
Abstract
In this study, Musca domestica pupae lectin (MPL) was screened for its immunomodulatory effect on macrophages. The phagocytosis of macrophages was improved significantly when they were treated with MPL: remarkable changes were observed in the morphology of the cells, the metabolic abilities of DNA and RNA were enhanced, and the production of hepatin was increased. Meanwhile, compared with the control group, not only the mRNA expressions of tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and interferon-γ (IFN-γ) in macrophages, but also the productions of proteins, were strongly induced by MPL; these effects were inhibited by pyrrolidine dithiocarbamate. Further study suggested that MPL could increase the nuclear factor-κB (NF-κB) p65 level in the nucleus. Overall, these results indicate that the improving immunomodulatory activity induced by MPL is mainly due to the increasing productions of TNF-α, IL-6, and IFN-γ and that the activation of macrophage by MPL is partly mediated via the NF-κB pathway.
Introduction
L
Musca domestica belongs to the subphylum Hexapoda and is rich in many kinds of bioactive substances. As one of the edible insects, 15 M. domestica and its larvae, bred under clean conditions, have been used with a long history as a medicinal food source for humans to improve health in China. 16 –18 On the other hand, M. domestica has been identified as a transport host for a variety of zoonoses. 19,20 The reason why these flies can survive and are rarely infected in promiscuous surroundings from larvae to adults has piqued the interest of researchers for years. Antibacterial and immunoactive substances in M. domestica, such as prophenoloxidase, antibacterial protein/peptide, lysozyme, and other secretions, have been widely studied. However, M. domestica pupae lectin (MPL) has been poorly studied.
In our preliminary studies, the galactose-specific MPLs with molecular sizes of 40 kDa, 84 kDa, and 55 kDa were extracted by affinity chromatography on Sepharose-4B and found to have immunoregulatory function and antitumor activity. 21 –24 Although it is well known that macrophages play a key role in host defense mechanisms, the immunoregulatory function of purified MPL on macrophages remains to be reported. The aim of the present study was to investigate the immunomodulatory effects of MPL extracted from M. domestica pupae on murine peritoneal macrophages in vitro.
Materials and Methods
Materials
RPMI-1640 medium was from Gibco (Grand Island, NY, USA), bovine calf serum was from Hyclone (Logan, UT, USA), and commercially available interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and interferon-γ (IFN-γ) enzyme-linked immunosorbent assay (ELISA) kits were from Jiancheng Bioengineering Institute of Nanjing (Nanjing, China). All other chemical reagents were of analytical grade.
Animals
M. domestica flies were raised by the Tianjin Sanitation and Epidemic Prevention Station, Tianjin, China.
Female Kunming mice between 6 and 8 weeks old (weighing 18.5±1.0 g) were purchased from the Research Center of Tianjin Animals Institute, Tianjin. Food and water were provided ad libitum. Mouse holding rooms were kept at 21–25°C and 50% relative humidity with a 12-hour/12-hour light/dark cycle.
The MPL was purified as previously described by our laboratory. 24
Isolation of peritoneal macrophages
Peritoneal fluid from female Kunming mice was harvested from peritoneal cavities by infusing 5 mL of ice-cold sterile phosphate-buffered saline (PBS) (pH 7.2). After centrifugation at 120 g for 5 minutes, the cell pellets were suspended in RPMI-1640 medium supplemented with 10% (vol/vol) bovine calf serum, 100 U/mL penicillin, and 100 U/mL streptomycin and seeded in a cell culture flask at a cell density of 5×105 cells/mL. After incubation for 3 hours at 37°C in a 5% CO2 humidified incubator, nonadherent cells were removed by washing twice with PBS, and freshly prepared medium was added. 25 The viability of the adherent cells was assessed by trypan blue exclusion test. The proportion of macrophages was determined by cell morphology.
Pinocytic activity assay
The cells (1×106 cells/mL) were cultured in 96-well microtiter plates in a final volume of 100 μL. Pyrrolidine dithiocarbamate (PDTC) (Sigma, St. Louis, MO, USA), a known chemical inhibitor of nuclear factor-κB (NF-κB), was added for 1 hour to provide additional evidence on the role of MPL in the activation of NF-κB. Afterward, MPL in various concentrations (5, 10, 20, 40, and 80 μg/mL) was added to both normal cells and PDTC-pretreated cells. After incubation at 37°C in 5% CO2 for 48 hours, the cell culture medium was removed, and 100 μL per well of 0.1% neutral red was added and incubated for 4 hours. The medium was then discarded, and macrophages were washed twice with PBS (pH 7.4). Washed cells were resuspended in 100 μL per well of cell lysing solution and cultured for 2 hours before the absorbance at 540 nm was measured using an ELISA reader (Thermo, Waltham, MA, USA).
Observation of morphological changes
The culture coverslips were placed on six-well plates. The prepared macrophages (100 μL per well) were added and allowed to incubate at 37°C in a 5% CO2 incubator for 4 hours. The cells were then treated with 20 μg/ml MPL for 48 hours or with PBS as the blank control. For the PDTC inhibition group, 20 μg/mL MPL was added to cells that were treated with 100 μM PDTC for 1 hour. Then the culture coverslips were taken out with macrophages adherent to them. The cells were fixed on the slide with 70% ethanol and washed three times with PBS for the observations by scanning electron microscopy, acridine orange staining, and periodic acid–Schiff staining.
Observation with the scanning electron microscope
Cells were prepared for scanning electron microscopy by mixing an equal volume of fixative solution containing 4% (vol/vol) glutaraldehyde in 0.2 M PBS (pH 7.2) at 25°C. The cells were rinsed with 0.1 M PBS and fixed in 1% osmium tetroxide for 30 minutes. The osmium-fixed cells were then rinsed with 0.1 M PBS and dehydrated with a graded ethanol series from 30% to absolute ethanol before air-drying at room temperature. The dried cells were then coated with gold and observed with a XL 30 E scanning electron microscope (Philips, Eindhoven, The Netherlands).
Acridine orange staining
Acridine orange (8.5 μg/mL) was added to each slide and incubated at 25°C away from light for 20 minutes. Fluorescence was detected by a fluorescence microplate reader using the following filter sets: excitation at 485 nm, emission at 530–640 nm. 26
Periodic acid–Schiff staining
For the detection of hepatin, the culture coverslips with adherent macrophages were washed three times with PBS (pH 7.2). After fixation in 75% ethanol for 10 minutes, the cells were then immersed in 1% periodic acid solution for 5 minutes and subsequently treated with Schiff's reagent for 15 minutes. All procedures were performed at room temperature. The stained cells were observed under the inverted microscope.
RNA preparation and reverse transcription–polymerase chain reaction
As described above, cultured peritoneal macrophages that were pretreated with or without 100 μM PDTC were stimulated with 10, 20, or 40 μg/mL MPL for 12 hours. Total RNA was obtained from 1×106 cells using TransZol reagent (TransGen Biotech, Beijing, China) following the manufacturer's instructions. The reverse transcription of 5 μg of RNA was performed using EasyScript First-Strand cDNA Synthesis SuperMix (TransGen Biotech). The polymerase chain reaction was carried out with the primers of 5′-CAGAAAACAATCTGAAACTTCCA-3′ (forward primer) and 5′-AGTTCTTCGTAGAGAACAACATA-3′ (reverse primer) for IL-6, 5′-TGAAGGGAATGGGTGTT-3′ (forward primer) and 5′-CAGGGAAGAATCTGGAAAGGT-3′ (reverse primer) for TNF-α, and 5′-TTCTGCCAGCACTATTTG-3′ (forward primer) and 5′-AGACATCTCCTCCCATCA-3′ (reverse primer) for IFN-γ.
ELISA for IL-6, TNF-α, and IFN-γ
Macrophages (1×106 cells/mL) were cultured in the presence of various concentrations of MPL in a 96-well plate in a total volume of 100 μL for up to 48 hours. Productions of IL-6, TNF-α, and IFN-γ were measured using ELISA kits.
Western blot analysis for NF-κB p65
After treatment with MPL for 48 hours, nuclear proteins were prepared using the Nuclear and Cytoplasmic Protein Extraction Kit (Beyotime, Jiangsu, China). Equal amount (30 μg) of protein were then subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (12% separating gel and 5% stacking gel) and transferred onto a nitrocellulose filter membrane (Bio-Rad, Hercules, CA, USA). After the nonspecific binding site was blocked with 5% skim milk in PBS containing 0.1% Tween-20, a primary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and a secondary antibody–enzyme conjugation were applied to the membrane consecutively. Immunoreactive bands were detected by a diamidobenzidine kit (Zhongshan Goldenbridge, Beijing).
Statistical analysis
Biological assays were performed in triplicate. Results were expressed as mean±SD values. Data were analyzed using analysis of variance (SAS version 9.0, SAS Institute, Cary, NC, USA); P<.05 was considered to be statistically significant by t test.
Results
Effects of MPL on pinocytic activity
Pinocytic activity of macrophages triggered by MPL was examined by the uptake of neutral red (0.1%). Compared with the control group, pinocytic activity of macrophages treated with MPL for 48 hours was enhanced significantly (P<.01) (Fig. 1). The macrophages treated with 20 μg/mL MPL also exhibited the greatest phagocytic activity. The pinocytic activity of macrophages was decreased in the presence of PDTC but was still significantly enhanced in comparison with the control group (P<.01).
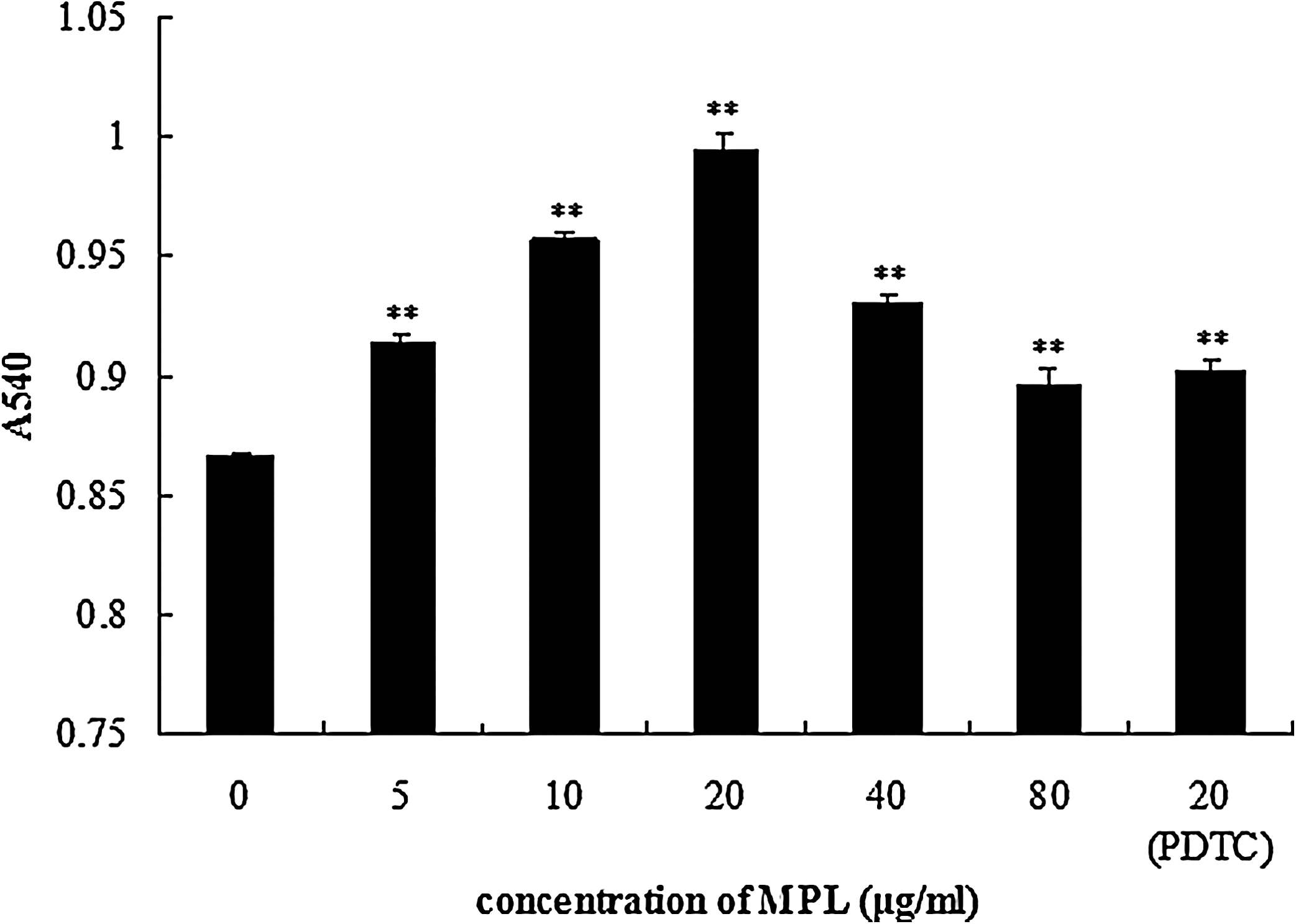
Effects of M. domestica pupae lectin (MPL) on pinocytic activity of macrophages. Macrophages were stimulated by MPL (5–80 μg/mL) for 48 hours. Data are mean±SD values (n=3). **P≤.01 versus control group (cells treated with 0 μg/mL MPL). PDTC, pyrrolidine dithiocarbamate. A540, absorbance at 540 nm.
Effects of MPL on appearance of macrophages
A micrograph of macrophages observed by scanning electron microscopy is shown in Figure 2. The typical morphology of a normal macrophage is shown in Figure 2A, which showed smoother surfaces. However, cells treated with 20 μg/mL MPL visibly increased in size and became irregular in shape compared with normal macrophages; moreover, MPL-treated macrophages became more adherent to the plastic culture dishes with many microvilli-like structures (Fig. 2B). In the PDTC inhibition group (Fig. 2C), although the cells were larger than normal cells, the surface folds and microvilli-like structures were fewer than in MPL-treated cells.
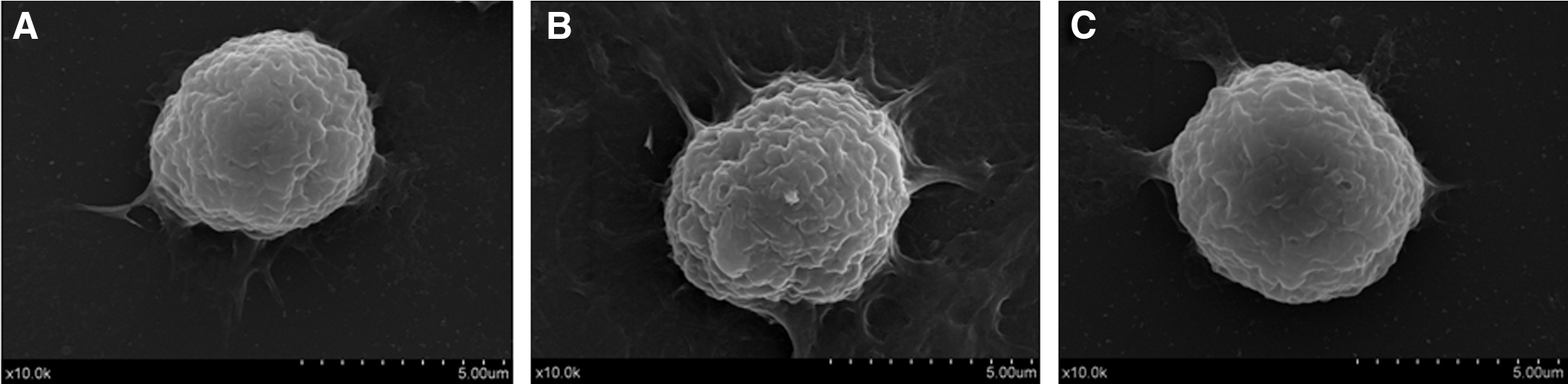
Scanning electron micrographs of macrophages treated with MPL:
Effects of MPL on DNA and RNA of macrophages
The metabolic activity of DNA and RNA in macrophages was studied through acridine orange staining. Red and yellow fluorescences were observed in cytoplasm and nucleus, respectively (Fig. 3). Compared with the control group (Fig. 3A), the intensity of yellow fluorescence in macrophages treated with 20 μg/mL MPL (Fig. 3B) showed obvious enhancement; the intensity of yellow fluorescence in PDTC-pretreated cells was stronger than in the control group, but a little weaker than in the cells treated with MPL (Fig. 3C).
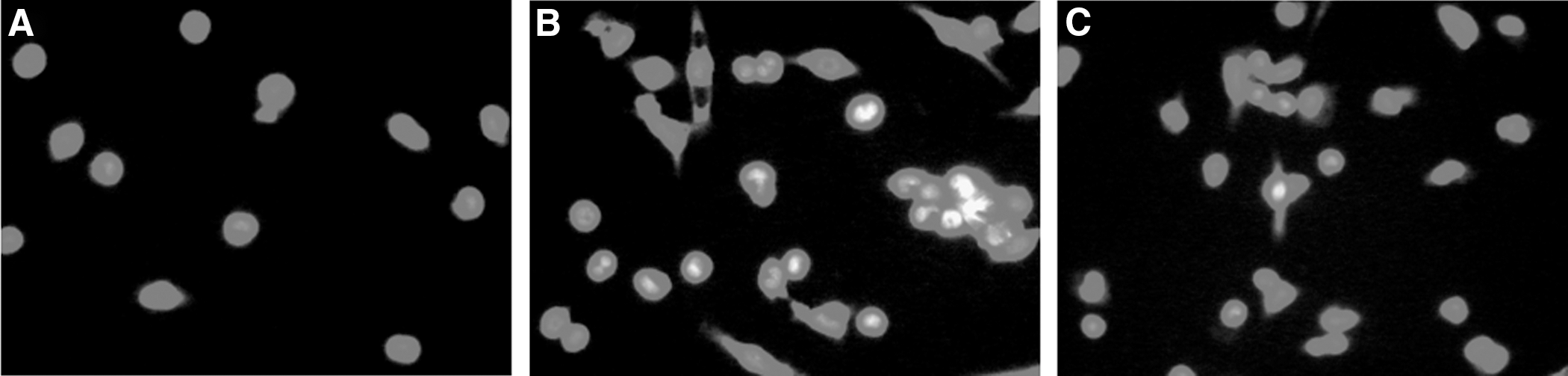
Acridine orange staining:
Effects of MPL on hepatin of macrophages
Periodic acid–Schiff staining was used for detecting the change of hepatin in macrophages. After being stained with Schiff's reagent, the hepatin in the cytoplasm of macrophages was stained to red (Fig. 4). Compared with staining of control cells (Fig. 4A), the degree of red in cytoplasm treated with 20 μg/mL MPL (Fig. 4B) was obviously enhanced, and the color of the cytoplasm of macrophages in the MPL-treated group was darker than that of macrophages pretreated with PDTC.
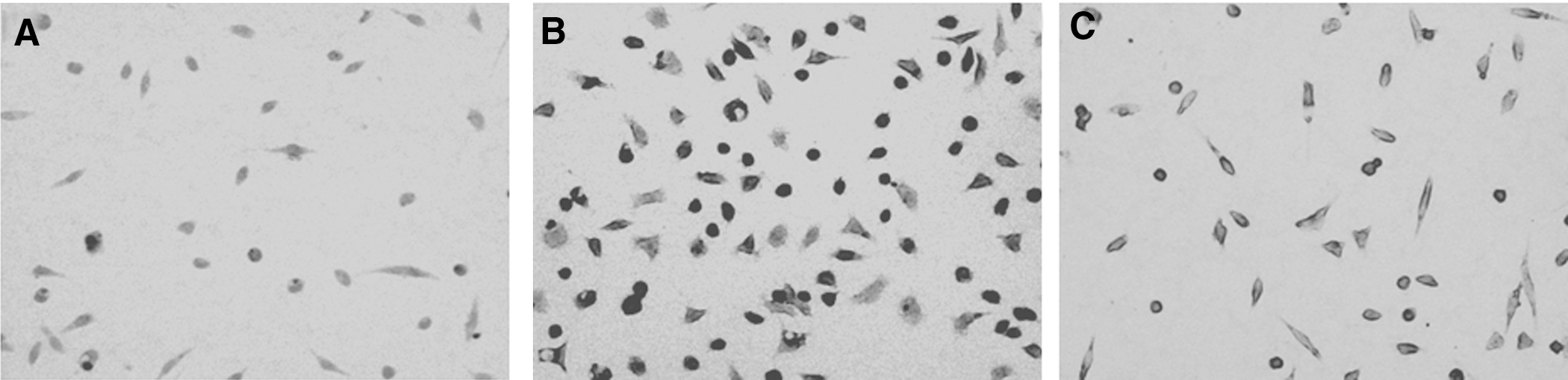
Periodic acid–Schiff staining:
Effect of MPL on mRNA expressions of IL-6, TNF-α, and IFN-γ
The effect of MPL on expression of macrophage cytokines was measured by reverse transcription–polymerase chain reaction (Fig. 5A and B). The mRNA levels of IL-6, TNF-α, and IFN-γ in MPL-treated cells were up-regulated compared with control cells. Moreover, the effect of MPL on mRNA expression was decreased by PDTC pretreatment.
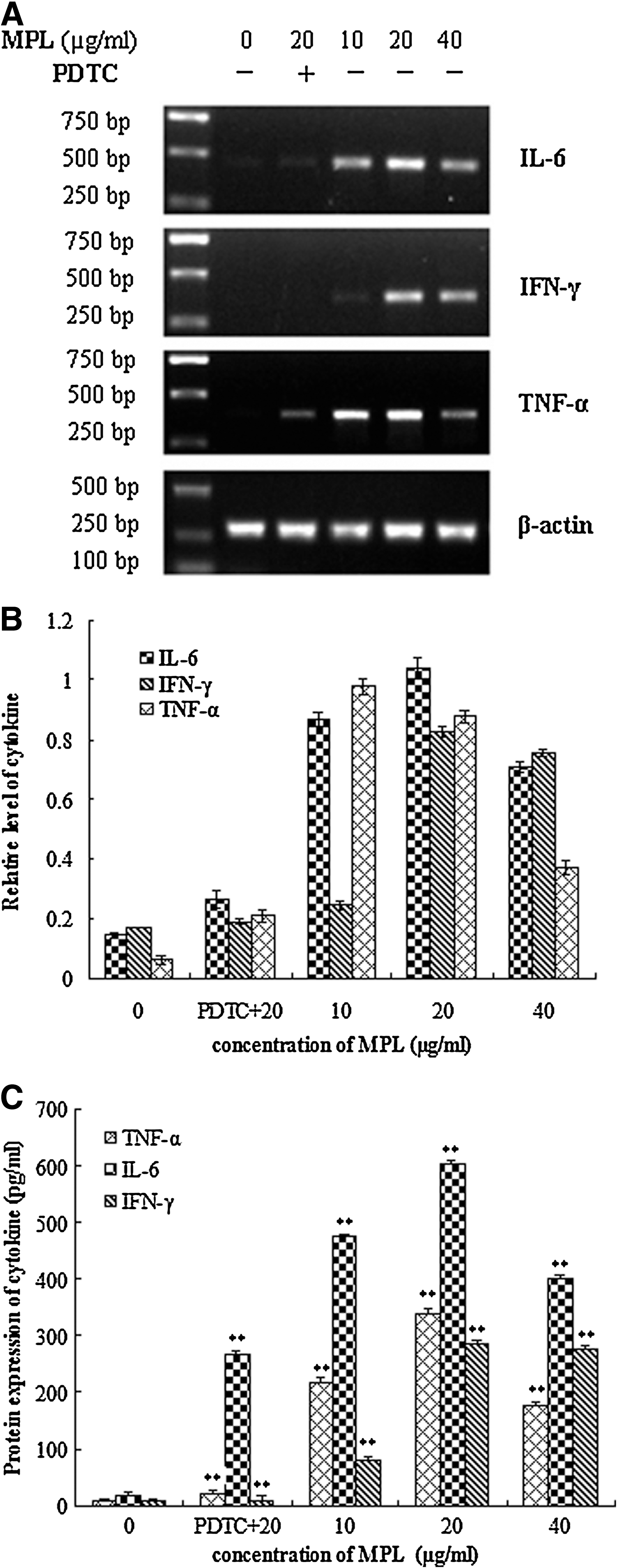
Effect of MPL on protein production of cytokines
TNF-α and IFN-γ concentrations in cell culture supernatants were consistent with mRNA expressions. The productions of IL-6, TNF-α, and IFN-γ in normal macrophages were at low levels, and the secretions of those three cytokines were significantly higher in the MPL-treated group than in normal controls (P<.01). Meanwhile, the up-regulation of cytokines from macrophages by MPL was partly inhibited with PDTC pretreatment (Fig. 5C).
Effects of MPL on NF-κB p65 in macrophages
Western blot analysis was carried out to determine whether NF-κB could be activated in macrophages. The result showed that the protein expression of NF-κB p65 in the nucleus was increased compared with the control group, whereas there was less expression of the protein in the PDTC-pretreated group (Fig. 6).
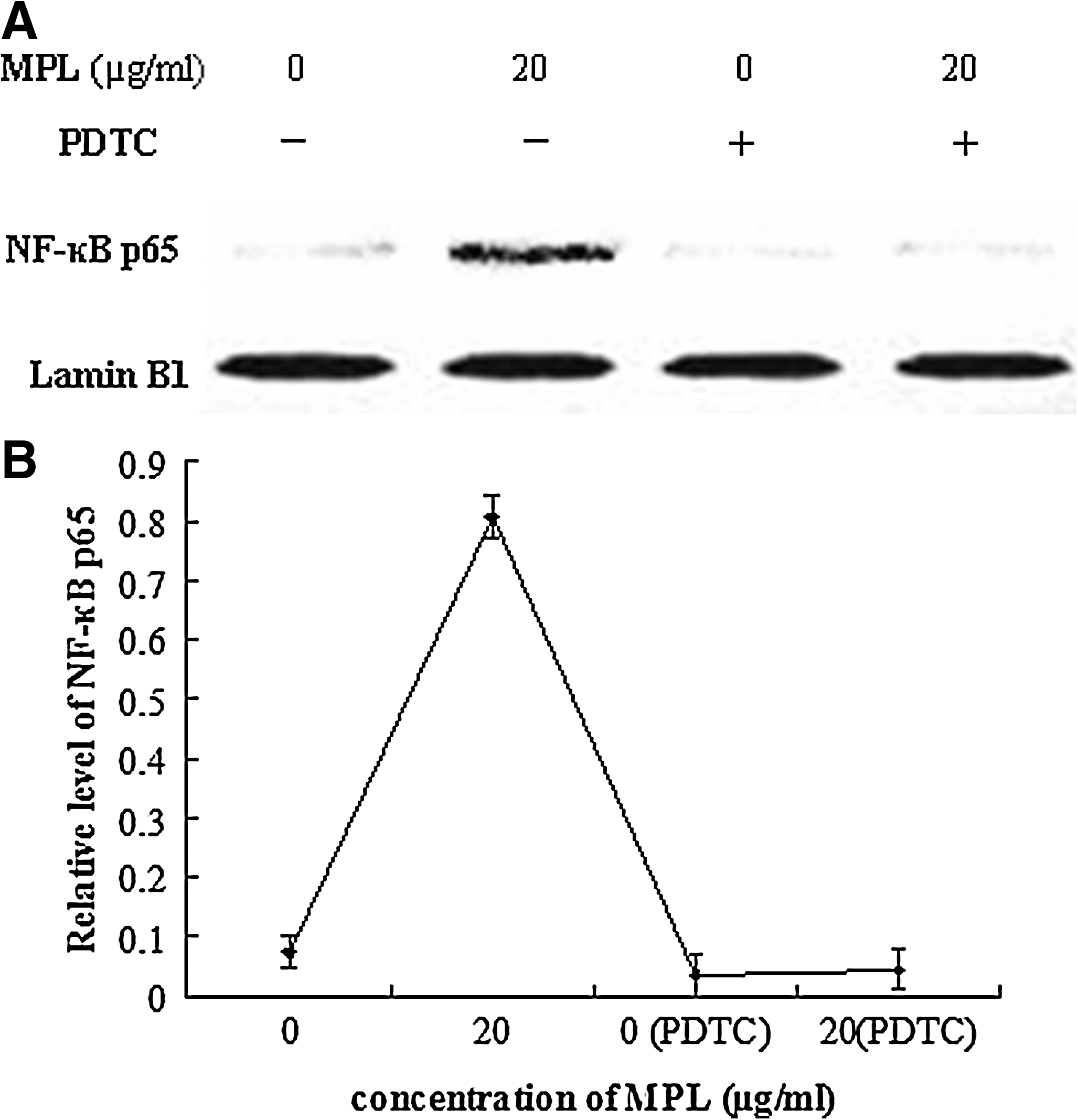
Effect of MPL on nuclear factor-κB (NF-κB) p65.
Discussion
Since Bernheimer first reported that Lepidoptera larvae had hemagglutination activity in 1952, 27 more than 20 kinds of Lepidoptera agglutinins have been purified. Lectins isolated from various Lepidoptera insects have been shown to profoundly affect the immune system in vivo and in vitro. 28 –31 In the present study, immunomodulatory effects of MPL isolated from M. domestica pupae, which inhibited the viability of human breast cancer (MCF-7) cells and human leucocythemia (K562) cells, 24 were investigated in vitro.
It is well known that as phagocytic cells, macrophages are widely distributed throughout the body and play important roles in host defense by coordinating innate immunity and inflammatory responses. 32 The phagocytotic function of phagocytes is an important indicator of the body's immune competence. 33 Several lectins have been reported to have antitumor activities, which were considered to be due to cooperation with macrophages. 34 –36 Furthermore, several studies have proven that lectins improve immunomodulatory activity in macrophages by enhancing the phagocytosis and the secretion of cytokines. 28 –30 Nucleic acid functions in encoding, transmitting, and expressing genetic information and plays a crucial role in biological phenomena such as growth, reproduction, and heredity. 37 As a basic material for synthesis of ATP, hepatin is found in the cytoplasm in many types of cells and forms an energy reserve for release of glucose. In our study, the phagocytosis of macrophages as well as the metabolism of nucleic acid was enhanced remarkably, and the secretion of hepatin in macrophages was increased, when treated with MPL. The appearance of cells also displayed characteristics of activated macrophages, which indicated that macrophages were activated by MPL. IL-6 is considered a major immune and inflammatory mediator. As a pleiotropic cytokine, TNF-α plays an important role in various physiological and pathological processes, including immune and inflammatory responses. 38 It has been reported that TNF-α can promote cell survival through stimulation of NF-κB activity in corneal epithelial cells. 39 As one of the most important endogenous mediators of immunity and inflammation, IFN-γ plays a critical role in normal immune functions, including macrophage activation, inflammation, host defense against intracellular pathogens, etc. 40 Not only the mRNA expressions, but also the protein secretions, of IL-6, TNF-α, and INF-γ in macrophages treated by MPL were up-regulated. Furthermore, the enhancement by MPL of phagocytosis and the up-regulation of IL-6, INF-γ, and TNF-α were partially inhibited by NF-κB inhibitor.
NF-κB was first described as a transcription factor whose function is crucial to immunoglobulin κ gene expression in mature B cells. 41 Constitutively active NF-κB has been found only in the nuclei of mature differentiated macrophages and B cells. 42,43 The NF-κB family plays a crucial role in controlling both innate and adaptive immunity. NF-κB proteins are present in the cytoplasm in association with inhibitory proteins that are known as inhibitors of NF-κB. After being activated, the NF-κB inhibitor proteins become phosphorylated and subsequently degraded by the proteasome. The degradation of NF-κB inhibitor proteins allows nuclear localization of NF-κB. In the nucleus, NF-κB molecules bind with their cognate DNA binding sites to regulate the transcription of a large number of genes, including antimicrobial peptides, cytokines, chemokines, and anti-apoptotic proteins. 41,42,44 Thus, the activation of NF-κB regulates diverse biological processes such as immune and inflammatory responses, cell growth and survival, and tumorigenesis. 45,46 Because of the central role played by these transcription factors in the regulation of the immune system, this pathway has received much attention and become a focus of drug discovery for the treatment of inflammatory diseases and cancer. In this study, we investigated whether the NF-κB was activated by MPL. As a result, the NF-κB p65 protein translocated to the nucleus and participated in the transcription and production of IL-6, INF-γ, and TNF-α. This finding suggested that MPL could up-regulate the expressions of IL-6, INF-γ, and TNF-α via activating the NF-κB p65 signal, among other signaling pathways.
In conclusion, our study documented that proper concentrations of MPL may effectively modulate phagocytosis levels and enhance immune activity of murine peritoneal macrophages in general. We anticipate that MPL will be found to be of great importance to immunity and in hypoimmune diseases via the NF-κB/p65 signaling pathway. Further work will be focused on delineation of the underlying mechanisms and further analysis of signaling pathways.
Footnotes
Acknowledgments
We appreciate the National Natural Science Foundation of China (grants 20676103 and 31000768) and the National High Tech Research and Development Program (863) (grant 2007AA10Z319) for funding.
Author Disclosure Statement
No competing financial interests exist.