
Abstract
Sulfated polysaccharides isolated from Ulva pertusa and fractionated using anion-exchange chromatography were investigated to determine their molecular characteristics and bioactivities. The crude and fractionated polysaccharides (F1, F2, and F3) were mainly composed of carbohydrates (59.9–65.9%), sulfates (11.6–15.3%), and uronic acid (7.30–16.4%) with small amounts of proteins (1.40–4.80%). Rhamnose (62.5–80.7%) was the major monosaccharide unit of these polysaccharides, with different levels of glucose (13.5–27.4%) and xylose (2.74–11.5%). The polysaccharides contained one or two major subfractions with weight-average molecular mass ranging from 51.1×103 to 1,690×103 g/mol. The relatively low in vitro anticancer activity of the polysaccharides (22.3–42.4%) suggested that they had little cytotoxicity against the cancer cell line used (AGS). On the other hand, the polysaccharides significantly stimulated Raw 264.7 cells, inducing considerable amounts of nitric oxide and various cytokines production, which suggested that they could be strong immunostimulators.
Introduction
S
The major monosaccharides of ulvan backbones from the green seaweeds Ulva rigida and Enteromorpha compressa are rhamnose, xylose, and glucuronic acid with repeating disaccharide units of α-
In this study, a water-soluble sulfated polysaccharide from U. pertusa was extracted and subsequently fractionated using ion-exchange chromatography. The purpose of this study was to investigate the molecular and structural characteristics of the fractionated polysaccharides and to evaluate their in vitro antitumor and immunomodulatory activities.
Materials And Methods
Materials
The green seaweed U. pertusa, harvested in the summer of 2009 from the coast of Anin, Gangwon Province, Korea, was purchased from an area wholesaler, extensively washed with distilled water, and air-dried at 60°C. The dried raw material was ground in a blender, sieved (pore size, <0.5 mm), and stored at −20°C before extraction of the polysaccharide. The moisture content of the milled raw material was 10.5%. All the chemicals and reagents used in this work were analytical grade.
Crude polysaccharide extraction
The sample powder (20 g) was treated with 85% ethanol (200 mL) at room temperature overnight with constant mechanical stirring to remove lipophilic substances, such as carotenoid and chlorophyll-related pigments, and low-molecular-weight proteins, followed by centrifugation at 18,500 g for 10 minutes at 10°C. The precipitate was collected, washed with acetone, and dried at room temperature. The dried biomass (20 g) was extracted with distilled water (400 mL) at 65°C with mechanical stirring for 2 hours, followed by centrifugation at 18,500 g for 10 minutes at 10°C. The supernatant was obtained and concentrated to 100 mL by evaporation under reduced pressure at 60°C. Ethanol (99%) (933 mL) was added into the supernatant to obtain an ethanol concentration of 70%, and the solution was kept at 4°C overnight. The crude polysaccharide was obtained by filtering the solution through a nylon membrane (pore size, 0.45 μm) (Whatman International Ltd., Maidstone, United Kingdom) and drying at 30°C in a vacuum oven overnight.
Fractionation of polysaccharide
The crude polysaccharide was fractionated into neutral and acid polysaccharide fractions using ion-exchange chromatography on a DEAE-Sepharose fast flow column (catalog number 17-0709-01, GE Healthcare Bio-Science AB, Uppsala, Sweden) equilibrated with distilled water. The unbound and bound polysaccharides were eluted with a solution of distilled water and increasing concentrations of NaCl (from 0.5 to 1.0 M). The three fractions obtained were hereafter referred to as F1, F2, and F3. The fractions were extensively dialyzed against distilled water for 3 days and lyophilized.
Determination of sulfate, total carbohydrate, protein, and uronic acid contents
Total carbohydrate and protein were determined by the phenol–sulfuric acid method using glucose as a standard 15 and by the Folin phenol reagent (Lowry) method using a commercial assay kit (DC Protein Assay kit, Bio-Rad, Hercules, CA, USA), respectively. The sulfate content of the polysaccharides was determined by the BaCl2-gelatin method using K2SO4 as a standard. 16 Uronic acids were analyzed colorimetrically by m-hydroxydiphenyl analysis according to procedure outlined by Filisetti-Cozzi and Carpita 17 using glucuronic acid as a standard.
Determination of monosaccharide composition
The monosaccharide compositions of the crude polysaccharide and fractions were quantitatively determined using a high-performance liquid chromatography system that consisted of a pump (Waters 510, Waters, Milford, MA, USA), an injection valve (model 7010, Rheodyne, Rohnert Park, CA, USA) with a 20-μL sample loop, a column (carbohydrate analysis column, 4.6×250 mm, Waters), and a refractive index (RI) detector (Waters 2414). The polysaccharide (6 mg) dissolved in 2 M trifluoroacetic acid (0.3 mL) was hydrolyzed at 120°C for 90 minutes. A mixture of acetonitrile and water (80:20, vol/vol) was used as a mobile phase at a flow rate of 2 mL/minute.
Determination of weight-average molecular weight
The weight-average molecular weight (M w) of the polysaccharides was determined by high-performance size exclusion chromatography (SEC) linked to an ultraviolet (UV), multiangle laser light scattering (MALLS), and RI detection system after the polysaccharides were dissolved in distilled water. The high-performance SEC–UV–MALLS–RI system was composed of a Waters 510 pump, Rheodyne model 7072 injector valve with a 200-μL sample loop, a guard column (TSK PW, TosoBiosep, Montgomeryville, PA, USA), an SEC column (TSK G5000 PW, 7.5×600 mm, TosoBiosep), a UV detector at 280 nm (Waters 2487), a MALLS detector (HELEOS, Wyatt Technology Corp., Santa Barbara, CA, USA), and an RI detector (model RI-150, Thermo Electron Corp., Yokohama, Japan). A solution of 0.15 M NaNO3 and 0.02% NaN3 was used as a mobile phase at a flow rate of 0.4 mL/minute. The normalization of the MALLS detector and the determination of volume delay between the MALLS and RI detectors were carried out using bovine serum albumin. The calculations of M w and radius of gyration (R g) were carried out using ASTRA version 5.3 software (Wyatt Technology Corp.).
In vitro anticancer activity assay
The anticancer activity of the polysaccharides was investigated with a human gastric carcinoma cell line (AGS) (American Type Culture Collection, Rockville, MD, USA) by using the 4-[3-(4-iodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolio]-1,3-benzene disulfonate (WST-1) colorimetric assay (Roche Diagnostics, Madison, WI, USA). The cell line (100 μL), with a concentration of 1×105 cells/mL in RPMI-1640 medium containing 10% fetal bovine serum, was plated in a 96-well microplate in triplicate and cultured for 4 hours at 37°C in the presence of 5% CO2. The polysaccharide solution (100 μL), which had various concentrations of 100, 500, and 1,000 μg/mL, or 10 μg/mL 5-fluorouracil (positive control) was added to the cultured cell line in the plate, and the cells were cultured for 48 hours. After cultivation, 120 μL of the WST-1 solution was added to the well, and the solution was further incubated for 4 hours at 37°C. The optical density was measured at 450 nm using a microplate reader (EL-800, BioTek Instruments, Winooski, VT, USA). The anticancer activity, expressed as a percentage of the cancer cell growth inhibition, was calculated according to the following formula: anticancer activity (%)=100×(1 – [A s/A c]), where A s is the absorbance of the sample solution and A c is the absorbance of the control for which 100 μL of H2O was used instead of 100 μL of sample solution. All assays were done in triplicate.
Macrophage proliferation, nitric oxide, and prostaglandin E2 production assay
Raw 264.7 (100 μL, 1×105 cells/mL) (American Type Culture Collection) were incubated in RPMI-1640 medium containing 10% fetal bovine serum with the polysaccharides at concentrations of 1.56, 6.25, and 12.5 μg/mL. All cultures were incubated for 72 hours at 37°C in the presence of 5% CO2. Proliferation was quantified by the WST-1 assay described above. The absorbance (A), converted into the macrophage proliferation ratio (%)=([A Test – A Control]/A Control)×100, at 450 nm was determined with an EL-800 microplate reader.
The immunomodulatory activity of the polysaccharides was determined on the basis of nitric oxide (NO) production in macrophage culture supernatants, and the nitrite concentration was measured by the Griess reaction as described by Green et al. 18 In brief, Raw 264.7 cells (100 μL of 1×106 cells/mL) were plated in 96-well flat-bottomed tissue culture plates and incubated for 24 hours at 37°C in the presence of 5% CO2. The cultured cells were treated with polysaccharide solutions (100 μL) at concentrations of 1.56, 6.25, or 12.5 μg/mL (in triplicates) or lipopolysaccharide (LPS) (1 μg/mL) used as a positive control and incubated for 18 hours at 37°C. The cultured cell supernatant (100 μL) was mixed with an equal volume of Griess reagent (1%, wt/vol) and incubated at room temperature for 10 minutes. The absorbance was measured at 540 nm using an EL-800 microplate reader. The NO production from the Raw cells was calculated with reference to a standard curve obtained with NaNO2 (1–200 μM in culture medium).
Raw 264.7 cells (1×106 cells/mL) were treated with the polysaccharides at various concentrations of 1.56, 6.25, and 12.5 μg/mL or LPS solution (1 μg/mL) used as a positive control in 24-well microplates and incubated for 18 hours. The supernatants were collected for measuring the prostaglandin (PG) E2 concentration (reference compound and assay kit purchased from R&D Systems, Inc., Minneapolis, MN, USA).
Reverse transcription–polymerase chain reaction
Total RNA of Raw 264.7 cells treated with LPS and the polysaccharide solutions was extracted using TRIzol® reagent (Invitrogen, Carlsbad, CA, USA). The concentration of total RNA was measured by a spectrophotometer before constructing cDNA with an oligo(dT)20 primer and Superscript III reverse transcripase (RT) (Invitrogen). The resulting cDNA was amplified by polymerase chain reaction (PCR) using GoTaq® Flexi DNA polymerase (Promega, Madison). RT amplification was conducted with an initial denaturation at 94°C for 3 minutes, followed by 30 cycles of denaturation (94°C for 30 seconds), annealing (56°C for 40 seconds), and extension (72°C for 1 minute) and the final extension step at 72°C for 10 minutes. The product of RT-PCR was separated by gel electrophoresis using 1% agarose gel stained with ethidium bromide, and the gels were viewed under UV transillumination. The forward and reverse primers for inducible NO synthase (iNOS), cycloxygenase (COX)-2, tumor necrosis factor-α, interleukin (IL) 1-β, IL-6, IL-10, and β-actin are shown in Table 1.
COX-2, cyclooxygenase-2; IL, interleukin; iNOS, inducible nitric oxide synthase; TNF-α, tumor necrosis factor-α.
Statistical analysis
A one-way analysis of variance was used to determine the statistical significance of the differences between the various experimental groups. Duncan's post hoc test was used to identify significant differences among the various means. Data are expressed as mean±SD values, and the results are taken from at least three independent experiments performed in triplicate. P values of ≤.05 were considered to be statistically significant.
Results and Discussion
Proximate analysis
The yield and proximate composition of the crude polysaccharide extracted from U. pertusa are shown in Table 2. The yield (14.2%) of the crude polysaccharide was close to the value (18.0%) reported by Chattopadhyay et al., 14 which was quite comparable to the values (8.0–18.3%) reported for other green seaweeds. 14,19,20 The crude polysaccharide consisted of mostly carbohydrates (61.4%) with sulfates (13.9%), uronic acid (13.8%) and proteins (3.4%), in a good agreement with the constituents of the sulfated polysaccharide from Ulva rotundata. 20 The proximate composition of the crude polysaccharide showed a good agreement with the literature, which indicated that the sulfated polysaccharides from other green seaweeds had comparable total carbohydrates (47–53%) with slightly higher amounts of sulfates (17.1–23.6%) and uronic acid (3.2–23.2%) and various protein contents from 1.0% to 13.9%. 9,14,19,21 As shown in Table 2, the major monosaccharide units of the crude polysaccharide were rhamnose (69.8%) and glucose (27.4%) with a minor amount of xylose (2.74%). In the monosaccharide composition of the sulfated polysaccharides from E. compressa, Monostroma latissimum, and G. oxysperma, rhamnose (57.0–78.6%) and glucose (11.5–17.0%) were also major sugars, but the amount of xylose (7.8–15.0%) was varied, indicating that there were considerable differences in the monosaccharide compositions. 4,14,22 Other monosaccharides such as galactose, mannose, and arabinose were not detected in the current study. In general, the proximate compositions of the sulfated polysaccharides from green seaweeds were significantly different depending on species, growing conditions, extraction procedures, and analytical methods. 23,24
Yield=(weight of crude polysaccharide/weight of alga powder) ×100.
Yield=(weight of fractionated polysaccharide/weight of crude polysaccharides injected into ion-exchange chromatography column) ×100.
ND, not detected.
The crude polysaccharide was fractionated on a DEAE-cellulose anion-exchange column and yielded three fractions: F1 (eluted with distilled water) (yield, 21.7%), F2 (eluted with 0.5 mol/L NaCl) (yield, 43.4%), and F3 (eluted with 1.0 mol/L NaCl) (yield, 34.9%). The constituents of three fractions were mainly carbohydrate (59.9–65.9%) with various contents of sulfates (11.6–15.3%) and uronic acid (7.3–16.4%). The sulfate contents slightly decreased from 15.3% (F1) to 11.6% (F3) when the NaCl concentration of the eluent at the ion-exchange chromatography increased to 1.0 M, whereas the opposite trend was observed for uronic acid. This result indicates the various ionic strengths of the three fractions. Rhamnose (62.5–80.7%) was the major sugar of the three fractions, although they also contained considerable amounts of glucose (13.5–25.9%) and xylose (2.8–11.5%). These results suggest that it was possible to obtain the sulfated polysaccharides with various ionic strengths and chemical compositions by fractionating the crude polysaccharide using ion-exchange chromatography.
Molecular characteristics of crude and fractionated polysaccharides
The UV and RI superimposed chromatograms for the crude and fractionated polysaccharides are shown in Figure 1. Tost of the crude polysaccharides were eluted from the SEC column with elution times of 26–49 minutes in two distinct peaks, indicating that they consist of two types of polymers with different molecular weights (Fig. 1A). Of the two peaks, proteins were included in peak II with the elution times of 42–48 minutes as shown in the UV chromatogram. The M w values of the peaks, obtained by the MALLS technique, which provides the absolute molecular weight of polymers, were 1,650×103 and 274×103 g/mol, respectively (Table 3). It was reported that the M w values of the water-soluble polysaccharides from other green seaweeds, such as U. rigida and Capsosiphon fulvescens, ranged from 2,000×103 to 385×103 g/mol, exhibiting significant variations. 5,11 These considerable differences in M w seem to be because of differences not only in the seaweed species, but also in the extraction and analysis methods. In particular, the proper solubilization of the polysaccharides with the additional physical treatment, such as the 30-second microwave heating applied in this study, was considered to be important to obtain the accurate M w of the polysaccharide because the polysaccharide chains could be aggregated by secondary interactions through hydrogen bonding. 24 The R g of the crude polysaccharide was also calculated from the peaks to estimate the approximate size of the polysaccharide (Table 3). The R g values of the peaks were 44.8 and 47.2 nm, respectively.
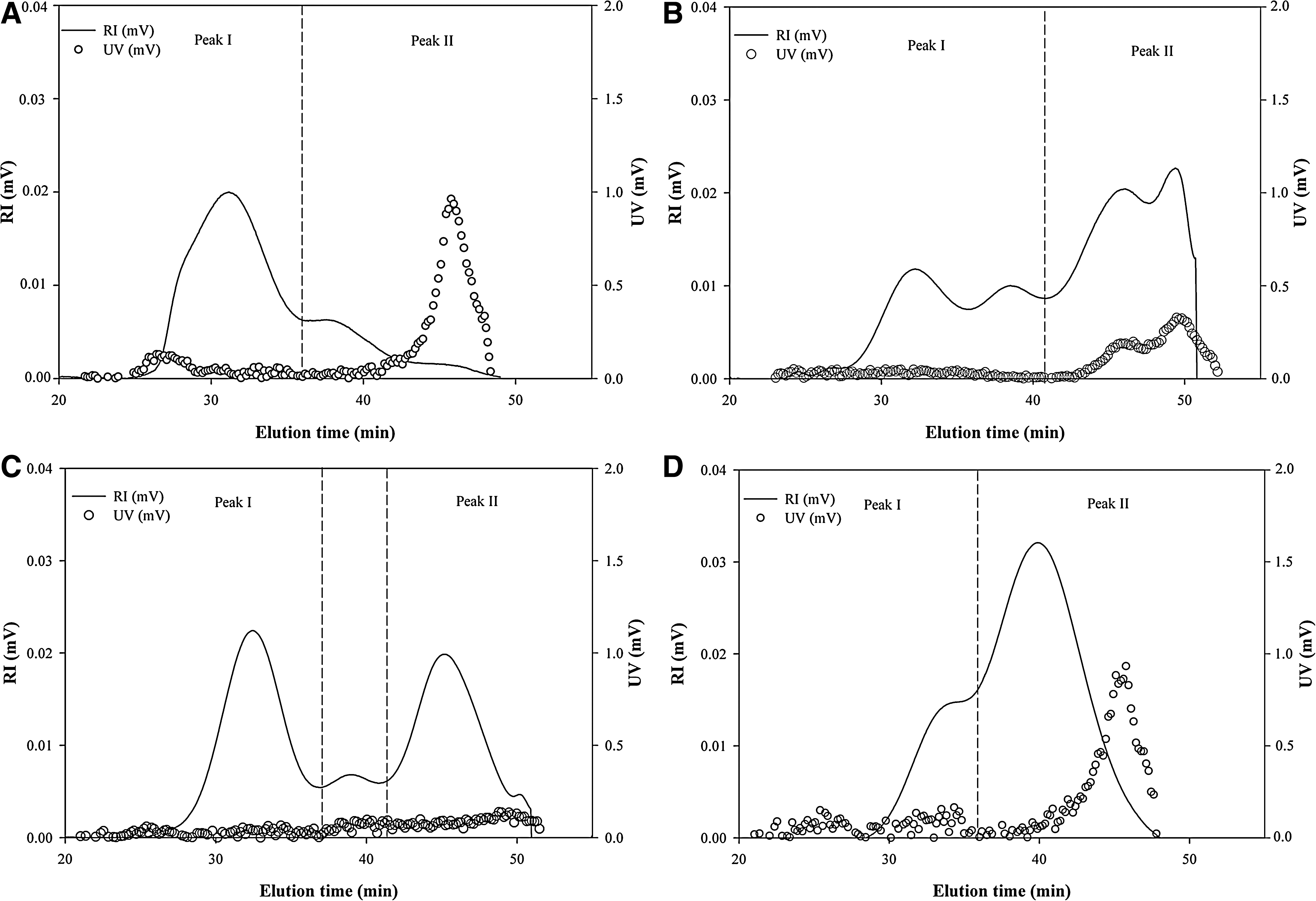
Refractive index (RI) and ultraviolet (UV) chromatograms of
M w, weight-average molecular weigh; R g, radius of gyration; SV g, specific volume for gyration.
As shown in the RI chromatograms, fraction F1 (Fig. 1B) exhibited two major peaks at elution times of 42–51 minutes as well as two small peaks at 29–41 minutes, whereas two distinct peaks between the elution times of 28 to 50 minutes were observed in fraction F2 (Fig. 1C). Fraction F3 was mostly eluted over 40 minutes with a shoulder at 34 minutes (Fig. 1D). These results indicated that the polysaccharides from fractions F1 and F2 were in heterogeneous distribution but those of fraction F3 were relatively homogeneous. The UV chromatograms of the fractions F1 and F3 showed strong UV responses at peak II, indicating the presence of a considerable amount of proteins. On the other hand, no distinct UV response was observed in fraction F2, indicating the absence of detectable proteins. These results suggested that most proteins from the crude polysaccharides were fractionated into fractions F1 and F3 by ion-exchange chromatography, which was in good agreement with the proximate composition of the fractions shown in Table 2. The M
w and Rg
values of the fractions varied dramatically from those of the crude polysaccharide, especially for peak I of F1 and F2, which showed considerably higher Mw
than that of the crude polysaccharide (Table 3). Both values of F3 were also significantly lower than those of the crude polysaccharides. Based on the M
w and R
g values, the specific volume for gyration (SV
g) of the polysaccharides could be calculated as reported by You and Lim
25
by the following equation:
in which the units for SV g, M w, and R g were cm3/g, g/mol, and nm, respectively, and N is Avogadro's number (6.02×1023/mol). SV g values imply the theoretical gyration volume per unit of molar mass and are inversely proportional to the degree of molecular compactness, giving mass-based information on the density of the polysaccharides. The SV g values of the first peaks from the crude and fractionated polysaccharides ranged from 0.1 to 0.2; in contrast, those of the second peaks showed significantly higher values (0.9–2.5) than those of the first peaks (Table 3). These results suggested that the polysaccharides in peak II had less compact and more expanded conformational structures than those of the first peaks. The overall results, therefore, indicated that the fractionation of the crude polysaccharide by ion-exchange chromatography resulted in the production of sulfated polysaccharides with considerable variations in their M w, R g, and molecular compactness.
Anticancer and immunomodulatory activities of the polysaccharides
The anticancer activities of the crude and fractionated polysaccharides were expressed as a percentage of growth inhibition of AGS cells (data are not shown). The crude and fractionated polysaccharides inhibited the growth of the cancer cell line in a dose-dependent manner and showed anticancer activity from about 22% to 42% at the concentrations of 100, 500, and 1,000 μg/mL. The fractionated polysaccharides exhibited slightly greater growth inhibition than the crude polysaccharide; however, the levels of growth inhibition by these polysaccharides were significantly lower than that of the commercial antitumor drug, 5-fluorouracil. This result suggested that these polysaccharides had no considerable direct cytotoxicity for the cancer cell line. Similar levels of anticancer activity were observed by the sulfated polysaccharides extracted from Enteromorpha prolifera and Ulva lactuca, which showed 26% and 22% inhibition of growth on AGS and HT-29 cell lines, respectively. 8,26
It has been reported that, despite little in vitro anticancer activity, the sulfated polysaccharide from the green seaweed Enteromorpha intestinalis exhibited considerable growth inhibition on the cancer cells in in vivo studies. 27 To explain the mechanism of the in vivo anticancer activities, it has been suggested that the polysaccharides might behave as biological response modifiers, enhancing the immune system by the stimulation of splenocytes and macrophages and the induction of NO or cytokine release, which could suppress the growth of cancer cells. 27 Therefore, in the current study, the immunomodulatory effects of the crude and fractionated polysaccharides were observed using Raw 264.7 cells by investigating the amounts of NO and PGE2 released.
The effect of the crude and fractionated polysaccharides on Raw 264.7 cell proliferation was measured at concentrations of 1.56–12.5 μg/mL (data not shown). Raw 264.7 cell proliferation was greatly increased by the presence of fraction F3, whereas fractions F1 and F2 weakly improved the cell proliferation. This result suggested that the polysaccharides were not toxic to Raw 264.7 cells over the concentration ranges tested.
The NO release-inducing capacities of the crude and fractionated polysaccharides at the concentrations of 1.56, 6.25, and 12.5 μg/mL, expressed as the amount of NO released from Raw 264.7 cells, are shown in Figure 2A. LPS (1 μg/mL) was used as a positive control. The level of NO released from Raw 264.7 cells by the crude polysaccharide was found to be considerably higher (19–26 μM) with a dose-dependent response, indicating its strong stimulatory activity on Raw 264.7 cells. Similar amounts of NO were released from Raw 264.7 cells by fraction F1. Further increase of NO release was found by fraction F2 with NO production of 29 μM at a concentration of 12.5 μg/mL, which was comparable to the amount of NO produced by the positive control, LPS. However, little stimulatory activity was observed by fraction F3, with NO production of 10 μM at 12.5 μg/mL. It was not clear why the crude polysaccharides and fractions F1 and F2 exhibited more NO production than fraction F3. According to Leiro et al., 5 a sulfated polysaccharide from U. rigida also significantly stimulated Raw 264.7 cells, resulting in considerable NO release. The authors suggested that the level of NO release was proportionally related to the sulfate content of the polysaccharide. On the other hand, Schepetkin et al. 28 observed a relationship between the Mw and NO production from macrophages, reporting that higher-molecular-weight polysaccharides induced a greater production of NO from Raw 264.7 cells. In the current study, such trends were weakly shown because fraction F3 had the lowest amount of sulfate (Table 2) and its Mw was lower than those of the other polysaccharides (Table 3), which seemed to be responsible for its low NO releasing capacity. However, it was too complex to obtain a strong correlation between the structural and biological activities of the polysaccharides because of their considerable compositional and structural heterogeneity. Further research is necessary to obtain a better understanding of their fine structures and biological activities.
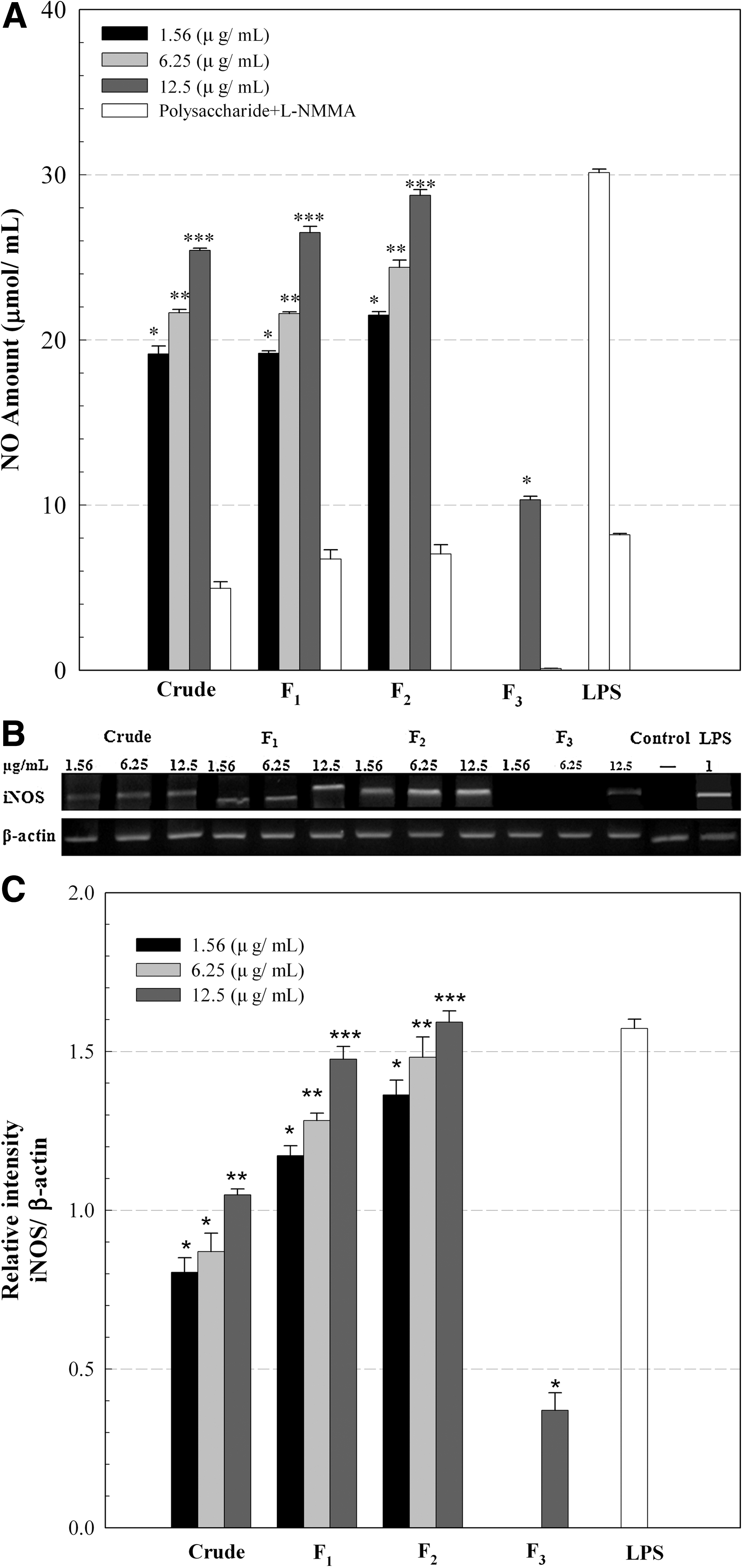
Amount of
Raw 264.7 cells containing the crude and fractionated polysaccharides were incubated in the presence of the iNOS inhibitor N
G-monomethyl-
The crude extract and fractions F1 and F2 at concentrations of 1.56–12.5 μg/mL induced considerable PGE2 production in the range of 2.0–2.6 ng/mL, which was comparable to the level of PGE2 production by the positive control, LPS, and was similar to the trend seen with NO release (Fig. 3A). However, PGE2 production by fraction F3 was relatively small compared with that by the other fractions. As also shown in Figure 3A, the production of PGE2 was considerably inhibited by the presence of indomethacin, a PG synthetase (COX) inhibitor, suggesting that the crude and fractionated polysaccharides might enhance the expression of COX in Raw 264.7 cells. The expression of an inducible form of COX-2 by the crude and fractionated polysaccharides was also determined by gel electrophoresis of RT-PCR products with primers for COX-2. As shown in Figure 3B and C, the crude extract and fractions F1 and F2 stimulated Raw 264.7 cells to produce considerably more COX-2 mRNA than fraction F3. Therefore, this result also suggested that the increased PGE2 production by the crude extract and fractions F1 and F2 might be due to the increased expression of COX-2 that was up-regulated in activated Raw 264.7 cells and thus facilitated the release of PGE2.
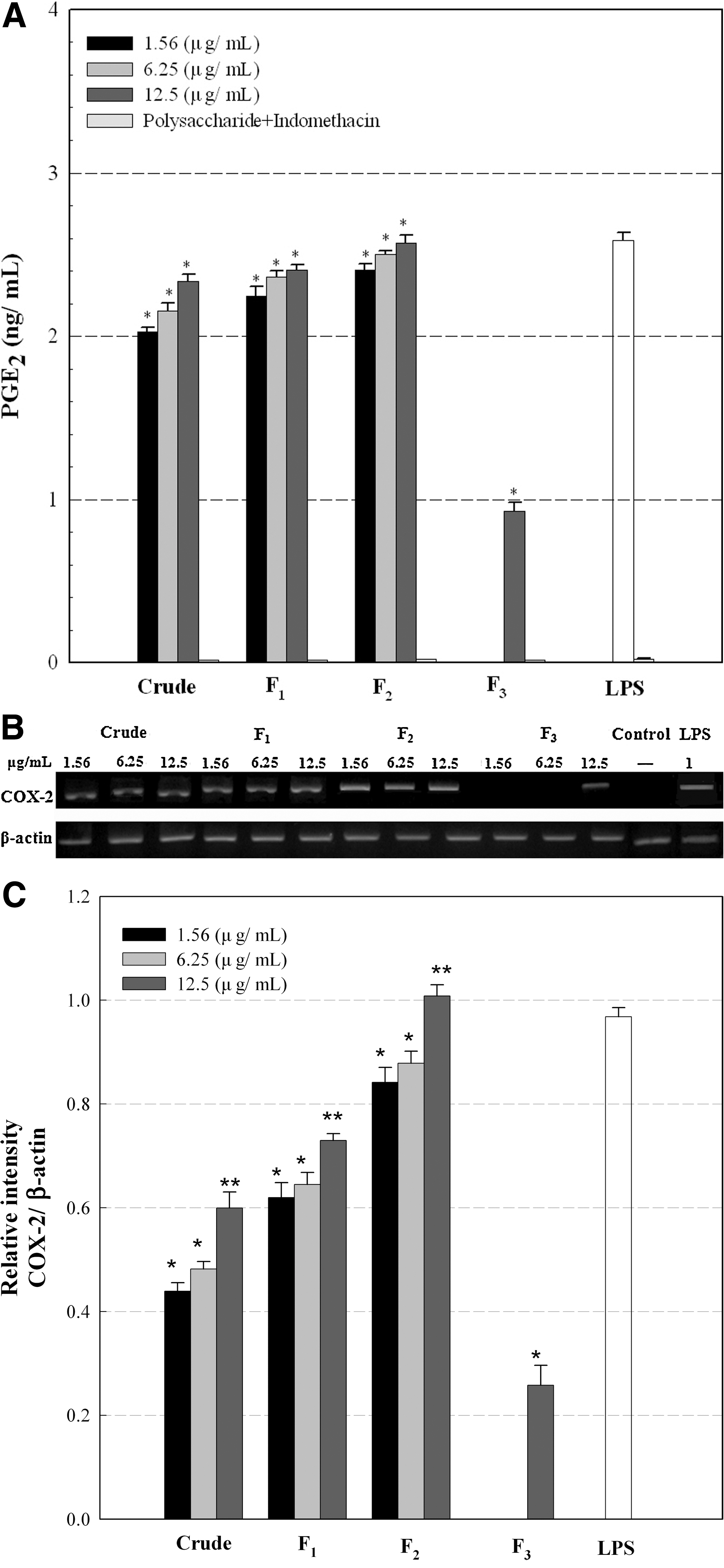
Effects of the crude and fractionated polysaccharides (F1, F2, and F3) on
A similar trend was observed in the mRNA expressions of other cytokines, including tumor necrosis factor-α, IL-6, and IL-10, by the crude and fractionated polysaccharides at the concentrations of 1.56–12.5 μg/mL (Fig. 4). The crude extract and fractions F1 and F2 increased mRNA expressions of these cytokines, whereas almost no expression of mRNAs was observed with fraction F3. It has been reported that the release of pro-inflammatory cytokines such as tumor necrosis factor-α, IL-1, IL-6, and PGE2 is essential for host survival from the injured and/or abnormal cells and tissues through the stimulation of the immune system. 27,29 However, excessive secretion of pro-inflammatory cytokines by immoderate macrophage activation could be harmful and impair repair of damaged cells and tissues, aggravating the sites by severe inflammation. Therefore, anti-inflammatory cytokines such as IL-4, IL-10, and IL-12 are also required to prevent the potential detrimental effects from excessive macrophage activation. 30 The crude extract and fractions F1 and F2 also increased the mRNA expression of IL-10 (Fig. 4). This result suggested that the released IL-10 might suppress the excessive macrophage activation, avoiding the potential aggravation at the recovering sites. The present data, therefore, suggested that the sulfated polysaccharides might be immunostimulating polysaccharides that could activate the macrophages through the production of pro-inflammatory cytokines while suppressing their overactivation by releasing anti-inflammatory cytokines. Such production of both pro-inflammatory and anti-inflammatory cytokines from Raw 264.7 cells was also observed by a sulfated galactan from Codium fragile, which implied its considerable immunostimulation without potential occurrence of severe inflammation. 10
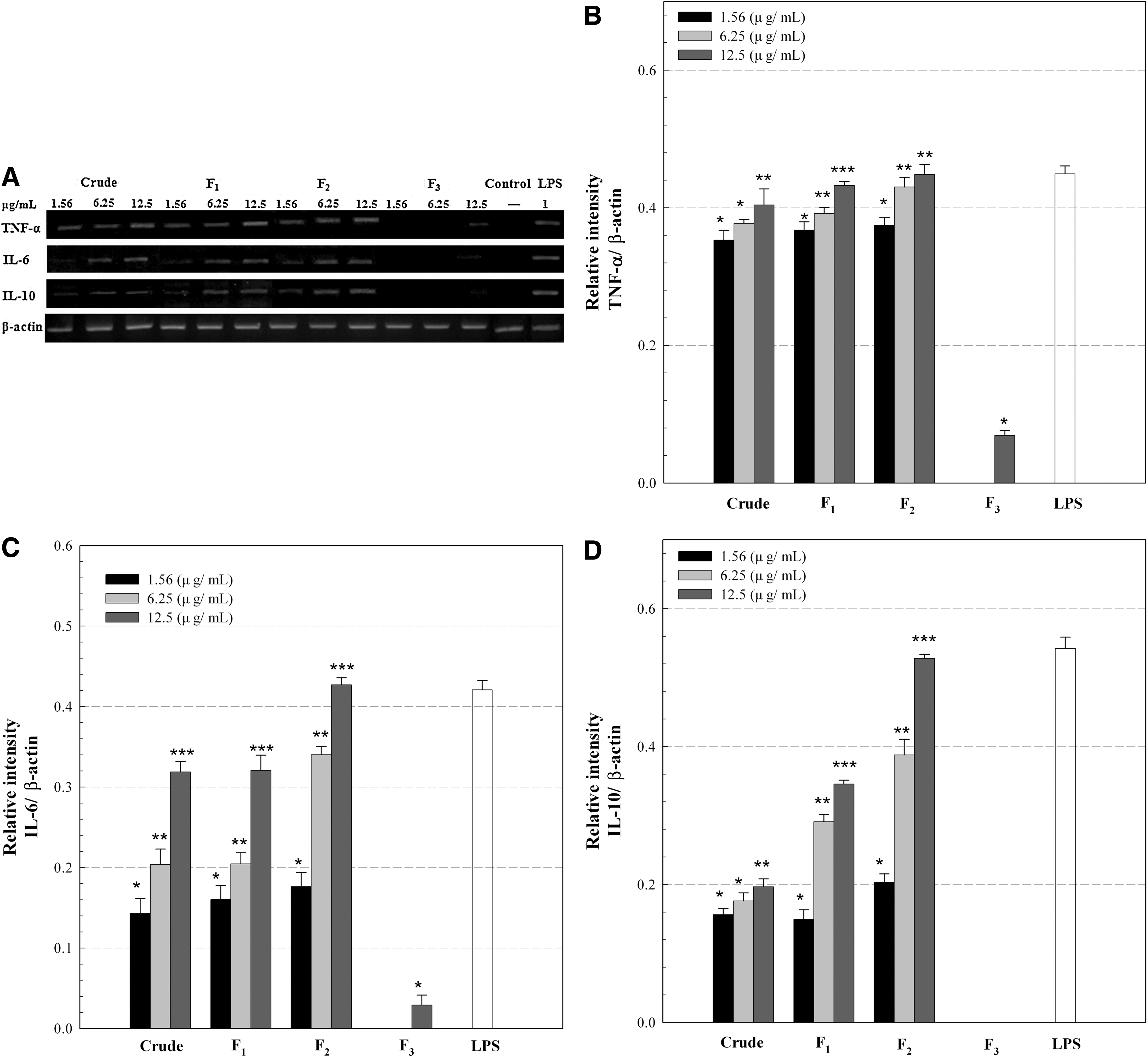
Conclusions
Sulfated polysaccharides, extracted from U. pertusa and fractionated by ion-exchange chromatography, were used to investigate their molecular characteristics and biological activities. The extracted polysaccharides consisted of mostly carbohydrates (61.4%), uronic acid (13.8%), and sulfate (13.9%) with minor amount of protein (3.4%). The ion-exchange chromatography led to the isolation of three polysaccharides having various chemical compositions and M w. Despite their few cytotoxic effects on the cancer cell line (AGS), these polysaccharides significantly induced NO as well as pro- and anti-inflammatory cytokine production from Raw 264.7 cells, suggesting that they might be strong immunostimulators. Searching for natural bioactive compounds that can stimulate the immune system has become an important research area of immunopharmacological and oncotherapeutic studies. The current study, therefore, revealed the potentials of these polysaccharides as medicinal, pharmacological, and functional food ingredients. Further studies about the structure–biological activities of the polysaccharides are now in progress to determine the relationship between the molecular structure and bioactivities.
Footnotes
Author Disclosure Statement
J.H.H. is an employee of PepsiCo Advanced Research. No competing financial interests exist for M.T., C.Y.K., and S.Y.