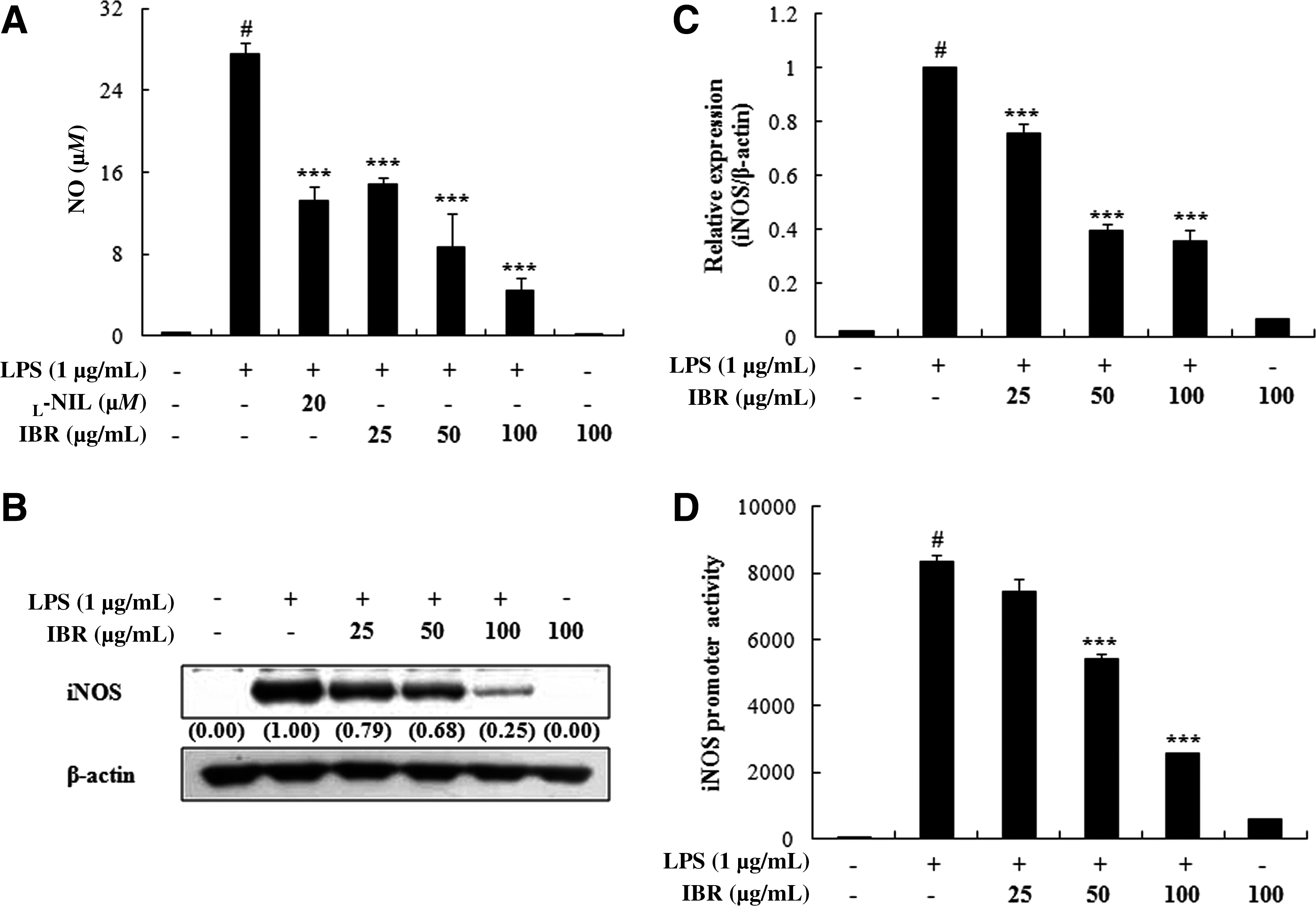
Effects of IBR on (
IBR inhibited LPS-induced TNF-α and IL-6 release and their mRNA expressions
To examine the effects of IBR on pro-inflammatory cytokine production, we examined its effect on LPS-induced TNF-α and IL-6 production and their mRNA expressions in LPS-induced RAW 264.7 macrophages using EIA and quantitative reverse transcriptase PCR, respectively. Pretreatment of RAW 264.7 cells with IBR considerably reduced LPS-induced TNF-α and IL-6 production (50% inhibitory concentration, 66.53±6.82 μg/mL and 54.90±2.17 μg/mL, respectively) and their mRNA expressions (Fig. 3). Collectively, the above findings show that IBR inhibits the LPS-induced expressions of iNOS, TNF-α, and IL-6 at the transcriptional level.

Effects of IBR (
IBR inhibited LPS-induced PGE2 production but had no effect on COX-2 mRNA expression
To assess the effects of IBR on LPS-induced PGE2 production in RAW 264.7 macrophage cells, PGE2 production was measured using an EIA. Stimulation of cells with LPS (1 μg/mL) resulted in a significant increase in PGE2 production compared with unstimulated control cells (Fig. 4A). However, pretreatment with IBR markedly inhibited this LPS-induced PGE2 production (42.7±2.05% at 100 μg/mL). In this experiment, NS-398 (3 μM) was used as a positive control to block PGE2 production (Fig. 4A). In addition, we investigated whether the inhibitory effects of IBR on PGE2 production involved the expressional modulation of COX-2, and thus we quantified COX-2 protein and mRNA levels by western blot and quantitative reverse transcriptase PCR, respectively. In unstimulated RAW 264.7 cells, COX-2 protein and mRNA levels were undetectable, but in response to LPS, COX-2 was obviously expressed at both the protein and mRNA levels. However, although IBR pretreatment significantly inhibited COX-2 protein expression, it did not affect the mRNA expression and promoter activity of COX-2 (Fig. 4C and D). To investigate whether the inhibitory effects of IBR on PGE2 production were via inhibition of COX-2 activity, the COX-2 enzyme activity was determined using recombinant proteins and inhibited by IBR in a concentration-dependent manner (Fig. 4E). These results implied that the suppression on PGE2 production by IBR is mediated via COX-2 regulation at the posttranscriptional and/or translational stages and its enzyme activity.
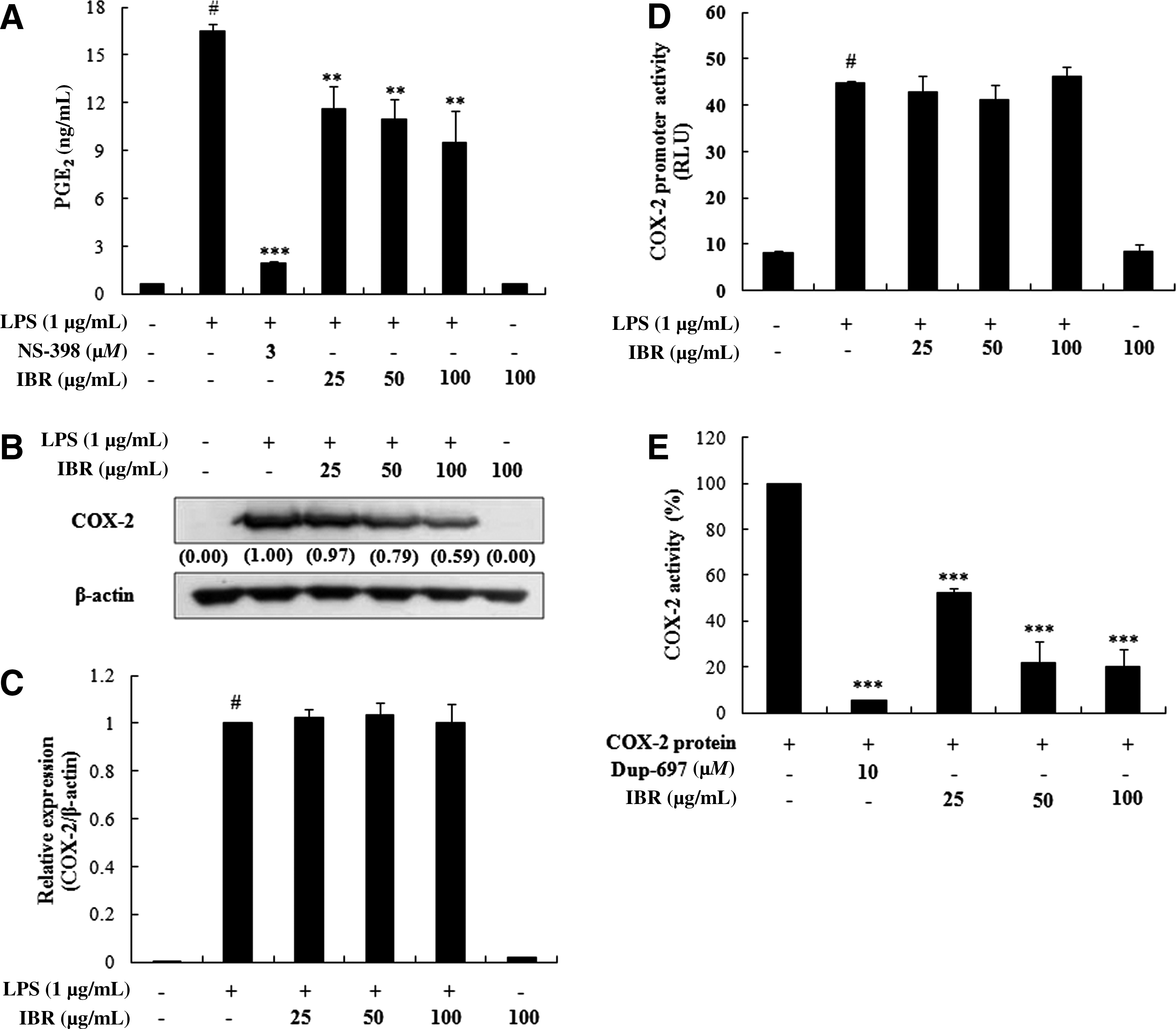
Effects of IBR on (
IBR reduced LPS-induced NF-κB-DNA binding activities and the nuclear translocation of NF-κB by blocking IκBα degradation
Because NF-κB activation is required for the expressions of iNOS, TNF-α, and IL-6 by LPS, 27,28 electrophoretic mobility shift assays were performed to determine whether IBR suppresses NF-κB activation. For these assays, nuclear extracts were obtained from RAW 264.7 macrophages stimulated with LPS (1 μg/mL) for 1 hour in the presence or absence of IBR. Treatment with LPS increased the binding activity of NF-κB to its consensus DNA sequence. However, pretreatment with IBR for 1 hour reduced this LPS-induced NF-κB-DNA binding (Fig. 5A). We next investigated whether IBR prevents the translocation of the p65 subunit of NF-κB to the nucleus by western blotting. It was found that LPS markedly induced the translocation of p65 to the nucleus and that pretreatment with IBR significantly suppressed this process (Fig. 5B). In addition, IBR pretreatment reduced the LPS-induced phosphorylation and subsequent degradation of IκBα, which inhibited NF-κB release from IκBα (Fig. 5C).
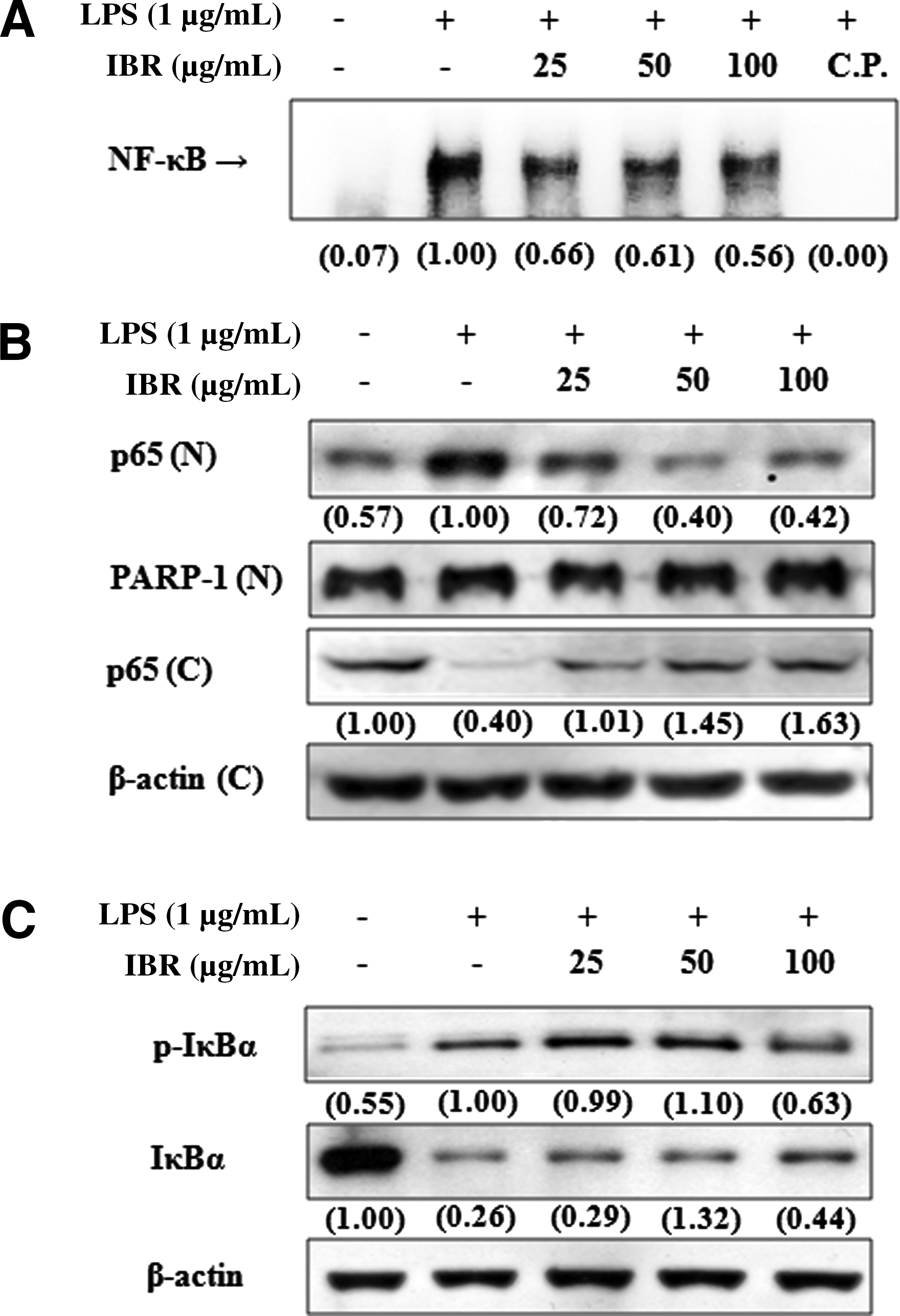
Effects of IBR on (
In vivo anti-inflammatory and antinociceptive activities of IBR
Because IBR was found to inhibit the inductions of iNOS and pro-inflammatory cytokines in macrophages, we sought to determine whether IBR affects acute-phase inflammation in animal models. In this study, we used the carrageenan-induced paw edema model because it is widely used to screen potential anti-inflammatory drugs. 29 Maximal edema formation was observed at 3 hours after a 1% carrageenan injection. However, IBR pretreatment (100 mg/kg, p.o.) reduced edema by 31.9% at 5 hours after carrageenan treatment, whereas the positive control, ibuprofen (100 mg/kg, p.o.), reduced edema by 69.9% (Fig. 6A). In addition, the antinociceptive properties of IBR were investigated on acetic acid–induced abdominal constriction in ICR mice. IBR significantly reduced the number of abnormal muscle constrictions by 18.2% at 25 mg/kg to 43.4% at 100 mg/kg against vehicle-treated controls (Fig. 6B). We also found that ibuprofen treatment (100 mg/kg, p.o.) as a positive control also effectively reduced carrageenan-induced edema and acetic acid–induced writhing.

(
Anti-inflammatory effect of isolated compounds from B. rapa
In a previous study we isolated the indole compounds arvelexin and caulilexin C from B. rapa and showed their inhibitory activity on human acyl-coenzyme A. 21 To determine if arvelexin and caulilexin C contribute to the anti-inflammatory effects of IBR, we investigated the effects of these compounds on the production of NO and PGE2 in LPS-induced RAW 264.7 macrophages (Fig. 7). It was found that arvelexin potently reduced the LPS-induced productions of NO and PGE2 inflammatory mediators in LPS-stimulated RAW 264.7 macrophages, whereas caulilexin C showed weak inhibitory effects.

Effects of arvelexin (AR) and caulilexin C (CA) on production of (
Discussion
The present study demonstrates for the first time the anti-inflammatory activities of IBR both in vitro, in LPS-stimulated RAW 264.7 macrophages, and in vivo, in the carrageenan-induced rat paw edema model. We found that IBR inhibited inducible NF-κB activation and the subsequent induction of pro-inflammatory mediators, such as iNOS, TNF-α, and IL-6, in RAW 264.7 macrophages and that it relieved carrageenan-induced edema formation in rats.
Several studies have indicated that ethanolic extracts of B. rapa prevent high-fat diet-induced obesity, 30 modulate the deleterious effects of diabetes, 16 and protect against cisplatin-induced nephrotoxicity. 31 During our continued work for screening anti-inflammatory activities, the ethanolic extract of B. rapa was found to inhibit the LPS-induced productions of NO, PGE2, TNF-α, and IL-6 (authors' unpublished data). Furthermore, activity-guided fractionation by anti-inflammatory activity against murine macrophages led to the isolation of an n-hexane–ethyl acetate fraction of B. rapa, which contained two indole acetonitriles, arvelexin and caulilexin C (see Materials and Methods). During preliminary testing of the anti-inflammatory activities of these two compounds, arvelexin potently inhibited the LPS-induced productions of NO and PGE2 more than caulilexin C in LPS-stimulated RAW 264.7 macrophages. According to our recently experiments, arvelexin exerts anti-inflammatory properties by down-regulating the IκB kinase–dependent NF-κB activation and consequent increase of pro-inflammatory mediators in LPS-induced RAW 264.7 macrophages. Moreover, arvelexin significantly reduced the septic lethality and serum levels of NO and cytokines in mice challenged with LPS. 32 These results suggested that anti-inflammatory activities of arvelexin at least contribute to the activity of IBR.
The pharmacologically induced down-regulations of LPS-inducible inflammatory mediators (for example, NO, TNF-α, and ILs) are regarded as an essential requirement for the alleviation of several disorders attributed to macrophage activation. Thus, RAW 264.7 macrophages provide us with an excellent model for anti-inflammatory drug screening and for the subsequent evaluations of the inhibitors of the pathways that lead to the inductions of pro-inflammatory enzymes and to the productions of pro-inflammatory cytokines. Our findings show that IBR inhibited the LPS-induced expressions and promoter activities of iNOS and the mRNA expressions of TNF-α and IL-6, indicating that IBR down-regulated the expressions of inflammatory genes at the transcriptional level. Recent evidence suggests that PGs are involved in inflammatory processes and that an inducible isoform of COX-2 is mainly responsible for the production of large amounts of PGs. 33 In the present study, IBR inhibited LPS-induced COX-2 at the protein level but barely affected its mRNA expression or promoter activity, which indicates that IBR affects COX-2 mRNA stability, COX-2 translation, or its protein stability. Accordingly, the molecular mechanism responsible for the decreased PGE2 production and COX-2 protein levels by IBR requires further investigation.
Because NF-κB regulates the expressions of pro-inflammatory enzymes, cytokines, and chemokines, it has been described as “a central mediator of immune response,” 34 and thus the NF-κB signaling cascade is an attractive therapeutic target for the development of treatments for inflammatory and autoimmune disorders. 35 In the present study, we investigated whether the anti-inflammatory activity of IBR is mediated by regulation of the NF-κB signaling pathway, and we found that IBR inhibited the LPS-induced DNA binding of NF-κB in RAW 264.7 macrophages. To identify the mechanism involved, we tested the effect of IBR on NF-κB activation signal. Western blotting revealed that IBR inhibited LPS-induced phosphorylation and degradation of IκBα and subsequently reduced nuclear levels of p65 in a concentration-dependent manner. These findings suggest that IBR inhibits NF-κB activation by suppressing IκBα phosphorylation and the translocation of p65 from the cytosol to the nucleus in LPS-induced RAW 264.7 macrophages.
Carrageenan-induced paw inflammation in the rat is a classical model of edema formation and hyperalgesia and has been extensively used in the development of anti-inflammatory drugs. Recent studies have shown that carrageenan induces the peripheral release of NO and PGE2, and NO is known to play a major role in edema formation and in the development of hyperalgesia during the inflammatory response and tissue injury. 36 In addition, it has been reported that carrageenan induces the release of TNF-α, which subsequently promotes IL-1 and IL-6 production in tissues. 37 This carrageenan-induced rat paw edema model demonstrated the ability of IBR to inhibit the edema induced by acute inflammation. Our results show that IBR markedly inhibited hind paw swelling in this model and that its inhibitory effect lasted for up to 5 hours after carrageenan injection. This result is consistent with the marked inhibitions of LPS-induced productions of NO, PGE2, TNF-α, and IL-6 productions by IBR in macrophages, and these results indicate that the anti-edema effects of IBR are due to its inhibition of NO, PGE2, TNF-α, and IL-6 syntheses in peripheral tissues. We also examined the antinociceptive effects of IBR using acetic acid–induced abdominal constriction model. Although acetic acid–induced pain (also called the abdominal constriction response or writhing) is a nonspecific model, it is widely used for the evaluation of peripheral antinociceptive activity. 38 The antinociceptive activities shown by IBR in this model indicated that IBR possesses peripherally mediated antinociceptive properties. However, we were not able to prove that the antinociceptive properties of IBR are centrally mediated. Acetic acid is an irritating agent that stimulates local peritoneal receptors to induce pain and characteristic abdominal constrictions when injected into the mouse peritoneal cavity (this effect has been associated with prostanoids, that is, with increased levels of PGE2 and PGF2α in peritoneal fluids). 39 Thus, the effects of IBR observed in the present study lead us to hypothesize that it probably inhibits PG synthesis.
In summary, we found that IBR inhibits the LPS-induced production of NO, TNF-α, and IL-6, which were coincidental with decreased expressions of iNOS, TNF-α, and IL-6 at the transcriptional level; however, the expression of COX-2 was not affected by IBR. Furthermore, these inhibitions were found to result from the prevention of NF-κB activation via IκBα degradation in RAW 264.7 macrophages. In addition, we found that IBR exhibited anti-inflammatory activity against carrageenan-induced paw edema and analgesic activity against nociceptive responses triggered by an intraperitoneal acetic acid injection. Accordingly, it appears that the anti-inflammatory and analgesic effects of IBR share a common molecular pathway probably involving the inhibition of pro-inflammatory mediators. Our findings suggest that IBR has the potential to ameliorate inflammatory disease associated with pain.
Footnotes
Acknowledgment
This research was supported by a grant from the GangHwa County for the Investigation of Biological Active Components and Evaluation of Pharmacological Efficacy in GangHwa Indigeous Crops.
Author Disclosure Statement
No competing financial interests exist.