
Abstract
An extract of Curcuma longa was tested in hypercholesterolemic rats to investigate its potential therapeutic effect on vascular conditions. Four experimental groups were used: normal diet (ND) control group, high cholesterol diet (HCD) group, and HCD subgroups supplemented with turmeric extract at 100 or 300 mg/kg of body weight (HCD100Tur and HCD300Tur groups, respectively). Turmeric extract was fed orally to animals, and dietary treatments lasted for 28 days. Hypercholesterolemia developed in the HCD, HCD100Tur, and HCD300Tur rats. Segments of the thoracic aorta were isolated, and an organ bath experiment was used to assess the vasorelaxation capability among all rats. Rats fed only HCD showed a marked decrease in acetylcholine-induced vasorelaxation compared with ND control rats. The HCD100Tur and HCD300Tur rats showed significant improvement in vasorelaxation compared with HCD rats. When vasorelaxation was induced by high concentrations of sodium nitroprusside, no differences in vasorelaxation were observed among the four groups of rats. A mechanistic study showed that HCD100Tur and HCD300Tur rats had significantly higher levels of the antioxidant enzymes superoxide dismutase and glutathione peroxidase than HCD rats. The transcript levels of heat shock protein 70 (hsp70), bcl2, bax-α, caspase (casp3), and glyceraldehyde 3-phosphate dehydrogenase in aortic tissues indicated that hypercholesterolemia significantly increased the expression of bax-α and casp3 but down-regulated bcl2 expression compared with the control group. Turmeric increased the expression of hsp70 and bcl2 but greatly reduced casp3 expression, indicating that turmeric improves vasorelaxation of the aorta in hypercholesterolemic rats by increasing antioxidant enzyme activities and likely suppressing apoptosis.
Introduction
V
Atherogenesis is commonly caused by the presence of free radicals, which mediate oxidative processes giving rise to specific products that aid in the onset of atherosclerosis. 7 One common consequence of atherosclerosis is the impairment of vasorelaxation, leading to consequences such as high blood pressure, acute ischemic stroke, and coronary heart disease. 8 –10 The number of deaths projected in 2030 from cardiovascular diseases is 23.6 million worldwide. 11 The search for drugs or functional foods that can prevent or even reverse cardiovascular disease in patients is therefore urgent.
Curcumin, an active compound in turmeric, has been reported to exert beneficial effects in patients having diseases such as digestive disorders, 12 inflammation, 13 diabetes mellitus, 14 and certain cancers. 15 –17 Recently curcumin has been shown to improve several aspects of cardiovascular health. Curcumin blocks homocysteine-induced endothelial dysfunction in porcine coronary arteries. 18 Healthy endothelial cells help prevent blood clotting and plaque formation and release nitric oxide, which promotes vasodilatation. Atherosclerotic lesions are decreased by curcumin treatment, resulting in a significant decrease in early atherosclerotic lesions (fatty streaks) in rabbits fed fat- and cholesterol-rich diets. 19 Curcumin also inhibits C-reactive protein–induced damage in vascular tissues, completely inhibiting the negative effects of C-reactive protein on these endothelial cells. 20
The objective of this study was to investigate the potential therapeutic values of turmeric extract on aorta relaxation and determine whether turmeric extract may serve as a functional food supplement to improve vascular health and protect vascular tissue from damage using rats with induced hypercholesterolemia as a model. In addition, the mechanism by which turmeric protects vascular tissues was investigated.
Materials and Methods
Dietary treatments
Twenty-four male Sprague–Dawley rats 6 weeks old were housed individually with 12 hours of alternating light and dark cycles for 28 days and randomly divided into four dietary groups: normal diet (ND) control group, high-cholesterol diet (HCD) group, and HCD supplemented with turmeric extract at 100 or 300 mg/kg/day (HCD100Tur and HCD300Tur, respectively). All rats were weighed each day, and the amount of food consumed was calculated throughout the experimental period. ND contained 14% protein, 10% fat, and 76% carbohydrate. HCD consisted of ND plus an additional 2% pure cholesterol, 1% cholic acid, and 5% oil (Guangdong Provincial Medical Laboratory Animal Center, Guangzhou, China). 21 Rats receiving the turmeric treatment (HCD100Tur and HCD300Tur rats) were orally administered the appropriate amount of turmeric extract suspended in double-distilled water according to their body weights, whereas vehicle-fed animals served as the placebo controls (ND rats). All rats had free access to water. The rats were fasted overnight at the end of the feeding period and euthanized the next day before dissection. Thoracic segments of the aorta of each rat were collected for further analysis. The protocols used in this animal study were approved by the Department of Health of the Government of Hong Kong and The Ethics Committee of the Hong Kong Polytechnic University.
Turmeric extract preparation
Turmeric from Sri Lanka was purchased at a local market and finely mashed with double-distilled water before filtering through layers of mesh. The residue was re-extracted twice with double-distilled water. The filtrate was cryodesiccated and stored at 4°C. Before use, the powder was resuspended in double-distilled water.
High-pressure liquid chromatography analysis of curcuminoids in turmeric extract
Standards were prepared by dissolving 50, 100, 250, 500, and 1000 ng of the curcuminoids bisdemethoxycurcumin (BDMC), demethoxycurcumin (DMC), and curcumin (95% purity; Changsha Nutramax Inc., Changsha, China) in high-performance liquid chromatography (HPLC)-grade methanol. The turmeric extract was suspended in methanol (1 mg/mL), sonicated for 10 minutes, filtered using Nylon-66 filter disks (pore size, 0.45 μm), and analyzed for curcuminoids using reverse-phase HPLC (Agilent 1100 HPLC System, Agilent Technologies, Waldbronn, Germany) as previously described. 22
Measurements of total cholesterol, HDL, LDL, and triglycerides in serum
Blood was collected by cardiac puncture and allowed to clot at 4°C for 2 hours after cervical dislocation. Serum was then collected and stored at −80°C. Serum total cholesterol (TC), triglycerides (TGs), LDL cholesterol (LDL-C), and HDL cholesterol (HDL-C) were then measured using an ALCYON® 300 analyzer (Abbott Laboratories, Abbott Park, IL, USA) using assay reagents (Triglycerides GPO-PAP, Cholesterol/HP, HDL-C plus 3rd generation, and LDL-C plus 2nd generation) purchased from Roche (Basel, Switzerland).
Calculation of atherosclerosis index
The atherosclerosis indexes (AIs) for the groups of rats fed with different diets were calculated using the formula AI=(TC – HDL-C)/HDL-C. 23 The AI has been used to indicate the risk of atherogenesis. 24
Vasorelaxation test on isolated thoracic aorta from rats fed different diets
Thoracotomy was performed rapidly on euthanized rats from each treatment group. The thoracic aorta segments were isolated and immediately immersed into ice-cold Krebs–Ringer (K-R) solution (135 mM NaCl, 5 mM KCl, 20 mM NaHCO3, 1.3 mM MgSO4·7H2O, 1.2 mM KH2PO4, 2.5 mM CaCl2, and 10 mM glucose). The aortic rings were cut into 3-mm ring segments and were fixed at their ends with two wires attached to force transducers connected to a PowerLab data acquisition system (ADInstruments, Sydney, Australia). The aortic rings were placed in organ baths containing K-R solution continuously supplemented with carbogen (5% CO2/95% O2) and contracted with 40 mM potassium chloride to determine vessel integrity. Basal tension of 1 g was applied to the vascular rings. The baths were then rinsed with K-R solution five times, and fresh K-R solution was replaced. When a steady relaxed state was attained, 1 μM phenylephrine was used to induce contraction of the aorta rings in the presence of 1 μM indomethacin and 1 μM neostigmine. After a steady contracted state was reached, acetylcholine was incrementally added to the K-R solution to obtain cumulative concentrations ranging from 10 nM to 3 mM. After each addition, the concentration of acetylcholine was maintained for 10 minutes to allow the tension to develop. The tension was measured using a force transducer, and the percentage of vasorelaxation attained using each acetylcholine concentration was calculated as ([TensionAcetylcholine – Basal Tension]/[Tensionat steady precontracted state – Basal Tension])×100.
Sodium nitroprusside–induced vasorelaxation test on isolated thoracic aortas
To test the sodium nitroprusside (SNP)-induced vasorelaxation, a steady state of contraction of aortic rings was achieved as described above. SNP was added gradually and cumulatively from 10 nM to 10 μM. For each SNP addition, 10 minutes was allowed for steady tension to develop. The percentage of vasorelaxation was calculated for each concentration of SNP.
Antioxidant enzyme measurement
Blood was collected into chilled centrifuge tubes by cardiac puncture immediately after cervical dislocation and allowed to clot for 2 hours at 4°C. The clotted blood was centrifuged at 540 g at 4°C for 10 minutes, yielding serum, which was stored at −80°C for subsequent antioxidant analyses. Superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GSH-PX) activities were measured using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The SOD, CAT, and GSH-PX activities are expressed as units per microliter of serum.
Reverse transcription–polymerase chain reaction
Total aorta RNA (2.5 μg) was used for reverse transcription (RT) of first-strand cDNA synthesis, which was then used for the subsequent polymerase chain reaction (PCR). The RT-PCR was carried out according to the manufacturer's protocol (Fermentas, Burlington, Canada). For each PCR, 1 μg of first-strand cDNA was used with 50 nM forward and reverse PCR primers. Sequence amplification was carried out with PCR Master Mix (Promega, Madison, WI, USA) and the primer sets listed in Table 1. Expression levels of the gene transcripts of interest—heat shock protein 70 (hsp70), bcl2, bax-α, and caspase (casp3)—were measured relative to that of a housekeeping gene (glyceraldehyde 3-phosphate dehydrogenase [gapdh]) to correct for differences in quantity and quality among different RNA samples. The PCR amplification protocol was as follows: denaturation for 3 minutes at 94°C, followed by 35 cycles of 1-minute denaturation at 94°C, 1-minute annealing at 55°C, and 1-minute extension at 72°C. The PCR products were resolved on a 0.8% agarose gel and stained with ethidium bromide. The light intensity of each band was measured using the freeware Image J (version 1.42) (National Institutes of Health, Bethesda, MD, USA). Gene expression levels were expressed as the ratio of the measured intensity of the genes of interest to the intensity of the gapdh gene.
casp3, caspase-3; gapdh, glyceraldehyde 3-phosphate dehydrogenase; hsp70, heat shock protein 70.
Statistical analysis
All data are expressed as mean±SD values. GraphPad (San Diego, CA, USA) Prism version 5.02 was used for curve fitting and data analysis. After validation of each parameter with respect to homogeneity of variance, differences among the groups were analyzed using a one-way analysis of variance followed by Bonferroni's test for multiple comparisons among means. A value of P<.05 was considered statistically significant.
Results
HPLC analysis of curcuminoids in turmeric extract
The curcuminoids were analyzed spectrophotometrically at 425 nm, and chromatograms of the turmeric extract are shown in Figure 1A. The three peaks at retention times of 9.86 minutes, 10.638 minutes, and 11.501 minutes were identified as BDMC, DMC, and curcumin, respectively, based on comparison of the absorption spectra with the standards (Fig. 1B) and previously published data. 25,26 The amount of each curcuminoid in the turmeric extract was determined by comparing the peak areas of the turmeric extract versus standard curves of the reference curcuminoids (data not shown). The total amount of curcuminoids measured in the water extract of turmeric was 7.58±0.25 mg/100 mg, slightly higher than that of some commercially available turmeric powders. 27 By weight, curcumin made up 4.56±0.22% of the turmeric extract, whereas BDMC and DMC made up 1.53±0.02% and 1.49±0.03% of the extract, respectively. Curcumin constituted 60% of the total curcuminoids. The HCD100Tur and HCD300Tur rats therefore received 7.6 and 22.7 mg of curcuminoids, respectively, containing 4.56 and 13.6 mg of curcumin/kg of body mass/day, respectively.
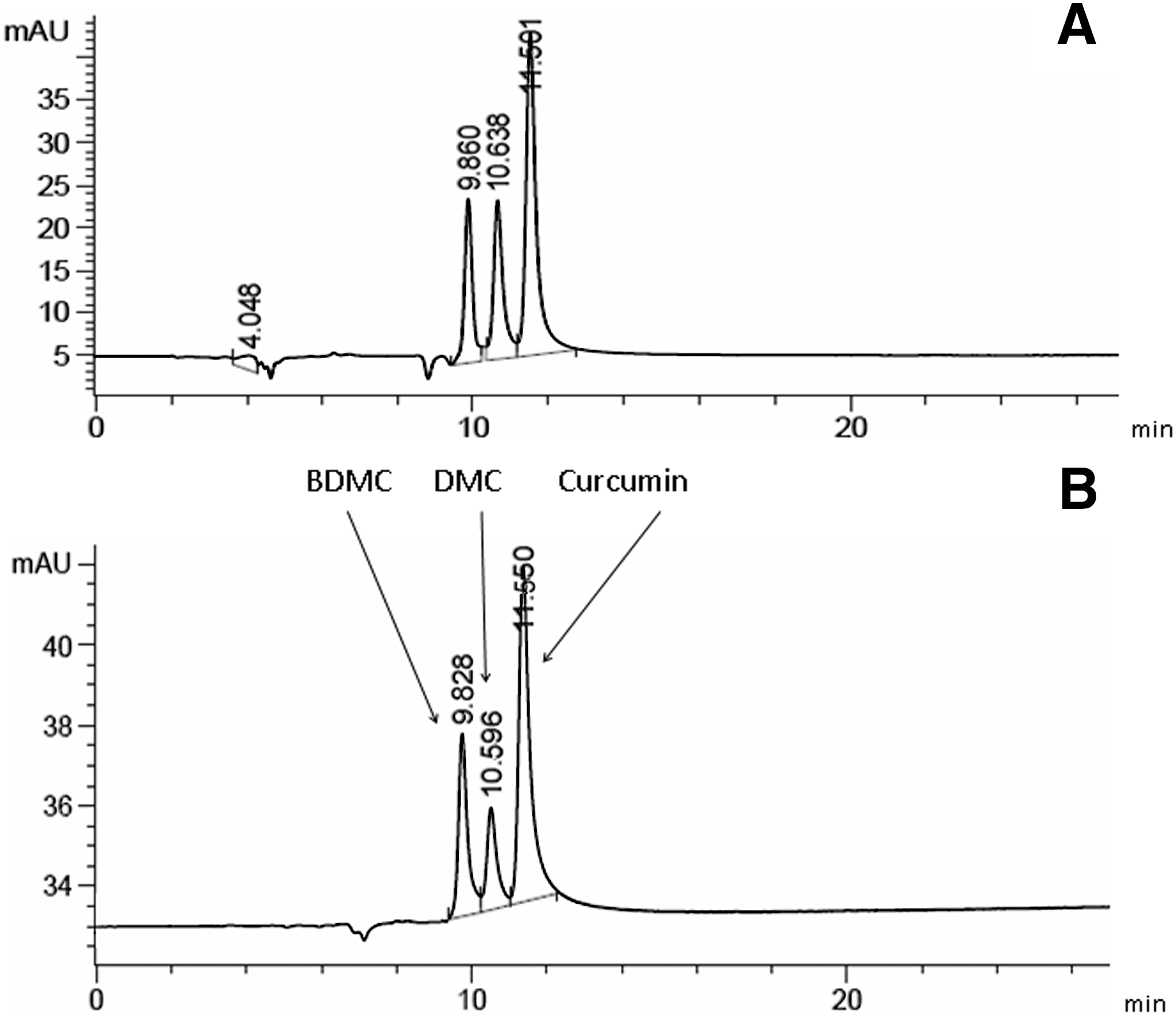
Chromatograms (425 nm) of
Effects of turmeric on TC, HDL-C, LDL-C, and TG concentrations in blood
The HCD rats developed hypercholesterolemia after 28 days on the HCD. On average, the HCD rats had significantly higher (93%) mean TC levels (4.27±0.42 mmol/L) than the ND group (2.21±0.12 mmol/L) (Table 2). In the HCD100Tur and HCD300Tur groups, the TC levels were 3.65±0.24 mmol/L and 3.23±0.22 mmol/L, respectively, representing a significant decrease of approximately 15–24% compared with the HCD group. However, these values were significantly higher than those for the ND rats (P<.05).
Plasma samples were prepared from rats fed with different diets (control normal diet [ND], high cholesterol diet [HCD], and HCD with turmeric extract at 100 or 300 mg/kg/day [HCD100Tur and HCD300Tur, respectively]), and the levels of total cholesterol (TC), triglycerides (TGs), high-density lipoprotein-cholesterol (HDL-C), and low-density lipoprotein-cholesterol (LDL-C) were measured. Data are mean±SD values (n=6).
The values obtained from the HCD rats were significantly higher than that of the ND rats (control) (P<.05).
The values obtained from the HCD100Tur rats and HCD300Tur rats were significantly higher than that of the ND rats (P<.05).
The values obtained from the HCD100Tur rats and HCD300Tur rats were significantly lower than those of the HCD rats (P<.05).
The values obtained from the HCD100Tur rats and HCD300Tur rats were significantly higher than those of the HCD rats (P<.05).
The value obtained from the HCD100Tur rats was significantly higher than that of the ND rats (P<.05).
HCD treatment, however, did not affect total serum TG levels in any of the four treatment groups. Similar results have been obtained by other researchers. 28
The mean HDL-C concentration in the ND rats was 1.91±0.21 mmol/L. There were no significant differences between the mean HDL-C concentrations of the HCD rats (1.71±0.16 mmol/L) and the ND rats. However, the HDL-C levels in both the HCD100Tur and HCD300Tur rats increased to 2.23±0.09 mmol/L and 3.03±0.23 mmol/L, respectively, representing increases of 17% and 59%, respectively, compared with the ND rats.
The ND and HCD rats had mean LDL-C concentrations of 0.48±0.14 and 2.64±0.81 mmol/L, respectively; therefore, the HCD value was approximately 5.5 times higher than the ND value. The HCD100Tur and HCD300Tur rats had mean LDL-C concentrations of 0.89±0.22 mmol/L and 0.47±0.11 mmol/L, respectively; these values were significantly lower (approximately 66% and 82%, respectively) than those of the HCD rats.
Antioxidant enzyme activities
The HCD rats had significantly lower levels of serum GSH-PX activity (179.2±18.6 U/mL) than the ND rats (297.2±62.6 U/mL, P<.05) (Fig. 2A). The mean GSH-PX activities measured in the HCD100Tur and HCD300Tur rats were 247.6±35.9 U/mL and 243.2±106.3 U/mL, respectively; these values were comparable to those observed in the ND group and significantly higher than those of the HCD group (P<.05).
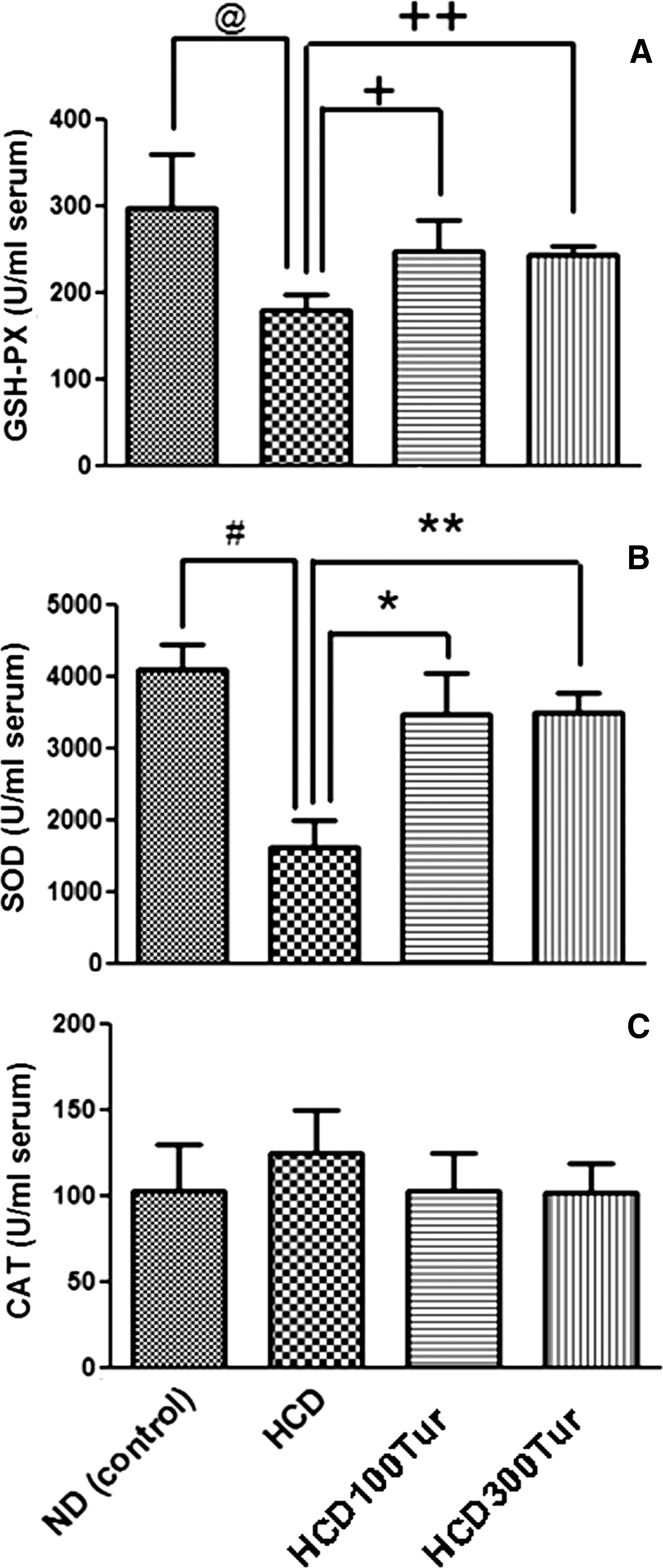
Effect of turmeric on activities of the antioxidant enzymes
Turmeric supplementation of the HCD group also increased SOD activity; the mean value of SOD activity in the ND group was 4099±348 U/mL (Fig. 2B), significantly higher than that of the HCD group (1620±368 U/mL, P<.05). The mean SOD activities in the HCD100Tur and HCD300Tur groups were 3461±587 and 3489±289 U/mL, respectively, which were significantly higher than that of the HCD rats (P<.05).
CAT activity was not significantly altered by the HCD or the turmeric extract (Fig. 2C). The mean CAT activities measured in all four groups were similar.
Turmeric extract improved vasorelaxation in rats fed HCD
A steady precontraction was induced in the thoracic aortic rings of all treatment groups using 1 μM phenylephrine before the aortas were exposed to various concentrations of acetylcholine (Fig. 3). All aortic rings showed a dose-dependent relaxation in response to acetylcholine. The ND rats' aortic rings had relaxed completely (100% vasorelaxation), and the aortic rings of the HCD100Tur and HCD300Tur rats exhibited 87% and 92% mean vasorelaxation, respectively, at 10 μM acetylcholine. Aortic rings prepared from HCD rats, however, only achieved approximately 82% relaxation, significantly lower than those from the control and HCD300Tur rats (P<.05). At 30 μM acetylcholine, the aortic rings of the HCD100Tur and HCD300Tur rats achieved 96% and 99% vasorelaxation, respectively, not significantly different from that of the ND control group but significantly higher than that of the HCD group (84%) (P<.05). Increasing the acetylcholine concentration further did not further relax the aortic rings of the HCD rats. These results show that HCD treatment probably caused tissue damage in the aortas and impaired their relaxation ability. Turmeric treatment could prevent this functional impairment, possibly by providing protection for aortic tissues under hypercholesterolemia.
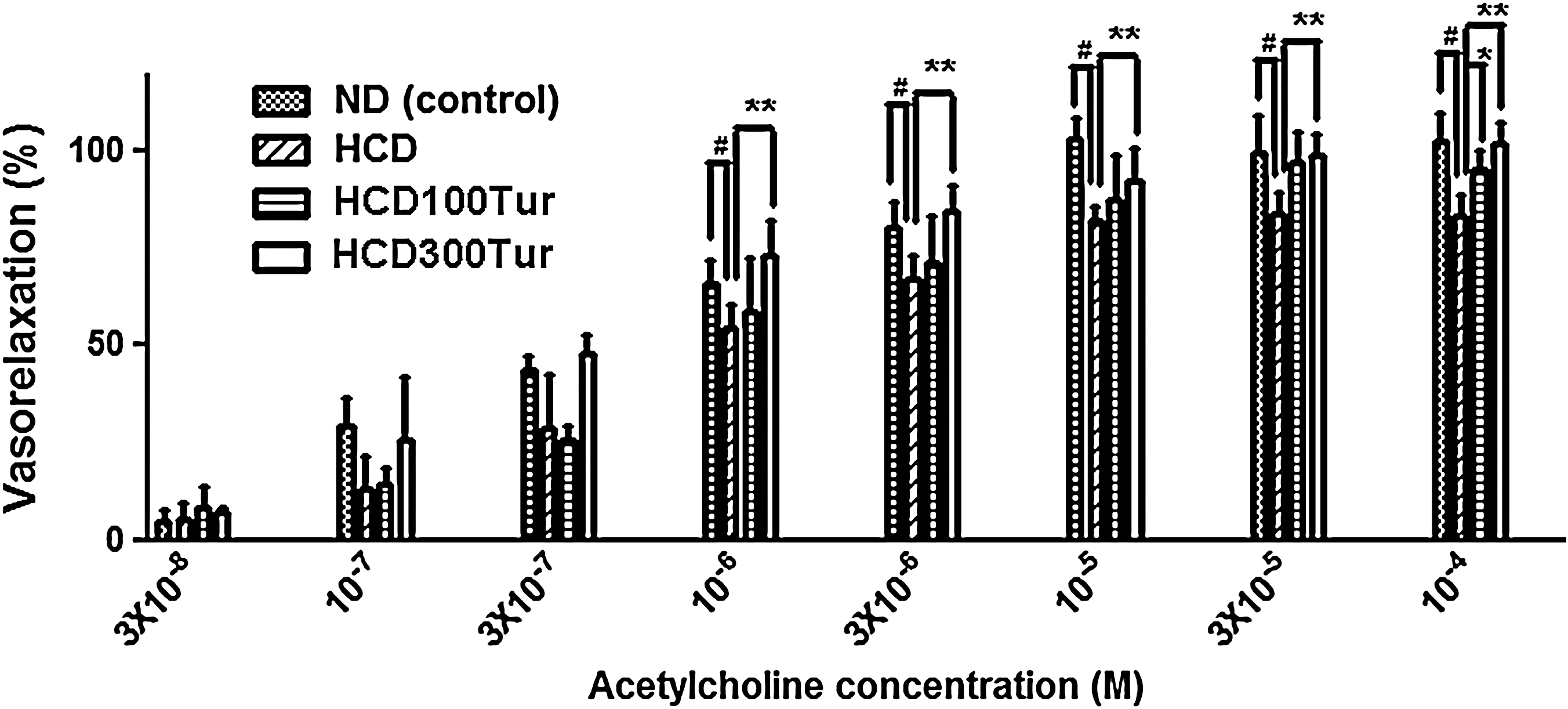
Measurements of acetylcholine-induced vasorelaxation in aorta rings prepared from rats receiving different diet treatments. #At 10−6 M, 3×10−6 M, 10−5 M, 3×10−5 M, and 10−4 M acetylcholine, the mean percentage of vasorelaxation measured in ND group rats' aorta rings (control) was significantly higher than the value obtained in the HCD group rats' aorta rings (P<.05). **At the same aforementioned concentrations of acetylcholine, the mean percentage of vasorelaxation measured in HCD300Tur group rats' aorta rings was significantly higher than that of the HCD group rats' aorta rings (P<.05). *At 10−4 M acetylcholine the HCDTur100 group rats' aorta rings had a significantly higher percentage of vasorelaxation compared with the HCD group rats' aorta rings (P<.05).
High concentrations of SNP induced full relaxation in HCD group aortic rings
To determine whether the significant decrease in vasorelaxation measured in HCD aortic rings was due to the release of insufficient nitric oxide by endothelial cells 29 or damage to the vascular smooth muscle cell layer of the aorta, exogenous SNP was added to the K-R solution in increasing doses to induce vasorelaxation (Fig. 4). At 10−8 M SNP, the percentage of vasorelaxation in the ND control rats was 35.3%, and this percentage was significantly lower (22.9%) in the HCD rats. The HCD300Tur rats had 37.4% vasorelaxation at the same concentration of SNP (10−8 M), and this value was significantly higher than that of the HCD rats (P<.05). At 10−7 M SNP, the HCD rats had significantly lower (69.6%) vasorelaxation than the ND (91.6%) or HCD300Tur (86.4%) rats (P<.05). At 10−6 M and 10−5 M SNP, all aortic rings showed complete relaxation, and there were no significant differences in the percentage of vasorelaxation among the groups. The vascular smooth muscle cells of the aortas were apparently not damaged because all aortic rings could relax fully at high concentrations of SNP. The decrease in vasorelaxation in the HCD rats, therefore, was most likely due to insufficient nitric oxide being present, suggesting that the endothelial cells of the thoracic aorta could have been damaged.
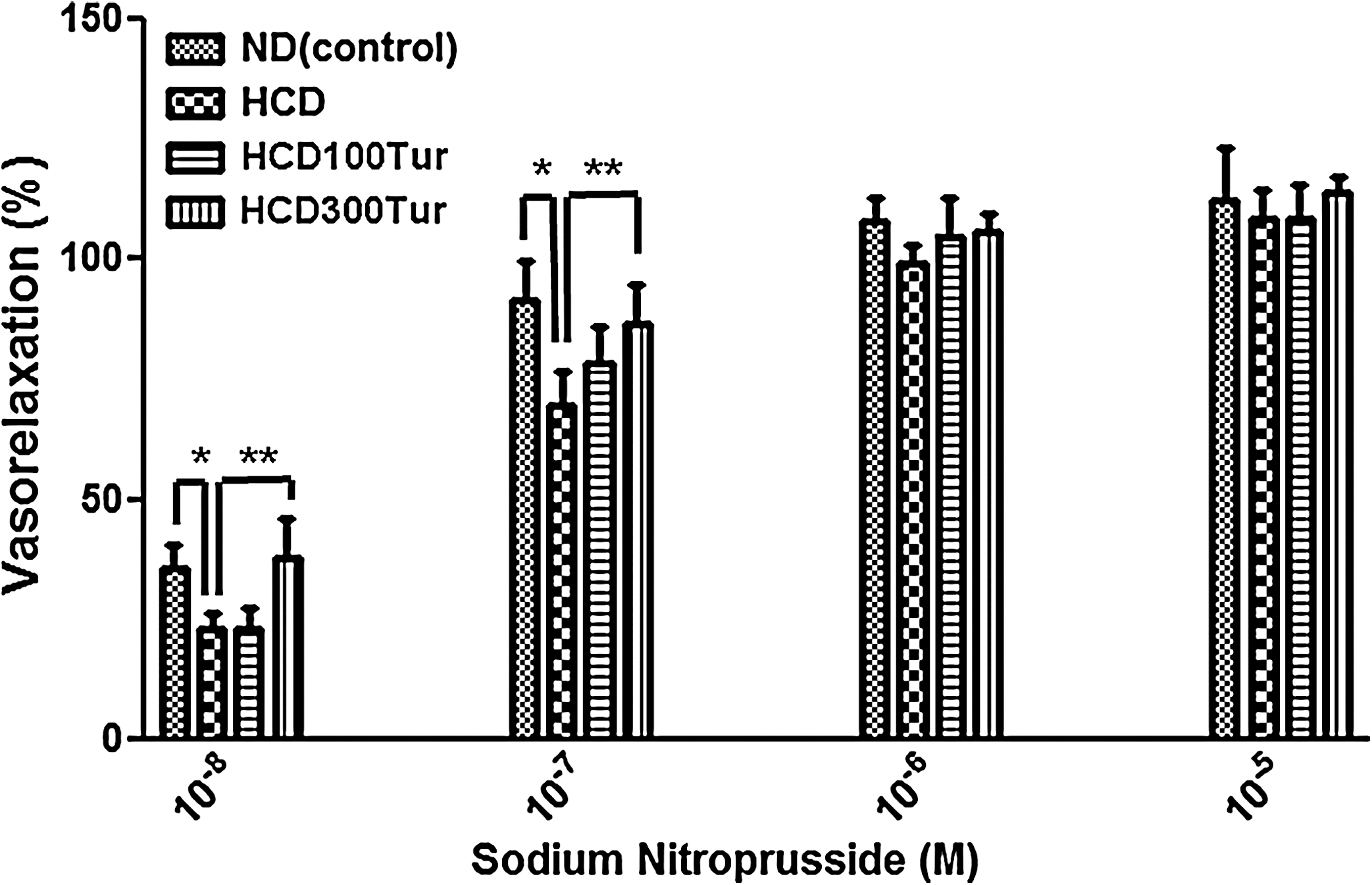
Measurements of vasorelaxation induced by sodium nitroprusside in aorta rings. *At 10−8 M and 10−7 M sodium nitroprusside, the mean percentage of vasorelaxation measured in ND group rats' aorta rings (control) was significantly higher than the value obtained in the HCD group rats' aorta rings (P<.05). **The HCD300Tur group rats' aorta rings had a significantly higher percentage of vasorelaxation induced by the same aforementioned concentrations of sodium nitroprusside compared with the HCD group rats' aorta rings. There were no significant differences in the percentage of vasorelaxation induced by 10−6 M and 10−5 M sodium nitroprusside among all groups of aorta rings.
Gene transcript levels of hsp70, bcl2, bax-α, and casp3
RT-PCR was used to determine the expression levels of various genes involved in apoptosis and the stress response in rat aortas under different dietary treatments (Fig. 5). The ratios of the expression levels of the genes of interest (hsp70, bcl2, bax-α, and casp3) to the internal reference gapdh were calculated and are summarized in Table 3. The hsp70:gapdh expression ratio in HCD rats was 0.87±0.03, significantly lower than the ratio of 0.98±0.01 in the ND rats (P<.05). The hsp70:gapdh expression ratios in the HCD100Tur and HCD300Tur rats, however, were 1.58±0.05 and 1.36±0.08, respectively, both of which were significantly higher than that of the ND rats (P<.05).
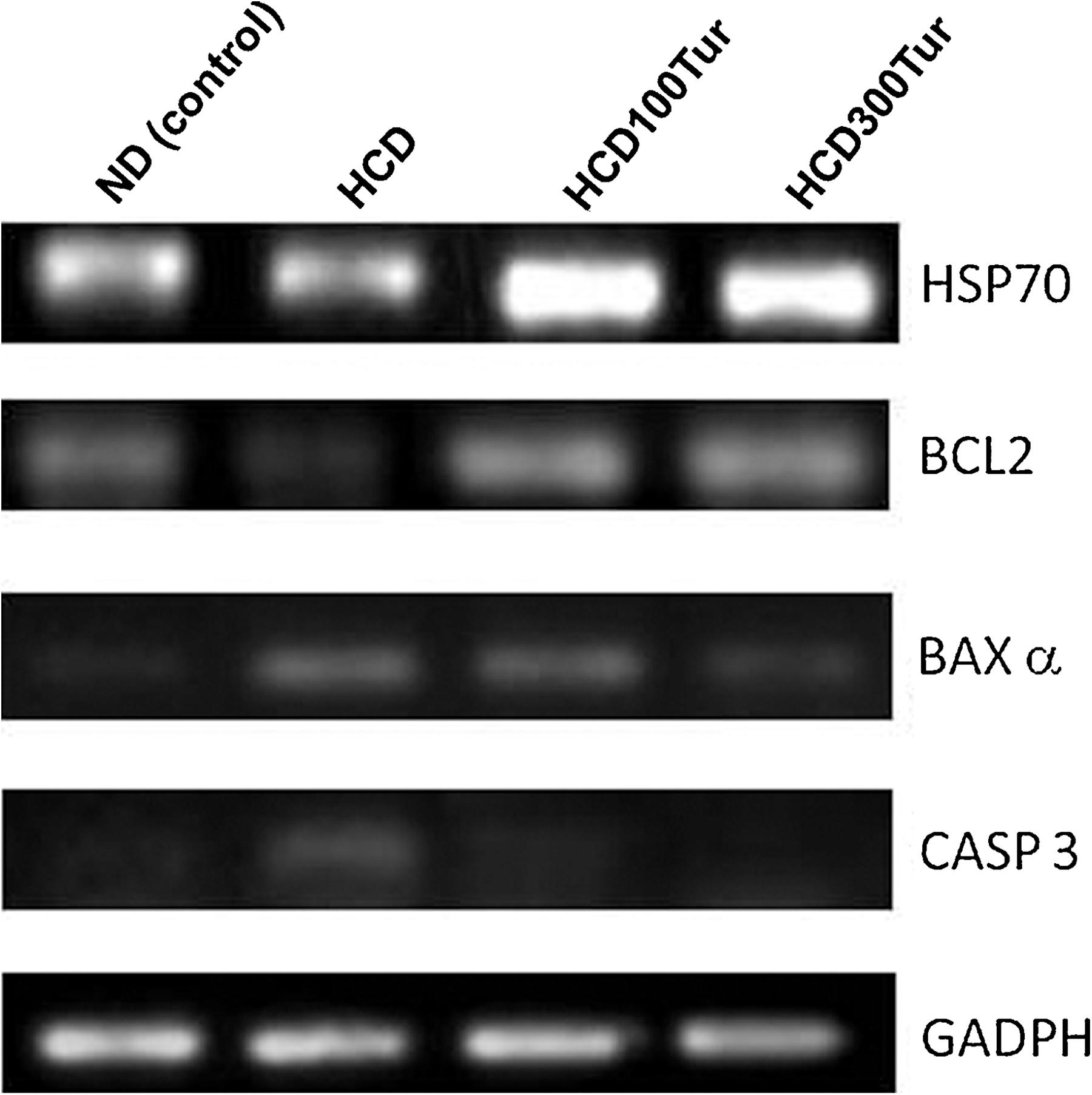
Effect of turmeric on transcriptions of hsp70, bcl2, bax-α, casp3, and gapdh genes in rat aorta. The reverse transcription–polymerase chain reaction products of genes of interest (HSP70, BCL2, BAXα, and CASP3) were electrophoresed on a 0.8% DNA agarose gel and stained with ethidium bromide. The light intensities of amplified DNA fragments under ultraviolet illumination were analyzed by using the free software Image J (version 1.42). GAPDH was used as an internal reference. The levels of gene transcripts are expressed as the ratio of light intensity of the gene of interest:light intensity of GAPDH. The calculated ratios are presented in Table 3.
The results of the reverse transcription–polymerase chain reaction suggest that turmeric can confer protection to rat aorta tissues by decreasing bax-α and casp3 expressions but increasing bcl2 and hsp70 expressions. Data are mean±SD values (n=6).
The values obtained from the HCD100Tur rats and HCD300Tur rats were significantly higher than that from the ND rats (control) (P<.05).
The values obtained from the HCD100Tur rats and HCD300Tur rats were significantly higher than that from the HCD rats (P<.05).
The value obtained from the HCD rats was significantly lower than that from the ND rats (control) (P<.05).
The value obtained from the HCD300Tur rats was significantly lower than that from the HCD rats (P<.05).
The value obtained from the HCD rats was significantly higher than that from the ND rats (control) (P<.05).
The value obtained from the HCD100Tur rats was significantly higher than that from the ND rats (P<.05).
The values obtained from the HCD100Tur rats and HCD300Tur rats were significantly lower than that from the HCD rats (P<.05).
The HCD rats had significantly lower anti-apoptotic gene bcl2 transcript levels in the aorta than did the ND rats. The mean bcl2:gapdh ratio calculated in the HCD rats was 0.23±0.01, significantly lower than the ratio in the ND rats (0.51±0.03, P<.05). In HCD100Tur and HCD300Tur rats, the bcl2:gapdh ratios were 0.68±0.02 and 0.79±0.03, significantly higher than the ratios in the ND or HCD rats (P<.05).
Transcription levels of the pro-apoptotic genes bax-α and casp3 were also measured. The mean ratio of bax-α:gapdh was 0.17±0.08 in the ND control rats and significantly higher in the HCD rats (0.49±0.05) (P<.05). The bax-α:gapdh ratios measured in HCD100Tur and HCD300Tur rats were 0.43±0.01 and 0.31±0.05, respectively, demonstrating that bax-α gene expression was down-regulated by turmeric.
The ND control rats had a mean casp3:gapdh ratio of 0.12±0.01. This ratio measured in HCD rats (0.36±0.02 [P<.05]) was significantly higher than in the ND control group. The increase in casp3 expression induced by HCD was suppressed when turmeric extract was supplemented to the rats' diet. The HCD100Tur and HCD300Tur rats had casp3:gapdh ratios of 0.18±0.01 and 0.13±0.03, respectively, significantly lower than the level in the HCD rats (P<.05).
Discussion
It has been reported that rats are resistant to developing atherosclerosis. 30 Diets containing 1% cholesterol and 0.5% cholic acid are capable of increasing plasma TC and LDL-C in rats without promoting atherogenesis. 31 Nevertheless, atherogenesis can still be induced in rats when they are fed a diet containing a higher percentage of cholesterol (2%) and cholic acid (1%). 32,33 The objective of our study was to investigate whether turmeric extract can improve vascular function in rats and protect vascular tissue from damage due to hypercholesterolemia (probably as the result of atherosclerosis). The diet chosen therefore contains higher concentrations of cholesterol and cholic acid, not only to induce hypercholesterolemia but also to ensure vascular tissue damage and vascular function impairment in the HCD-treated rats.
In our study, turmeric reduced plasma TC and LDL-C levels and increased plasma HDL-C levels. The AI in HCD rats was five times greater than that of the ND rats (Fig. 6), suggesting that the HCD rats had a much higher risk of developing atherosclerosis than the ND rats. The HCD100Tur and HCD300Tur rats, in contrast, had significantly lower AIs than the HCD rats, suggesting that turmeric supplementation in HCDs may significantly decrease the risk of developing atherosclerosis. Together with this reduction in the risk of atherogenesis, it seems reasonable to suggest that turmeric may lower the risk of developing cardiovascular diseases such as myocardial infarction, stroke, and aneurysm, 34,35 which are mainly caused by chronic development and progression of atherosclerosis.
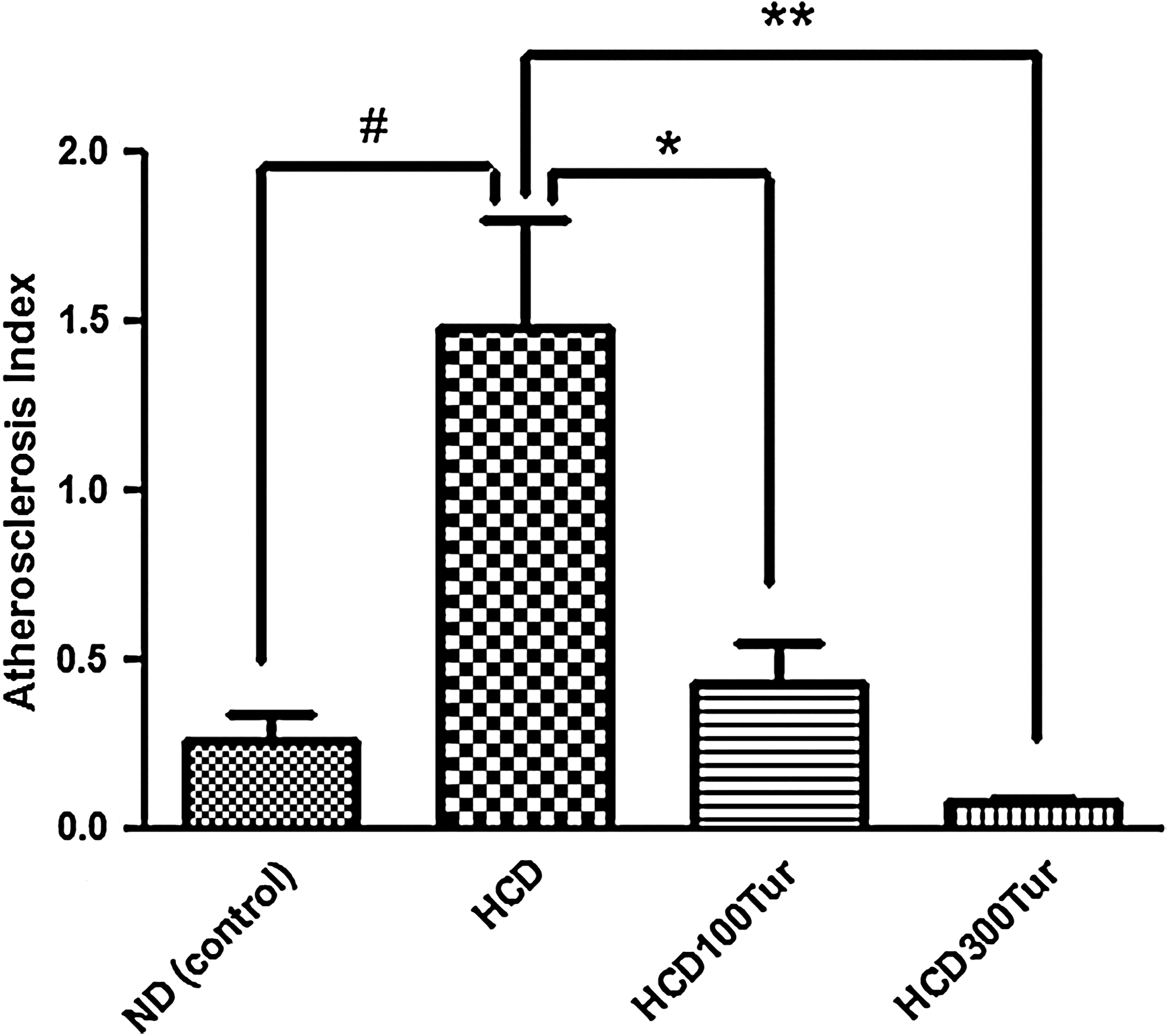
Atherosclerosis index calculated in rats receiving different diet treatments. #The value obtained from the HCD rats was significantly higher than that from the ND rats (control) (P<.05). *The value obtained from the HCDTur100 rats was significantly lower than that from the HCD rats (P<.05). **The value obtained from the HCD300Tur rats was significantly lower than that from the HCD rats (P<.05). Data are mean+SD values (n=6).
Vasorelaxation induced by acetylcholine was significantly lower in the HCD rats than in the control ND rats, possibly because of either endothelial cell damage, 36 which can lead to a decrease in nitric oxide release resulting in a decline in vasorelaxation, 37 or damage to the smooth muscle cells of the aorta. However, complete vasorelaxation was achieved in all treatment groups at high SNP concentrations (10−6 M and 10−5 M), indicating that there was no physical damage to the smooth muscles of the thoracic aorta. The smooth muscle of aortic wall from HCD rats, however, was less sensitive to the vasodilating effect of nitric oxide, likely because senescence developed in the smooth muscle cells in atherosclerotic lesions; 38 therefore high concentrations of SNP were required by aorta rings from HCD rats in order to have comparable degree of vasorelaxation to the control. Although aortas in HCD rats required high concentrations of SNP to vasorelax, aortas in HCD300Tur rats did not. Turmeric extract at 300 mg/kg of body mass was effective in maintaining cellular homeostasis of aortic tissues under hypercholesterolemia, likely by preventing atherosclerosis and senescence in aortic tissues.
The mechanistic study showed that the transcripts of bax-α and casp3 were up-regulated but bcl2 gene expression declined in aortic tissues when the animals were fed HCD, strongly suggesting that HCD rats had a higher risk of apoptosis. This risk, however, was greatly reduced by turmeric supplementation in the HCD100Tur and HCD300Tur rats, which up-regulated the transcript level of the bcl2 gene and lowered both bax-α and casp3 levels. This may explain the significant decrease in vasorelaxation observed in the thoracic aortas of the HCD rats, in which apoptosis may have occurred, most likely in the endothelial cells.
In addition to the pro- and anti-apoptotic genes, hsp70 gene expression, which plays a role in the onset and progression of vascular diseases by protecting arteries, was also measured using RT-PCR and found to be increased significantly by turmeric. The expression of hsp70 is rapidly up-regulated in cells under conditions of oxidative stress, which then become resistant to otherwise lethal levels of stress. 39 HSP70 also exerts a protective effect on arteries against atherosclerosis 40,41 and has anti-inflammatory and anti-apoptotic functions. 42 Our study suggests that turmeric could possibly exert vascular protection through the up-regulation of hsp70 gene expression in the aorta.
Our study also showed that the activities of the antioxidant enzymes GSH-PX and SOD were significantly reduced in HCD rats, but not in HCD100Tur and HCD300Tur rats (whose enzyme activities increased to levels comparable to the ND control rats). SOD catalyzes the breakdown of superoxide into oxygen and hydrogen peroxide, providing an important antioxidant function in most cell types when they are exposed to oxidative stress. 43
GSH-PX protects cells against oxidative damage. Decreased GSH-PX is correlated with an increased risk of coronary artery disease 44 and carotid artery disease. 45 The results of our animal study clearly showed that turmeric could increase the activities of both GSH-Px and SOD, thereby rendering greater protection to aortic tissues from oxidative damage.
At doses of 100 and 300 mg/kg of body weight, turmeric extract effected vascular protection and enhanced vasorelaxation in rats. The same doses of turmeric also attenuated and prevented fatty liver in rats in a previous study. 22 These doses were chosen because they are equivalent to 1–3 g of turmeric extract/day when metabolically scaled to humans. 46,47 This quantity is realistic for people to take as most turmeric supplements contain 0.5–1 g of turmeric per capsule.
In conclusion, turmeric improved vasorelaxation and reduced the risk of atherosclerosis in rats. Turmeric increased the bcl2/bax-α ratioin the aortas of rats consuming HCD (Table 4), suggesting that turmeric may prevent apoptosis in aortic tissue. Turmeric also induced higher antioxidant enzyme activities, suggesting that it may protect cells from oxidative damage. A larger-scale study on the beneficial effects of turmeric is being undertaken to establish its impact on health.
Data are mean±SD values (n=6).
The value obtained from the HCD rats was significantly higher than that from the ND rats (control) (P<.05).
The values obtained from the HCDTur100 rats and HCD300Tur rats were significantly lower than that of the HCD rats (P<.05).
Footnotes
Acknowledgments
This study was funded by the Department of Applied Biology and Chemical Technology at the Hong Kong Polytechnic University. The authors would like to thank Mr. Jimmy Tsz for reading the manuscript and Ms. Josephine Keavney for editing this manuscript.
Author Disclosure Statement
No competing financial interests exist.