
Abstract
Saussurea lappa has been used in Chinese traditional medicine for the treatment of abdominal pain, tenesmus, nausea, and cancer; previous studies have shown that S. lappa also induces G2 growth arrest and apoptosis in gastric cancer cells. In this study, we investigated the effects of hexane extracts of S. lappa (HESLs) on the migration of DU145 and TRAMP-C2 prostate cancer cells. DU145 and TRAMP-C2 cells were cultured in the presence of 0–4 μg/mL HESL with or without 10 ng/mL epidermal growth factor (EGF). HESL inhibited the basal and EGF-induced migration of prostate cancer cells in a dose-dependent manner, whereas HESL did not influence the viability of these cancer cells under the conditions used in this study. Active fractions of HESL were separated via column chromatography, and the structure of the active principle was determined using 1H and 13C nuclear magnetic resonance spectroscopy. The active compound, dehydrocostus lactone (DHCL), in fraction 7 dose-dependently inhibited the basal and EGF-induced migration of prostate cancer cells. HESL and DHCL reduced matrix metalloproteinase (MMP)-9 and tissue inhibitor of metalloproteinase (TIMP)-1 secretion but increased TIMP-2 levels in both the absence and presence of EGF. Our results demonstrate that the inhibition of MMP-9 secretion and the stimulation of TIMP-2 secretion contribute to reduced migration of DU145 cells treated with HESL and DHCL. These results indicate that HESL containing its active principle, DHCL, has potential as an antimetastatic agent for the treatment of prostate cancer.
Introduction
P
The process of cancer metastasis is a multistep event, involving migration, invasion, and the destruction of intercellular matrix, intravasation into blood vessels, lymphatics, or other channels of transport, survival in the circulation, extravasation out of the vessels in the secondary site, and growth in the new location. 8 Hence, interruption of one or more of these steps can serve as an effective approach for preventing metastasis. The migration of prostate cancer cells through the basement membrane is one of the earliest events in this process and is a critical step in the metastatic spread of prostate cancer. 9 Proteolytic degradation of the extracellular matrix proteins that comprise the basement membrane is regarded as a prerequisite for the migration and invasion of cancer cells. Matrix metalloproteinases (MMPs) are typical proteolytic enzymes that are responsible for the degradation of extracellular matrix and perform crucial roles in the process of cancer metastasis. 10 MMPs are overexpressed in almost all human cancers, including prostate cancer. 10 –12 The proteolytic activities of MMPs have been shown to be regulated via the inhibition of active enzymes by endogenous inhibitors such as tissue inhibitor of metalloproteinases (TIMPs), as well as by several synthetic inhibitors. 13,14 As MMPs play important roles in metastasis, the inhibition of the activity and expression of MMPs holds great promise for the prevention or inhibition of metastasis.
The dried roots of Saussurea lappa have been widely used in traditional systems of medicine in Asia, including China, as treatments for abdominal pain, tenesmus, nausea, and cancer. 15 Previous studies have demonstrated that S. lappa exerts anti-ulcer, 16 anti-inflammatory, 17 and antitumor 18,19 effects. The principal components of S. lappa are sesquiterpene lactones such as costunolide and dehydrocostus lactone (DHCL). Several studies have reported that sesquiterpene lactones inhibit the growth of different types of cancer cells. 20 –23
We previously reported that the hexane extract of S. lappa (HESL) and its active principle, DHCL, induce apoptosis in DU145 human prostate cancer cells. 22 However, to the best of our knowledge, the effects of S. lappa on the metastasis of cancer cells have yet to be determined. In this study, we attempted to determine whether HESL and its active principle, DHCL, inhibit the migration of DU145 and TRAMP-C2 prostate cancer cells. Additionally, we attempted to ascertain whether or not HESL and DHCL influence the secretion of MMPs and TIMPs in DU145 cells.
Materials and Methods
Materials
The reagents used in this study were purchased from the following suppliers: Dulbecco's modified Eagle's medium (DMEM) and DMEM/Ham's F12 nutrient mixture (DMEM/F12) from Gibco BRL (Gaithersburg, MD, USA); fetal bovine serum (FBS), trypsin-EDTA, and penicillin/streptomycin from Cambrex Bio Technology (Walkersville, MD, USA); epidermal growth factor (EGF) from R&D Systems (Minneapolis, MN, USA); antibodies against MMP-9, TIMP-1, and TIMP-2 from Santa Cruz Biotechnology (Santa Cruz, CA, USA); horseradish peroxidase–conjugated anti-rabbit, anti-mouse, and anti-goat immunoglobulin G from Amersham Biosciences (Arlington Heights, IL, USA); and Immobilon™ western chemiluminescent horseradish peroxidase substrate and Centricon Plus-20 filters from Millipore Corp. (Billerica, MA, USA). Unless otherwise noted, all other materials were obtained from Sigma-Aldrich Co. (St. Louis, MO, USA).
Preparation of extract, fractionation, and identification of active compounds
The dried roots of S. lappa Clarke were purchased from a local drug store in Chuncheon, Korea, and authenticated by Professor Emeritus Hyung Jun Ji (Seoul National University, Seoul, Korea). HESL was prepared and fractionated via column chromatography as described previously. 22 The ability of each fraction to inhibit DU145 cell migration was evaluated using transwell migration assay as described below. Active fraction 7 (F7), which evidenced the highest levels of activity, was further purified by repeating the vacuum liquid chromatography procedure on silica as described previously. 22 The structure of the active compound in the active fraction (F7) was previously identified. 22
Cell culture
DU145 human prostate cancer cells and TRAMP-C2 transgenic mouse prostate cancer cells were acquired from the American Type Culture Collection (Manassas, VA, USA) and maintained in DMEM/F12 and DMEM, respectively, containing 100 mL/L FBS, 100 kU/L penicillin, and 100 mg/L streptomycin in a humidified atmosphere of 5% CO2 at 37°C.
Transwell migration assay
The migration ability of DU145 and TRAMP-C2 cells was measured as previously described in detail. 24 Serum-deprived DU145 cells were plated in 6.5-mm Transwell inserts in 24-well plates at 2.5×104 cells per filter and treated for 4 hours with various concentrations of HESL or DHCL in serum-deprivation medium (DMEM/F12+1% charcoal-stripped FBS). Serum-deprived TRAMP-C2 cells were plated in 6.5-mm Transwell inserts in 24-well plates at 2.0×104 cells per filter and treated for 1 hour with various concentrations of HESL or DHCL in serum-deprivation medium (DMEM + 1% charcoal-stripped FBS). Bovine serum albumin (0.1%) and/or EGF (10 ng/mL) were used as chemoattractants. Cells that had migrated were stained with hematoxylin and eosin.
Wound migration assay
The wound migration assay was conducted as previously described. 24 DU145 cells were incubated with 0 μg/mL HESL, 4 μg/mL HESL, 10 ng/mL EGF, and 10 ng/mL EGF + 4 μg/mL HESL or 0 μg/mL DHCL, 2 μg/mL DHCL, 10 ng/mL EGF, and 10 ng/mL EGF + 2 μg/mL DHCL in DMEM/F12 containing 10 mL/L charcoal-stripped FBS for 0, 6, or 12 hours. Cell migration was observed via microscopy at the indicated time points. The measured width of injury was plotted as a percentage of the width at 0 hour.
Preparation of conditioned media and western blot analysis
Cells were serum-starved in DMEM/F12 for 24 hours and treated for 12 hours with various concentrations of HESL or DHCL in the absence or presence of 10 ng/mL EGF. Media conditioned for 12 hours were collected and concentrated via centrifugal ultrafiltration using a Centricon Plus-20 filter unit with a molecular exclusion of 10,000. The proteins of the concentrated conditioned media were then determined with a BCA protein assay kit (Pierce, Rockford, IL, USA). The volumes of media loaded onto the gel were adjusted for equivalent protein levels. Western blot analyses were conducted as described previously. 25 Signals were detected via an enhanced chemiluminescence method using Immobilon western chemiluminescent horseradish peroxidase substrate. The relative abundance of each band was quantified using the Bio-profile Bio-1D application (Vilber-Lourmat, Marne-la-Vallée, France).
Statistical analysis
The data were expressed as mean±SEM values and analyzed via analysis of variance. The differences between the treatment groups were assessed via Duncan's multiple range test, using the SAS system software for Windows, version 9.1 (SAS Institute, Cary, NC, USA). Differences were considered significant at P<.05.
Results
HESL inhibits both basal and EGF-stimulated migration of prostate cancer cells
We first examined the effects of different concentrations (1–4 μg/mL) of HESL on DU145 and TRAMP-C2 cell migration. The Transwell migration assay revealed that HESL markedly inhibited the migration of DU145 and TRAMP-C2 cells at 4 hours and 1 hour, respectively. Treatment of DU145 and TRAMP-C2 cells with 4 μg/mL HESL resulted in a 76.5% and 59.4% reduction in the numbers of migrated cells, respectively (Fig. 1A and 1B, respectively).
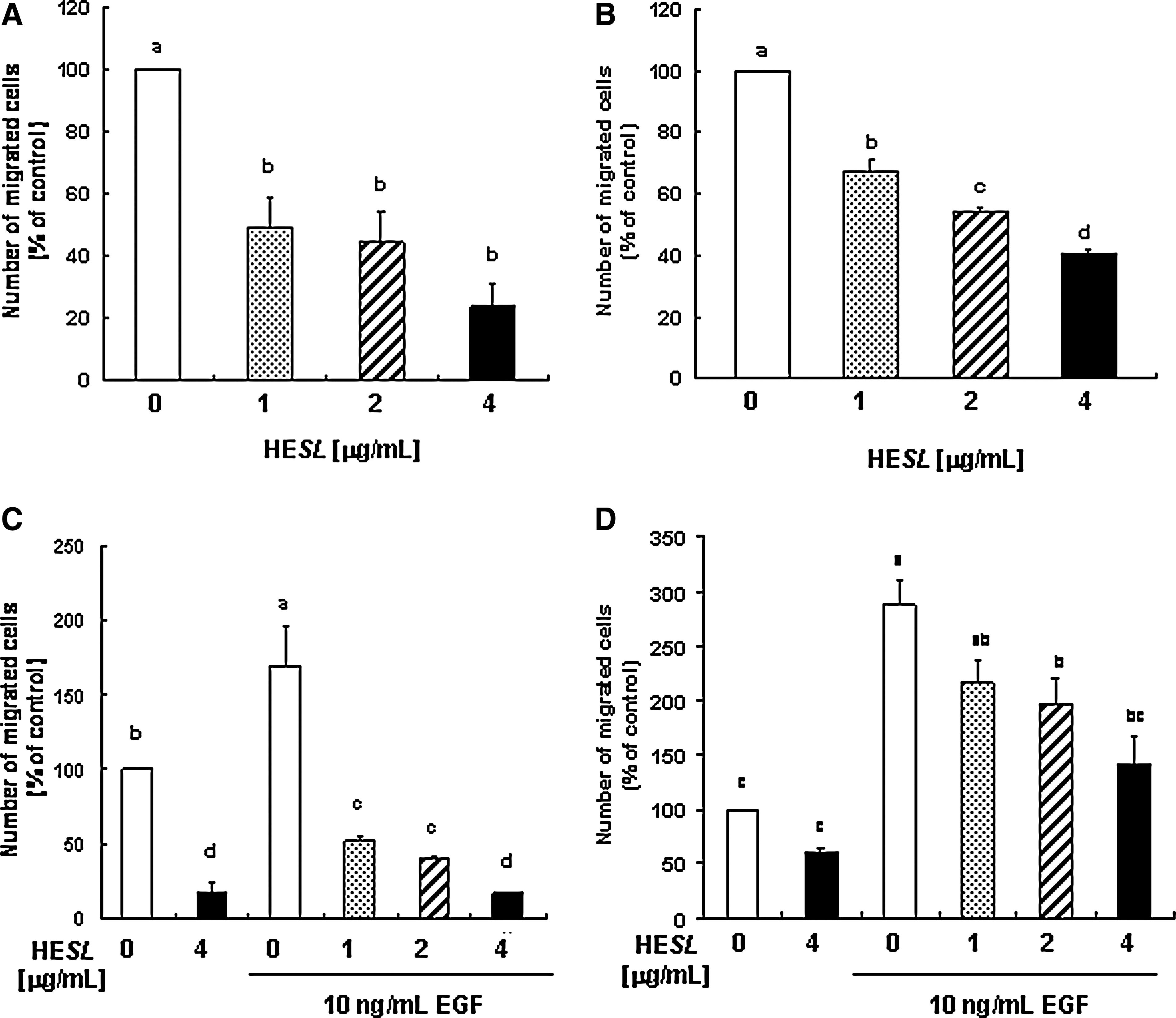
Hexane extract of S. lappa (HESL) inhibits basal and epidermal growth factor (EGF)-stimulated migration of prostate cancer cells.
EGF and EGF receptor are up-regulated in human prostate cancer 26 and also play a pivotal role in up-regulating motility during prostate cancer invasion. 26,27 Kwon et al. 24 previously reported that EGF stimulated the cell migration and invasion of DU145 cells, whereas insulin-like growth factor-I or heregulin exerted no such effect. We subsequently attempted to determine whether HESL reduces the EGF-stimulated migration of DU145 and TRAMP-C2 cells. The number of migrated cells was increased in EGF-treated DU145 and TRAMP-C2 cells, and these increases were noticeably reduced by HESL treatment (Fig. 1C and 1D, respectively). Results from the wound migration assay showed that HESL significantly inhibited both the basal and EGF-stimulated migration of DU145 cells (Fig. 2).
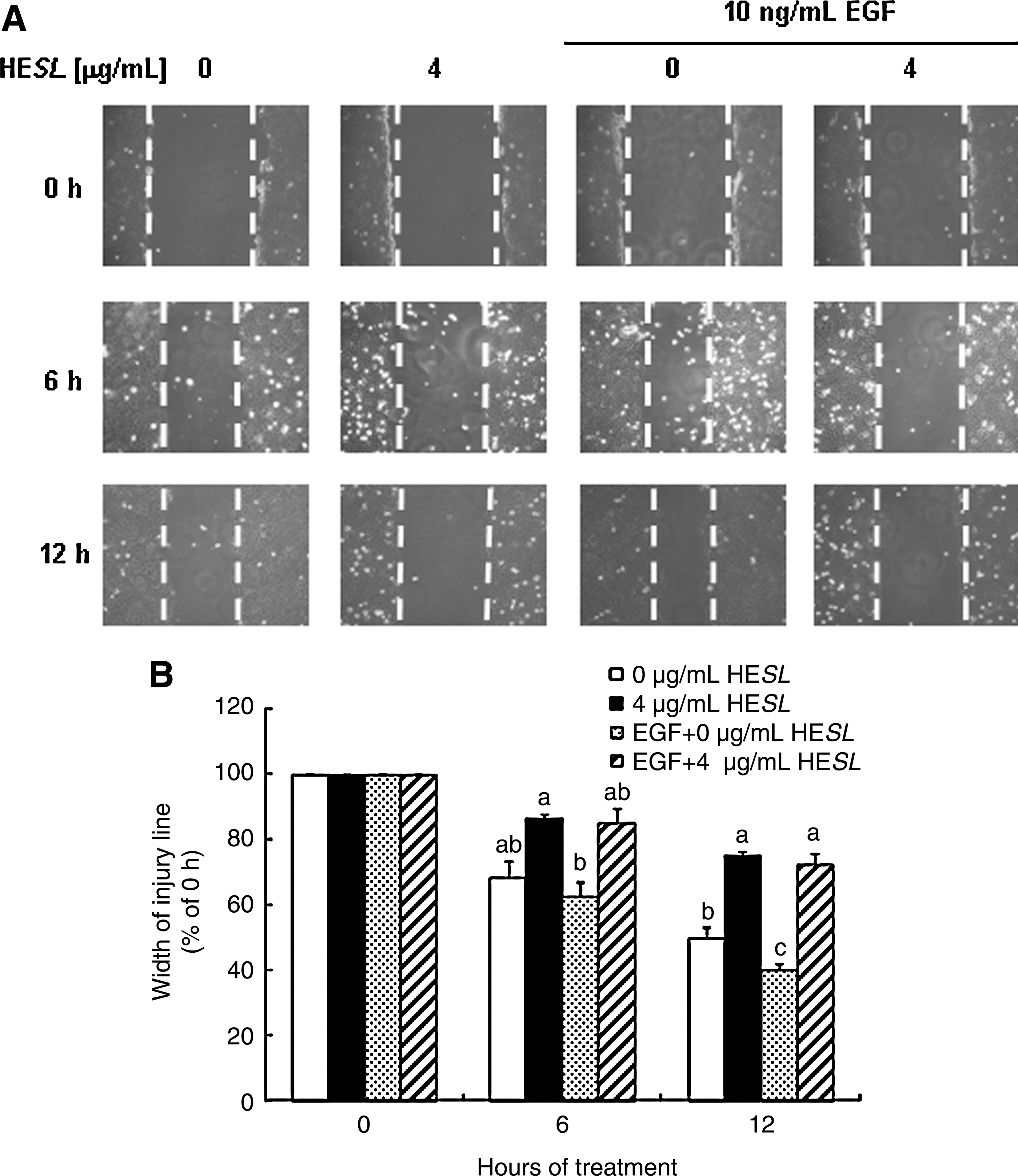
HESL inhibits basal and EGF-induced migration of wounded DU145 cells. DU145 cells were plated in six-well plates at 8×104 cells per well in Dulbecco's modified Eagle's medium/F12 supplemented with 10% fetal bovine serum. One day later, the monolayers were serum-deprived in Dulbecco's modified Eagle's medium/F12 containing 1% charcoal-stripped fetal bovine serum for 24 hours. Cell populations with 90% confluency were incubated for 1 hour with 1 μg/mL mitomycin C in Dulbecco's modified Eagle's medium/F12 containing 1% charcoal-stripped fetal bovine serum. After mitomycin C treatment, the injury line was made with a yellow tip and rinsed with phosphate-buffered saline. The cells were incubated with 0 μg/mL HESL, 4 μg/mL HESL, 10 ng/mL EGF, or 10 ng/mL EGF + 4 μg/mL HESL in Dulbecco's modified Eagle's medium/F12 containing 1% charcoal-stripped fetal bovine serum for 0, 6, or 12 hours.
We observed previously that the treatment of DU145 cells with 4 μg/mL HESL for 24 and 48 hours resulted in G1 cell cycle arrest and apoptosis, respectively. 22 We determined cell viability at 12 hours and noted that HESL did not affect the viability of DU145 and TRAMP-C2 cells at the concentrations used in this study (data not shown). We also conducted the wound migration assay in the presence of mitomycin C to eliminate the contribution of proliferation.
HESL decreases secretion of MMP-9 and TIMP-1 but increases TIMP-2 secretion in basal and EGF-stimulated DU145 cells
To determine the effects of HESL on secretion of MMP-9 and TIMPs in DU145 cells, the cells were treated for 12 hours with various concentrations of HESL in the absence or presence of EGF, and concentrated conditioned media were prepared for western blot analysis. As is shown in Figure 3, EGF significantly stimulated MMP-9 and TIMP-1 secretion, and HESL inhibited the secretion of these proteins, regardless of whether or not the cells were treated with EGF. The TIMP-2 secretion levels were slightly decreased by EGF but were increased by treatment with HESL regardless of EGF treatment (Fig. 3).
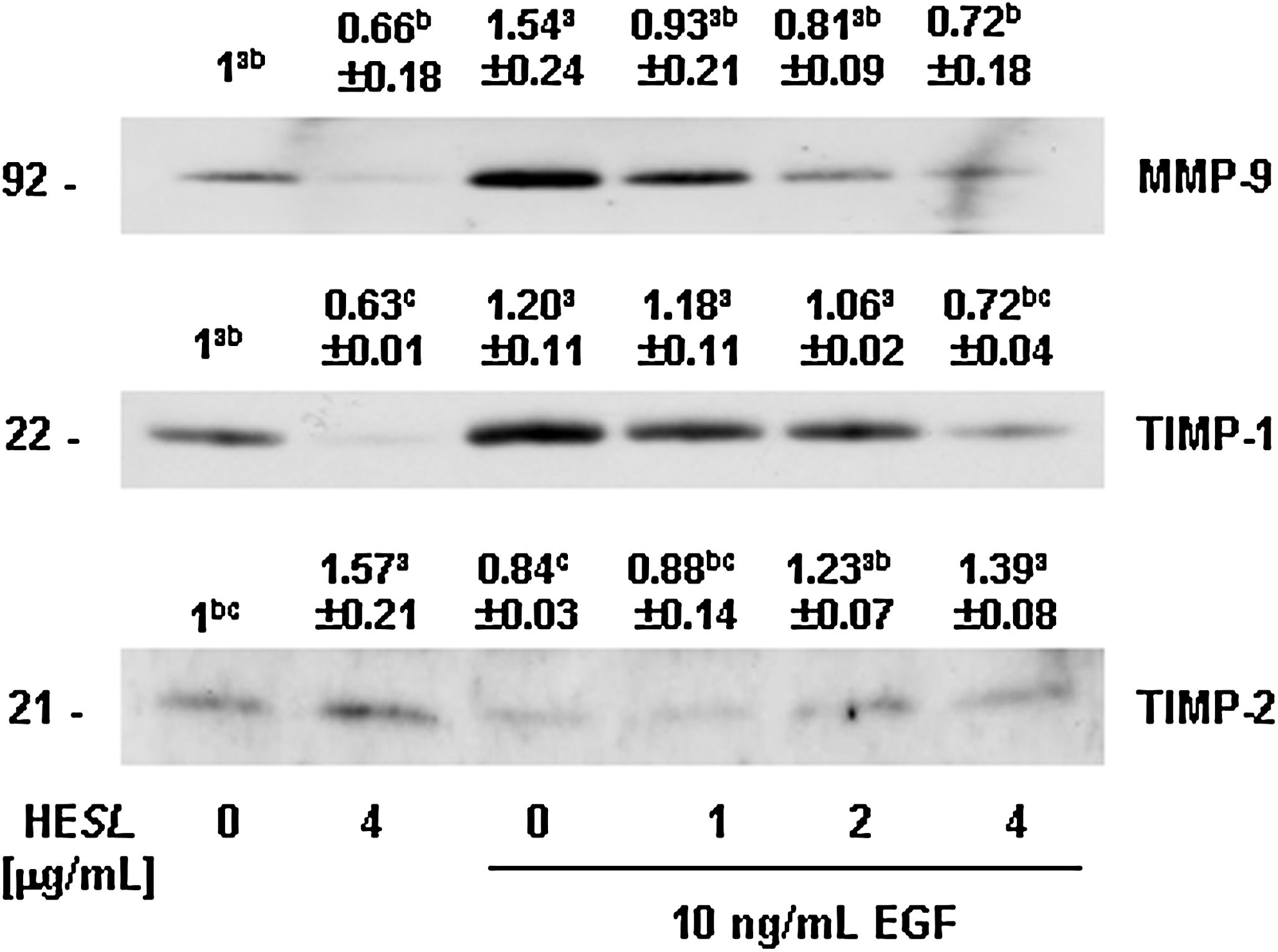
HESL reduces matrix metalloproteinase (MMP)-9 and tissue inhibitor of metalloproteinase (TIMP)-1 secretion but increases TIMP-2 secretion in DU145 cells. Cells were plated in 100-mm-diameter dishes at 2×106 cells per dish in Dulbecco's modified Eagle's medium/F12 supplemented with 10% fetal bovine serum, serum-starved in Dulbecco's modified Eagle's medium/F12, and then treated with 0–4 μg/mL HESL in Dulbecco's modified Eagle's medium/F12 containing 0 or 10 ng/mL EGF for 12 hours. Media conditioned for 12 hours were collected and concentrated for western blotting. Photographs of chemiluminescent detection of the immunoblots, which are representative of three or four independent experiments, are shown. The relative abundance of each band was quantified, and the control levels were set to 1. The adjusted mean±SEM (n=3 for MMP-9 and TIMP-1; n=4 for TIMP-2) of each band is shown above each blot. abcMeans without a common letter differ, P<.05.
Identification of DHCL as an active compound in HESL
To identify the active principle of HESL that inhibits cell migration, we separated the fractions of HESL via column chromatography in vacuo with silica gel (70–230 mesh, Merck, Darmstadt, Germany) 22 and evaluated the ability of the various fractions to suppress DU145 cell migration. Among the 15 fractions eluted by gradient systems of n-pentane–n-hexane–ethyl acetate (10:0:0–0:5:5 by volume), F7 was determined to most potently decrease DU145 cell migration (Fig. 4). F7 was also the most potent fraction for reducing the number of viable cells. We previously determined the structure of the active compound in the F7 fraction and identified it as DHCL (Fig. 4B). The analytical properties of DHCL have been evaluated previously. 22
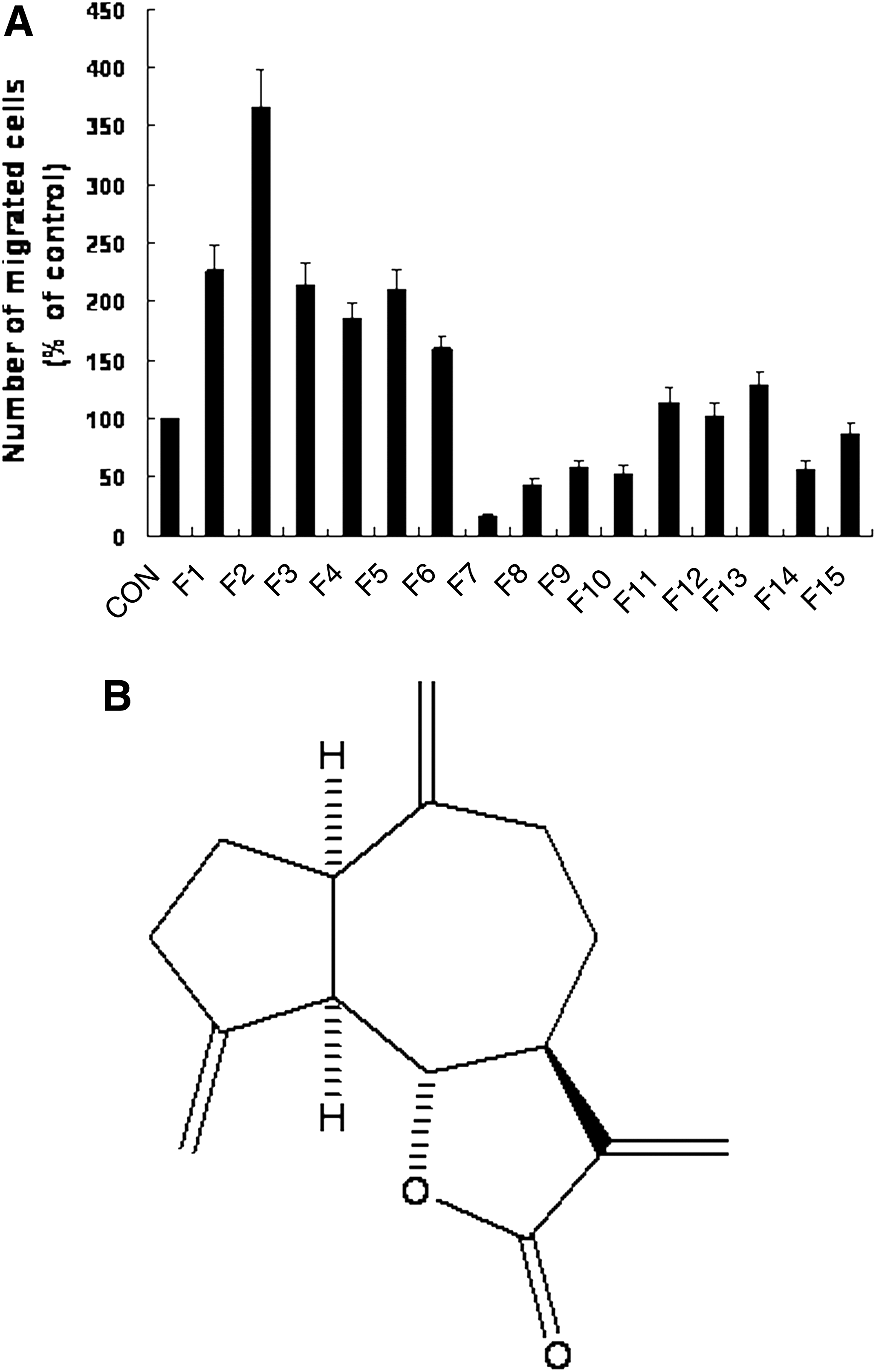
Fraction 7 from HESL is the most potent inhibitor of DU145 cell migration. Cells were serum-deprived in Dulbecco's modified Eagle's medium/F12 containing 1% charcoal-stripped fetal bovine serum for 24 hours. The lower side of a 6.5-mm-diameter Transwell filter was coated with type IV collagen. Serum-deprived cells were plated into the upper compartment at 2.5×104 cells per well and treated with different fractions (F) of HESL at 4 μg/mL. The lower compartment was filled with Dulbecco's modified Eagle's medium/F12 containing 1% charcoal-stripped fetal bovine serum and 0.1% bovine serum albumin. Cells were incubated for 4 hours. Migrated cells were stained with hematoxylin and eosin. Data are mean±SEM values (n=3, P<.05).
DHCL inhibits basal and EGF-induced migration of prostate cancer cells
We examined the effects of different concentrations (0.5–2 μg/mL) of DHCL on the migration of DU145 and TRAMP-C2 cells. DHCL dose-dependently inhibited the migration of DU145 and TRAMP-C2 cells (Fig. 5A and 5B, respectively). We next determined whether DHCL reduces the EGF-stimulated migration of DU145 and TRAMP-C2 cells. The migrations of DU145 and TRAMP-C2 cells were markedly increased by EGF treatment, and these increases were distinctly reversed by DHCL treatment (Fig. 5C and 5D, respectively). Additionally, the results from the wound migration assay demonstrated that DHCL noticeably inhibited both the basal and EGF-stimulated migration of DU145 cells (Fig. 5E). Like HESL, DHCL significantly decreased basal and EGF-induced MMP-9 and TIMP-1 secretion in DU145 cells. TIMP-2 secretion levels in the DU145 cells were increased dramatically as the result of treatment with 2 μg/mL DHCL, regardless of EGF treatment (Fig. 5F).

Dehydrocostus lactone (DHCL) inhibits basal and EGF-induced migration of prostate cancer cells. The migration of
Discussion
Recently, natural compounds derived from medicinal plants have attracted an increasing amount of attention as excellent sources of chemopreventive and/or chemotherapeutic agents (reviewed by Gullett et al. 28 ). S. lappa is a well-known medicinal plant with a long history of pharmaceutical use in Asia and has been previously reported to inhibit the growth of various cancer cells. 18,19,21 In our previous study, we reported that HESL inhibits cell cycle progression and induces apoptosis. 22 In this study we demonstrated that HESL inhibits the migration of DU145 and TRAMP-C2 cells. Because HESL has multiple targets in prostate cancer cells, it is a worthy candidate as an agent for the treatment/prevention of prostate cancer.
The lethality of most cancers is the result of metastasis from primary cancers to other tissues. 7 Therefore, it is important to determine the antimetastatic effects of various natural compounds in order to develop safe and effective chemopreventive agents. In this study, we determined the effects of HESL on the migration of prostate cancer cells in an attempt to explore the antimetastatic effects of HESL, as the migration of cancer cells is one of the most important steps in the cancer metastasis process. 8 We noted that HESL effectively inhibited the migration of DU145 and TRAMP-C2 prostate cancer cells and that DHCL is the active compound in HESL. To the best of our knowledge, this is the first study to demonstrate that HESL and DHCL exert antimetastatic effects in prostate cancer cells. In our previous study we also observed that HESL and DHCL inhibited cell cycle progression and induced apoptosis when DU145 cells were incubated for 24 and 48 hours, respectively. Neither HESL nor DHCL suppressed cell viability or induced apoptosis within 1, 4, or 12 hours at the concentrations used herein (data not shown). Collectively, these findings indicate that the migration of prostate cancer cells is much more sensitively affected by HESL and DHCL than are the cell cycle progression and apoptosis characteristics of these cells. Future animal studies will be required to determine whether or not HESL and DHCL inhibit the metastasis of prostate cancer cells in vivo.
Tumor metastasis is largely regulated by extracellular signals, including growth factors such as EGF. EGF signaling is associated closely with the progression of prostate cancer (reviewed by Lu and Kang 29 ). The up-regulation of EGF/EGF receptor expression has been previously demonstrated in metastatic prostate cancer. 30 Exogenous EGF has been determined to enhance the migration and invasion behaviors of various prostate cancer cell lines, including Mat-LyLu cells, 31 PC-3M cells, 32 and DU145 cells. 24 Therefore, it is reasonable to surmise that the natural compounds that suppress EGF signaling could be used to inhibit the progression of prostate cancer. In this study, we noted that EGF markedly stimulated the migration of DU145 and TRAMP-C2 cells and that this migration was effectively inhibited by HESL and DHCL. These results demonstrate that HESL and DHCL both have potential as inhibitors of prostate cancer progression.
The MMPs are a group of zinc-dependent endopeptidases responsible for the proteolysis of the basement membrane and extracellular matrix proteins. It was previously reported that enhanced MMP production is correlated with invasion, metastasis, and angiogenesis of tumors. 33 Among the several known MMPs, MMP-9 (92-kDa gelatinase B) degrades type IV collagen and gelatin, the major components of the basement membrane and extracellular matrix. MMP-9 is secreted by various tumor cells, and the levels of this enzyme correlate with the metastatic potential of tumor cells. 33 Several antimetastatic compounds have been shown to inhibit MMPs via different mechanisms, including the inhibition of expression of the genes for MMP proteins, interference in proenzyme activation, and the direct inhibition of proteolytic enzyme activity. 34 Thus, the inhibition of the expression and activity of MMP-9 is regarded as an intriguing target for cancer treatment. In this study, EGF was shown to stimulate the secretion of MMP-9, and HESL and DHCL effectively inhibited both the basal and EGF-stimulated secretion of this protease in DU145 cells, thereby indicating that the decrease in MMP-9 contributes to the reduction in migration of HESL- and DHCL-treated DU145 cells.
Results from clinical trials using synthetic MMP inhibitors have generally proven disappointing, primarily because of a lack of overall response and dose-limiting toxicity. 35,36 Therefore, TIMPs, natural inhibitors of MMPs, are currently being considered as a promising class of target molecules for cancer therapy. Among the four members of the TIMP family, tumor cells have been shown to manufacture primarily TIMP-1 and TIMP-2. In this study, we determined that EGF did not affect TIMP-2 secretion but that HESL and DHCL significantly increased TIMP-2 secretion. This suggests that the increased TIMP-2 may inhibit MMP activity, thereby resulting in a suppression of the migration of the HESL- and DHCL-treated DU145 cells. Unlike TIMP-2, TIMP-1 secretion is increased by EGF, and HESL and DHCL inhibit both basal and EGF-induced TIMP-1 secretion. It has been previously reported that EGF induces TIMP-1 expression and increases invasion in FTC-133 thyroid carcinoma cells 37 and DU145 cells. 24 Furthermore, other studies have demonstrated that TIMP-1 is involved in the cell survival pathways via its interaction with the CD63/integrin-β1 complex and that these functions occur independently of MMP inhibition. 38,39 Kim et al. 40 demonstrated increased levels of TIMP-1 in the sera and lungs of BALB/c mice when lung metastasis was induced by the injection of 4T1 breast cancer cells. Additionally, it has been previously reported that high serum TIMP-1 levels are correlated with poor progression of several cancer types. 41,42 These results indicate that TIMP-1 may enhance, rather than suppress, cancer metastasis.
In conclusion, the findings of this study demonstrate that HESL inhibits the migration of DU145 and TRAMP-C2 prostate cancer cells, possibly through the down-regulation of MMP-9 and TIMP-1 and the up-regulation of TIMP-2. Additionally, the findings of this study show that the antimetastatic effect of HESL is attributable, at least in part, to the effects of DHCL. These results implicate HESL containing DHCL as a powerful candidate for a preventive and/or therapeutic agent against prostate cancer metastasis. Future in vivo efficacy studies with HESL and DHCL in preclinical prostate cancer models are warranted.
Footnotes
Acknowledgments
This work was supported by a National Research Foundation Grant funded by the Korean Government (The Regional Core Research Program/Medical & and Bio-Material Research) and supported by the Ministry of Knowledge Economy through the Center for Efficacy Assessment and Development of Functional Foods and Drugs at Hallym University, Korea.
Author Disclosure Statement
The authors have no conflicts of interests associated with this study to declare.