
Abstract
Agaricus bisporus white button mushroom (WBM) is widely consumed in most countries for its culinary properties. Recently, its dietary intake has been shown to protect against breast cancer. Mushroom polysaccharides are known for their immunomodulating and antitumor properties; however, little is known regarding the properties of A. bisporus polysaccharides. Using size-exclusion chromatography to fractionate the crude extract of A. bisporus, two polysaccharide fractions (designated as ABP-1 and ABP-2) were obtained. The estimated molecular masses of ABP-1 and ABP-2 were 2,000 kDa and 40–70 kDa, respectively, and their sugar compositions consisted mainly of glucose, mannose, xylose, and fructose. Analysis of the effects of the polysaccharides on murine macrophages demonstrated that both fractions stimulated the production of nitric oxide, interleukin-6, and tumor necrosis factor-α. Modulation of macrophage function by A. bisporus polysaccharides was mediated in part through activation of nuclear factor-κB with the production p50/105 heterodimers. Both ABP-1 and ABP-2 had the ability to inhibit the growth of human breast cancer MCF-7 cells but had little effect on the growth of human colon, prostate, gastric cancer, and murine Sarcoma 180 cells as assessed by a tetrazolium dye [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide]–based assay. However, when murine Sarcoma 180 cells exposed to ABP-1 or ABP-2 were implanted subcutaneously into mice, a reduction in tumor growth was observed compared with that observed in control mice. Taken together, our data provide a molecular basis to explain in part the reported beneficial therapeutic effects of A. bisporus WBM intake and suggest that macrophages likely contribute to the antitumor effects of Agaricus polysaccharides.
Introduction
T
Sterols and lectins have been suggested to be among the bioactive substances of A. bisporus WBM, 6,14 but little is known regarding the potential antitumor and immunomodulating properties of polysaccharides. Mushroom polysaccharides are known for their potent immunostimulatory properties, which can be demonstrated not only through direct antitumor activity against cancer cells, 15 –17 but also through activation of cells involved in innate immunity. 18 –20 In the latter, activation of macrophages leads to secretion of pro-inflammatory cytokines—cytotoxic and inflammatory molecules, such as interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and nitric oxide (NO), that promote destruction of tumor cells. 21 –23 Based on these and other studies showing that many mushroom polysaccharides exert macrophage immunomodulatory and antitumor activities, 3,16,18 we suggest that the putative protective effect of dietary intake of A. bisporus mushroom on breast cancer may be in part attributable to the immunomodulatory and antitumor properties of A. bisporus polysaccharides.
In order to address these questions and to further define the contribution of bioactive compounds to the beneficial nutritional properties of A. bisporus WBM, we extracted and purified polysaccharides from the fruiting bodies of A. bisporus WBM and analyzed their immunomodulatory and antitumor effects on murine macrophage as well as human and murine cancer cells in vitro and in vivo.
Materials and Methods
Reagents
Standard dextrans, Escherichia coli O127:B8 lipopolysaccharide (LPS), and streptomycin were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Dulbecco's modified Eagle's medium, RPMI 1640 medium, fetal bovine serum, and penicillin/streptomycin were purchased from Invitrogen (Carlsbad, CA, USA). Other chemicals used were of reagent grade and obtained commercially.
Animals
Six-week-old male C57BL/6 mice, obtained from Daehan Biolink Co., Ltd. (Seoul, Korea), were housed individually in stainless steel cages with access to food and water ad libitum in a room with controlled temperature (22±2°C), a relative humidity of 55±5%, and a 12-hour cycle of light and dark. All experiments were done in accordance to procedures approved by the University Animal Care and Ethics Committees.
Fractionation and purification of polysaccharides from A. bisporus
The fruiting bodies of A. bisporus WBMs obtained from retail outlets were washed with distilled water, sliced into small pieces, and then homogenized. The homogenate was autoclaved for 2 hours, followed by cooling to room temperature before centrifugation at 3,600 g for 20 minutes as described previously. 24 The supernatant was collected and precipitated with 95% ethanol (1:4 vol/vol) overnight at 4°C. The precipitate containing the polysaccharides was pelleted by centrifugation at 10,000 g for 20 minutes and then resuspended in distilled water. The supernatant was filtered through a membrane filter (pore size, 0.45 μm) and lyophilized to obtain a crude polysaccharide extract with a yield of 2.32 g/kg of fruiting bodies.
The crude polysaccharide extract dissolved in 3 mL of distilled water at a concentration of 10 mg/mL was fractionated by size-exclusion chromatography on a Sepharose CL-6B column (2.4×99 cm) equilibrated with distilled water and eluted with the same solution at a flow rate of 5 mL/12 minutes. The polysaccharide elution profile was determined by the phenol–sulfuric acid method. The relevant fractions were collected, pooled, and concentrated by freeze-drying. Estimation of molecular size of the purified polysaccharide fractions was based on the elution volume and the molecular weight using a standard dextran series of T2000 (20×105), T450 (4.5×105), T150 (1.5×105), T70 (7×104), and T40 (4×104) for calibration of the Sepharose CL-6B column.
Chemical composition analysis
Total protein content of polysaccharide was determined by the method of Lowry et al. 25 using bovine serum albumin as a standard. The total sugar was determined by the phenol–sulfuric acid method 26 using glucose as a reference standard. The sugar composition was analyzed by a Hewlett Packard 5890 gas chromatograph (Agilent Technologies, Palo Alto, CA, USA) equipped with a flame-ionization detector on a BP-20 capillary column (12 m×0.22 mm i.d., 0.25-μm film thickness; SGE Analytical Science Pty Ltd., Ringwood, VIC, Australia) based on the hydrolysis and acetylation method of Jones and Albersheim. 27
Cell culture
Murine J774A.1 macrophages, murine Sarcoma 180 cells, and human cancer cell lines (breast adenocarcinoma [MCF-7], colon adenocarcinoma [HT29], prostate cancer [DU145], and stomach gastric adenocarcinoma [CRL-1739]) were obtained from Cell Bank Australia (Westmead, NSW, Australia). Murine J774A.1 macrophages and murine Sarcoma 180 cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% (vol/vol) heat-inactivated endotoxin-free fetal calf serum, 2 mM glutamine, and antibiotics (100 μg/mL streptomycin and 100 U/mL penicillin). Human cancer cell lines were grown in RPMI 1640 medium supplemented with 10% fetal calf serum and antibiotics (100 μg/mL streptomycin and 100 U/mL penicillin). All cell lines were cultured at 37°C in a humidified atmosphere containing 5% CO2. Cell numbers and viability were assessed microscopically by trypan blue exclusion before use.
NO production
J774A.1 macrophages were incubated at 37°C in an atmosphere of 5% CO2 for 24 hours in culture medium containing various doses of polysaccharide fraction or LPS as a positive control in wells of 96-well flat-bottom microtiter plates at a density of 1×105 cells per well. After incubation, 100 μL of the culture supernatant was removed and assayed for NO production using a colorimetric method with sodium nitrite as a standard. In brief, supernatants were mixed with equal volumes of Griess reagent 21 (0.1% [wt/vol] naphthyl ethylenediamine and 1% [wt/vol] sulfanilamide in 5% [vol/vol] phosphoric acid) in wells of 96-well flat-bottom microtiter plates. After 5 minutes at room temperature, the absorbance was measured at 550 nm in a Bio-Rad (Hercules, CA, USA) microplate reader.
Assay of IL-6 and TNF-α production
J774A.1 cells were incubated for 24 hours in culture medium with or without various doses of polysaccharide fraction or LPS as a positive control in 96-well flat-bottom microtiter plates at a density of 2×105 cells per well. IL-6 and TNF-α secreted in the culture supernatants were quantifieded using enzyme-linked immunosorbent assay kits (BD Biosciences, San Jose, CA, USA). Cytokine concentrations were determined by extrapolation from TNF-α and IL-6 standard curves, according to the manufacturer's protocol.
Assay of nuclear factor-κB activation
J774A.1 cells were incubated for 24 hours with or without polysaccharide fraction or LPS as a positive control. Analysis of p50 binding to its consensus oligonucleotide was performed in the cell lysates using the enzyme-linked immunosorbent assay–based nuclear factor-κB (NF-κB) family EZ-TFA transcription factor assay kit (Millipore, Temecula, CA, USA). In brief, nuclear extract proteins (5 μg) from control or polysaccharide-treated cells were incubated with an oligonucleotide containing the NF-κB consensus binding site (5′-GGG ACT TTC C-3′) bound to wells of a 96-well flat-bottom microplate. After extensive washes, the NF-κB complexes bound to the oligonucleotides were incubated with rabbit anti-NF-κB family gene antibody (1:250–1:1,000 dilution), followed by further extensive washing. The plate was then incubated with a secondary antibody (goat anti-rabbit, horseradish peroxidase–conjugated, 1:500 dilution), and tetramethylbenzidine (substrate) was added for color development, which was read at 450 nm using a reference wavelength of 650 nm.
Antitumor activity of the polysaccharide in culture
The in vitro effects of the polysaccharides on the growth of murine Sarcoma 180 cells and several human tumor cell lines were determined by the ability of living cells to reduce the yellow 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) dye to a purple formazan product. Cells were plated at a density of 1×105 cells per well in wells of 96-well flat-bottom microtiter plates. After 24 hours, the polysaccharide fraction in phosphate-buffered saline was added to each well to a final concentration ranging from 10 to 100 μg/mL followed by incubation for 24–72 hours. Phosphate-buffered saline was used as a control. After incubation, the plates were centrifuged, and the media were removed and replaced by adding 200 μL of fresh medium containing MTT (100 μg/200 μL) to each well. After a further incubation for 2 hours, the cells were lysed with dimethyl sulfoxide (200 μL per well), and the reduced intracellular formazan product was quantified in a Bio-Rad enzyme-linked immunosorbent assay microplate reader at 550 nm. Cell viability was expressed as the percentage of control absorbance at 550 nm.
Effect of the polysaccharide on tumor growth in vivo
Murine Sarcoma 180 cells (6×106 cells/200 μL) following centrifugation were resuspended in phosphate-buffered saline containing 200 μg of the polysaccharide fraction before implantation by subcutaneous injection into the left hind groin of the mice. Control mice were implanted with the tumor cells alone. Each group consisted of 10 mice. On day 28, the tumors were removed and weighed. The antitumor effect was measured by calculating the inhibition ratio (%) using the expression ([A – B]/A)×100, where A and B are the average tumor weight of the control and treated groups, respectively. 16
Statistical analysis
Data are expressed as mean±SE values. The group mean was compared using a one-way analysis of variance and Duncan's multiple range tests. The statistical difference was considered significant at P<.05. The analysis was performed using SPSS Version 18 Software (IBM SPSS, Chicago, IL, USA).
Results and Discussion
Fractionation and purification of A. bisporus polysaccharides
Polysaccharides from A. bisporus WBM were fractionated by Sepharose CL-6B size-exclusion chromatography to obtain two main fractions selected on the basis of the total carbohydrate elution profile (designated as ABP-1 and ABP-2) (Fig. 1A). ABP-1 was represented by a single symmetrical peak on the size-exclusion chromatogram, suggesting that the polysaccharide fraction was relatively homogeneous compared with the ABP-2 fraction, which was represented by a broad peak. Each of the two fractions contained at least 81% carbohydrate and 19% protein (Table 1). Based on the calibration curve obtained from analysis of dextran molecular weight standards, the molecular mass of the ABP-1 fraction was relatively large, with an estimated molecular mass of 2,000 kDa compared with that of the ABP-2 fraction, which contained polysaccharides with molecular masses ranging from 40 to 70 kDa (Fig. 1B). The molecular mass of the ABP-1 polysaccharide fraction was outside the range of those (800–1,200 kDa) reported for other mushroom fruiting body polysaccharides. 28,29 Our method of hot water extraction involved homogenization followed by autoclaving for 2 hours, which may produce polysaccharides with a wider range of molecular masses, so this could explain the differences between laboratories. In the present study, despite the generally single symmetrical peak on the chromatogram, it is possible that the ABP-1 fraction may contain large protein-bound polysaccharides of molecular masses within a void exclusion range that cannot be resolved into distinct peaks by gel filtration chromatography. Clearly, further studies are necessary to determine the molecular characteristics of these polysaccharides. In the current study, sugar composition analysis revealed that the A. bisporus polysaccharide fractions consisted primarily of glucose, mannose, xylose, and fructose (Table 1). ABP-1 fraction consisted mainly of mannose and xylose; the major sugar constituents of ABP-2 fraction were glucose and fructose. Both fractions contained small or trace amounts of ribose, rhamnose, and fucose.

Gel filtration chromatogram and molecular weights of A. bisporus polysaccharides (ABPs).
Effect of A. bisporus polysaccharides on macrophage IL-6, TNF-α, and NO production and NF-κB activation
Macrophages play a pivotal role in host defense, 30 including production of cytokines, antigen processing and presentation to T cells, phagocytosis of pathogens, and autophagy. A variety of mushroom polysaccharides can modulate macrophage function as determined by the production of pro-inflammatory molecules such as cytokines and NO, 28,31,32 which play a critical role in host immune response and tumorigenesis. In the present study, treatment of J774A.1 macrophages with A. bisporus polysaccharide fractions resulted in a dose-dependent increase in the production of IL-6, TNF-α, and NO (Fig. 2), with ABP-1 being significantly more active than ABP-2, especially at the low concentrations of 50 and 100 μg/mL for IL-6 and TNF-α (P<.01). Both ABP-1 and ABP-2 stimulated IL-6 and TNF-α production significantly more than LPS (P<.01). The difference in the immunostimulatory capacities of ABP-1 and ABP-2 fractions may be due to ABP-1 being more complex, being of higher molecular weight, and, in addition to glucose (13.5%), also containing higher proportions of xylose (25.5%), fructose (12.8%), and mannose (39.2%) (Table 1). IL-6 and TNF-α are important pro-inflammatory cytokines that can act on immune and inflammatory cells in a paracrine/autocrine manner in host defense, apoptosis, immune regulation, and homeostasis. 33,34 In addition to enhancing cytokine production, A. bisporus polysaccharides also activated macrophage NO production (Fig. 2C). Both ABP-1 and ABP-2 fractions were equally effective in inducing NO production, the levels of which were comparable to that induced by LPS at 1 μg/mL. However, ABP-2 was more effective than ABP-1 at 200 μg/mL compared with LPS alone. NO production by ABP-1- and ABP-2-stimulated J774A.1 macrophages treated with polymyxin B endotoxin–removing gel was essentially the same, whereas LPS treatment combined with polymyxin B significantly diminished NO production (data not shown), suggesting that LPS was not responsible for the observed effects of ABP-1 and ABP-2 polysaccharides. Taken together, the results suggest that NO production by J774A.1 macrophages stimulated with ABP-1 and ABP-2 may be dependent on the composition of the monosaccharides that form the characteristic highly repetitive structure of polysaccharides. However, macrophage immunostimulatory activity of ABP-1 and ABP-2 seems to be positively correlated with molecular weight of the polysaccharides, with the higher-molecular-weight ABP-1 fraction being the most active in regard to IL-6 and TNF-α production.
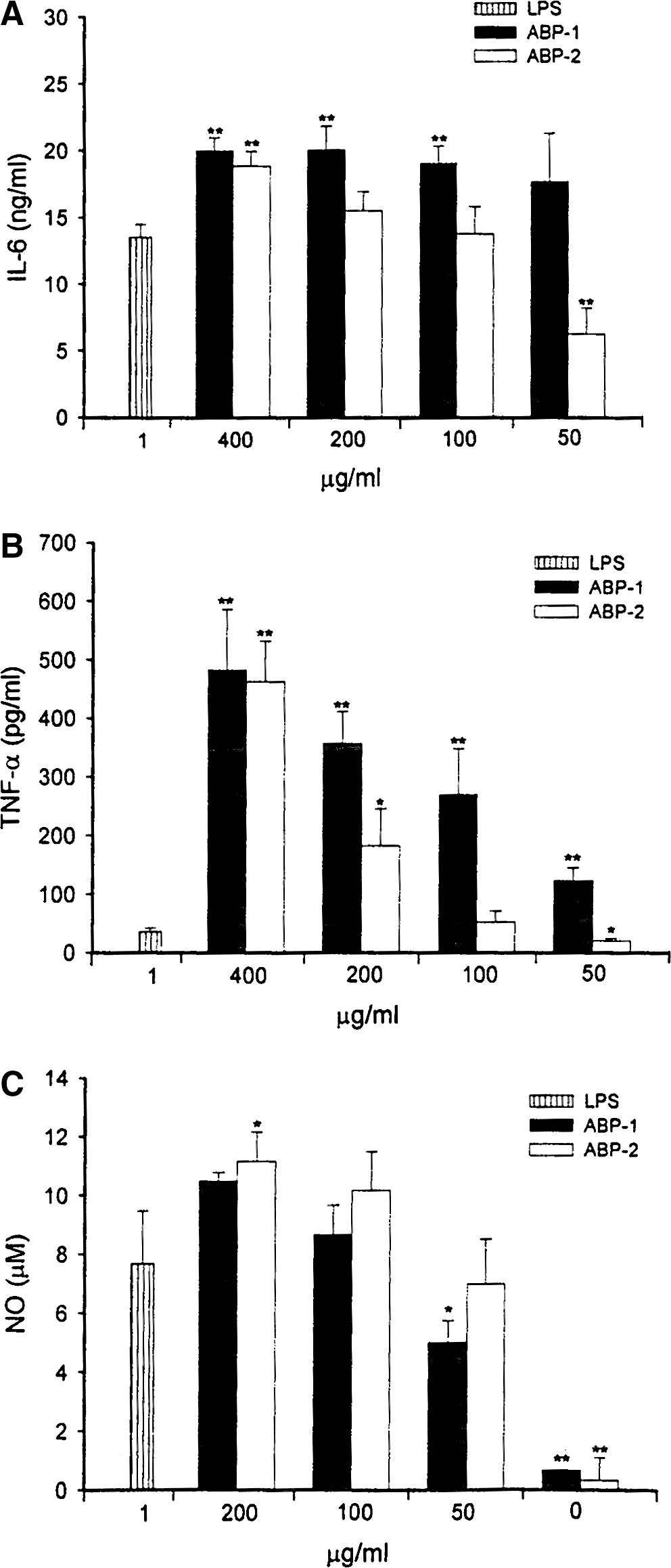
Effects of A. bisporus polysaccharides on
Activation of macrophages is known to lead to protein phosphorylation by kinases as well as the activation of nuclear transcription factors such as NF-κB, which includes the families 34 Rel (c-Rel), RelA (p65), RelB, NF-κB1 (p50/p105), and NF-κB2 (p52/p100) heterodimers. Our results demonstrated that treatment of J774A.1 macrophages with ABP-1 or ABP-2 at 100 μg/mL led most commonly to the activation of heterodimer RelA/NF-κB1 (p50) (Fig. 3). ABP-1 was more active than ABP-2 (P<.05) in stimulating NF-κB activity, but both were more active than LPS alone (P<.01) (Fig. 3). In support of our findings, several mushroom polysaccharides have been previously reported to activate NF-κB, leading to increased immune cytokine mRNA and inducible NO synthase gene expression. 17,35 However, whether ABP-1 and ABP-2 polysaccharides enhance IL-6, TNF-α, and NO production through NF-κB, or vice versa, requires further investigation.
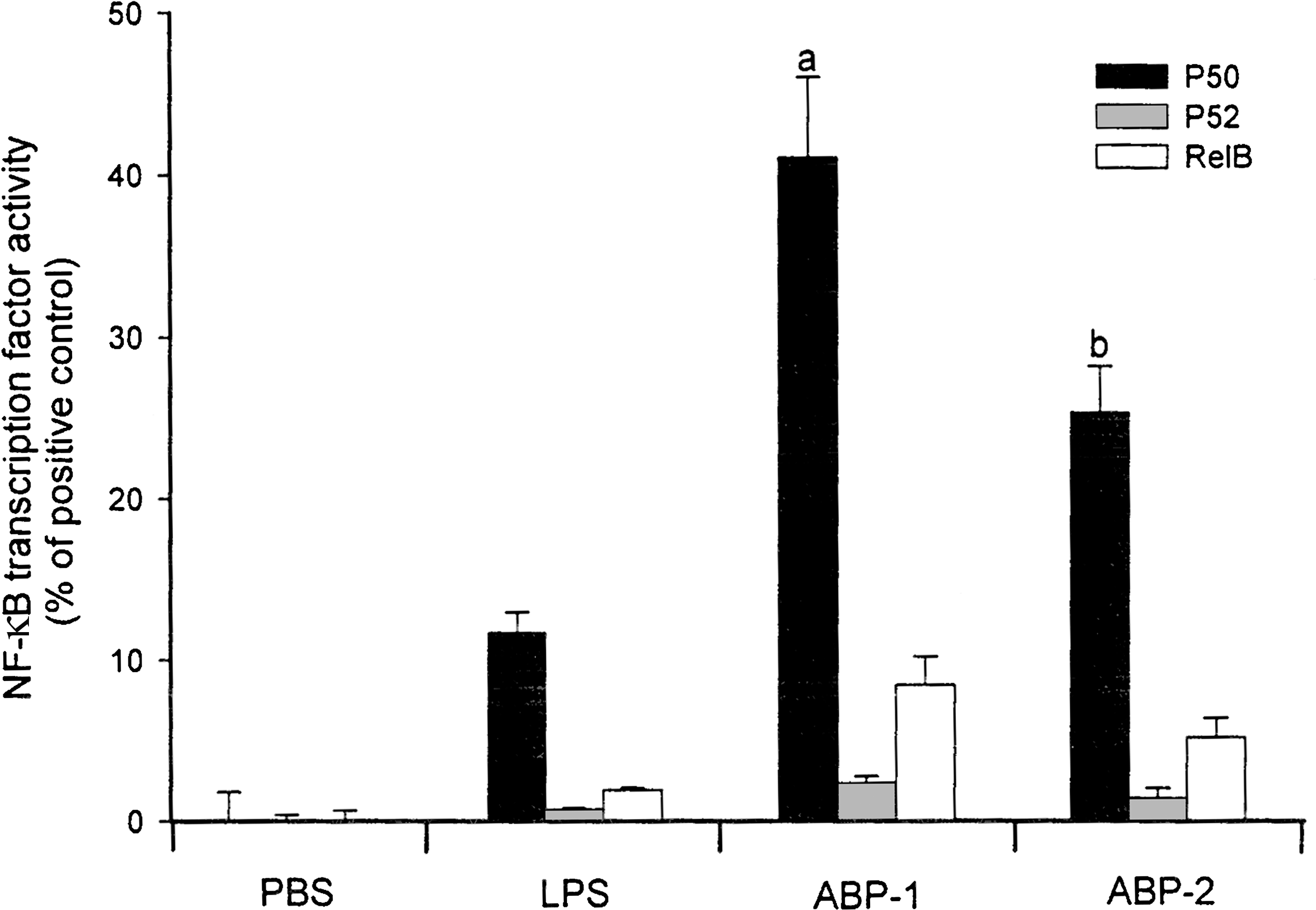
Effects of A. bisporus polysaccharides on nuclear factor-κB (NF-κB) activation. Murine J774A.1 macrophages were incubated for 24 hours with the indicated concentrations of A. bisporus ABP-1 and ABP-2 polysaccharide fractions, and p50, p52, and RelB heterodimers bound to oligonucleotide containing the NF-κB consensus binding site 5′-GGG ACT TTC C-3′ were quantified by enzyme-linked immunosorbent assay. E. coli LPS was used a positive control. All results were expressed as percentages of NF-κB family transcription factor activity of a TNF-α-treated HeLa cell extract. Data are mean±SD values of triplicate samples from one representative experiment. a P<.01 versus LPS alone, b P<.05 versus ABP-1. PBS, phosphate-buffered saline.
Effect of A. bisporus polysaccharides on growth of tumor cells in vitro and in vivo
The effect of increasing concentrations of ABP-1 and ABP-2 polysaccharide fractions on human tumor cells and murine Sarcoma 180 cells using the MTT assay was examined during their active growth phase set at 24 hours. Treatment of tumor cell lines with ABP-1 and ABP-2 at various doses (both 0–100 μg/mL) dose-dependently inhibited the growth of breast cancer cells (MCF-7) but had no or minimum effect on other tumor cells (Fig. 4), including murine Sarcoma 180 cells (Fig. 4E). In all cases, ABP-1 was found to be more effective than ABP-2. In particular, ABP-1 was more effective than ABP-2 in inhibiting MCF-7 breast cancer cell growth with an estimated 50% inhibition concentration value of 50 μg/mL (Fig. 4A). The dose–response data shown in Figure 4 showed that the polysaccharides exhibited no cytotoxicity against any of the tumor cells tested (based on a 50% inhibition concentration of >25 μg/mL). When mouse Sarcoma 180 cells were treated with either ABP-1 or ABP-2 fraction (each 100 μg/mL) followed by subcutaneous implantation into the left hind groin of the mice, ABP-1 was found to be more effective in reducing tumor growth (49.8%) (P<.003) than ABP-2 (18.9%) (P<.1) compared with the control group (Fig. 5). Because the polysaccharides do not possess significant in vitro cytotoxic activity against murine Sarcoma 180 cells, the antitumor activity in vivo may be related to their enhancement of macrophage function.
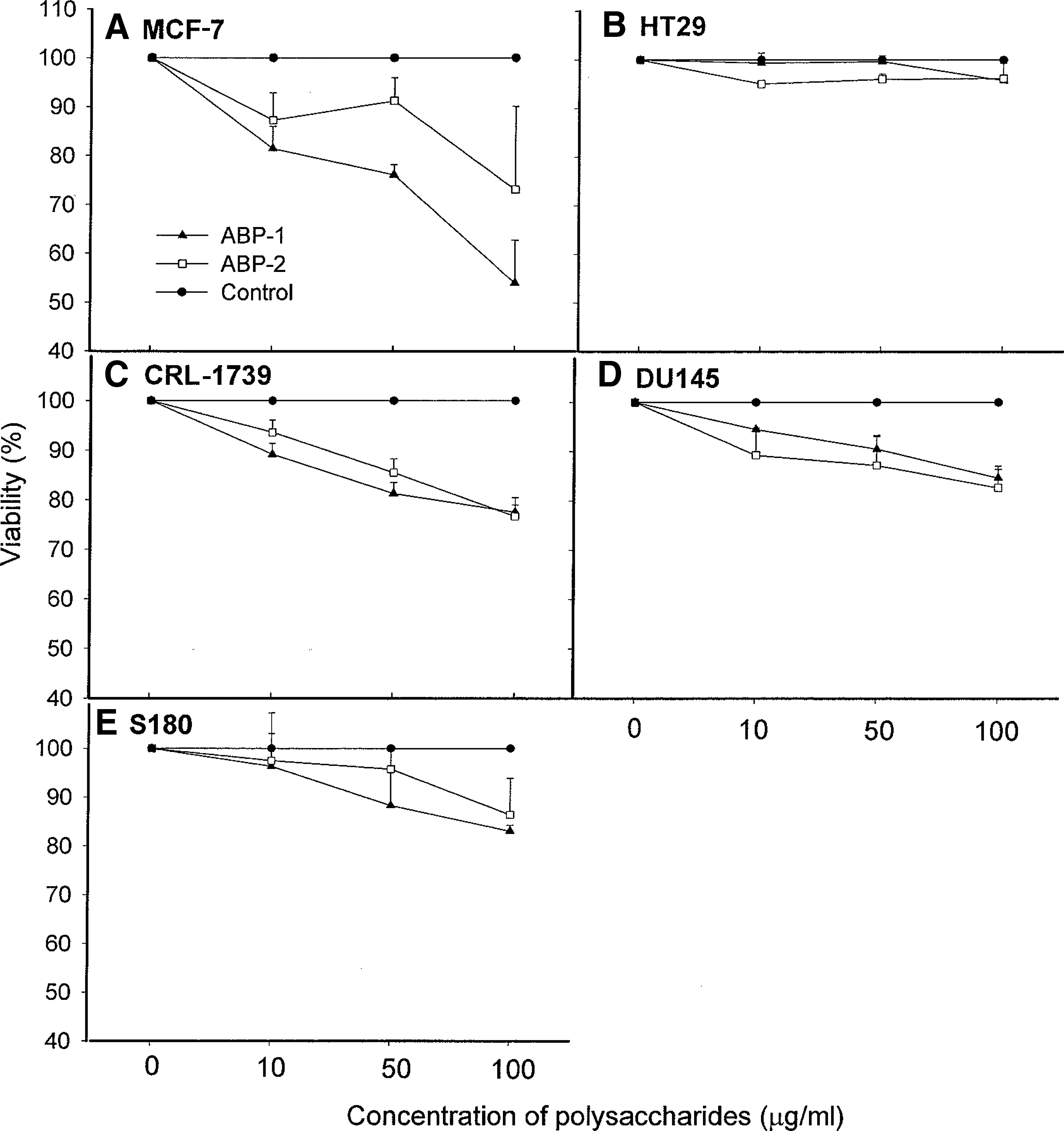
Effects of A. bisporus polysaccharides on the viability of tumor cells in vitro.
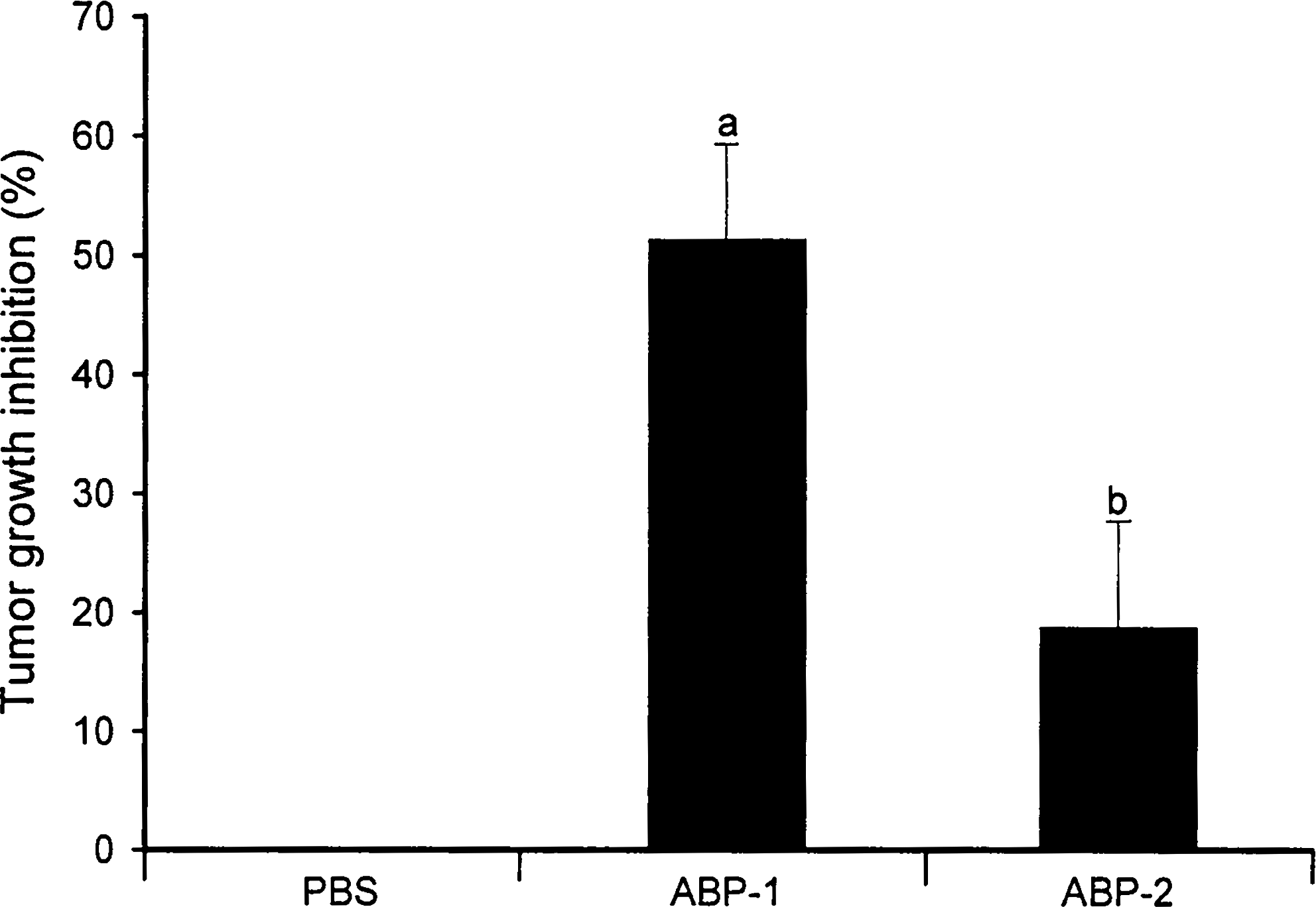
Effect of A. bisporus polysaccharides on tumor growth in S180 tumor-bearing mice. Mice were injected subcutaneously in the left hind groin with S180 cells (6×106 cells per animal) in PBS containing 200 μg of ABP-1 or ABP-2 polysaccharide fraction. Control mice received S180 cells with PBS. On day 28, the antitumor effect was determined by weighing the tumor (in g) and by calculating the inhibition ratio (%) using the expression ([A – B]/A)×100, where A and B are the average tumor weights of the control and the treated groups, respectively. Data are mean±SD values from one experiment representative of 10 animals. a P<.003, b P<.083 compared with PBS control.
In summary, previous studies have reported that some of the biological properties of A. bisporus WBM were due to small molecules such as sterols. 6,7 The present study demonstrated that Agaricus polysaccharides have significant immunomodulatory and antitumor properties, which may in part contribute to the therapeutic potential of A. bisporus WBM. 5 The difference in the effects of ABP-1 and ABP-2 fractions on cancer cells could be explained by the fact that high-molecular-weight polysaccharides are more active than those of low molecular weights in stimulating macrophages 29,32 as well as inhibiting in vitro and in vivo tumor growth. 36 However, it may also be related to their monosaccharide compositions (Table 1), the primary structure, the type of sugar linkages (α, β, etc.), and the degree of branching. 16,37 It has been reported that A. bisporus polysaccharides with a high glucose content and a branched β(1→3) glucan had minimal effect on human colon cells but had a greater effect on mouse bone marrow macrophages in term of NF-κB activation. 37 Our results are consistent with these observations in that both ABP-1 and ABP-2 polysaccharides stimulated the production of NO, IL-6, and TNF-α and activated NF-κB in J774A.1 macrophages but had no effect on the proliferation of human colon cancer cells or murine sarcoma cells. The observation that ABP-1 and ABP-2 polysaccharides significantly inhibited tumor growth of mouse Sarcoma 180 cells in vivo but not in vitro suggests that enhancement of macrophage function may be necessary for the antitumor activity of A. bisporus polysaccharides. In addition, MCF-7 breast cancer cells were particularly more sensitive than other cancer cell lines to Agaricus polysaccharides, suggesting that the potency of Agaricus polysaccharides may be related to the difference in the interaction between the highly repetitive structures of polysaccharides and the membrane targets on tumor cells. Overall, our data provide a molecular basis to explain the protective effect of dietary intake of A. bisporus WBM reported for breast cancer. 5 Further studies are necessary to determine the precise mechanism(s) of action of Agaricus polysaccharides involved in the expression of the various immunomodulatory and antitumor effects and to elucidate the relationship between structural characteristics and biological activity.
Footnotes
Acknowledgment
This research was supported in part by a grant from the Australian Mushroom Growers' Association and Horticulture Australia.
Author Disclosure Statement
No competing financial interests exist.