
Abstract
Oxidative stress has become widely viewed as an underlying condition in diseases such as ischemia/reperfusion disorders, central nervous system disorders, cardiovascular disease, cancer, diabetes, etc. The role that antioxidants play in the process of carcinogenesis has recently gained considerable attention. β-Sitosterol, a naturally occurring sterol molecule, is a relatively mild to moderate antioxidant and exerts beneficial effects in vitro by decreasing the level of reactive oxygen species. The present study evaluated the antioxidant potential of β-sitosterol in 1,2-dimethylhydrazine (DMH)-induced colon carcinogenesis. The enzymatic and nonenzymatic antioxidants and lipid peroxides in colonic and hepatic tissues were evaluated. Generation of reactive oxygen species, beyond the body's endogenous antioxidant capacity, causes a severe imbalance of cellular antioxidant defense mechanisms. Elevated levels of liver lipid peroxides by DMH induction were effectively decreased by β-sitosterol supplementation. β-Sitosterol also exhibited a protective action against DMH-induced depletion of antioxidants such as catalase, superoxide dismutase, glutathione peroxidase, glutathione reductase, glutathione S-transferase, and reduced glutathione in colonic and hepatic tissues of experimental animals. Supplementation with β-sitosterol restored the levels of nonenzymatic antioxidants (vitamin C, vitamin E, and glutathione). Histopathological alterations in DMH-induced animals were restored to near normal in rats treated with β-sitosterol. Thus, β-sitosterol by virtue of its antioxidant potential may be used as an effective agent to reduce DMH-induced oxidative stress in Wistar rats and may be an effective chemopreventive drug for colon carcinogenesis.
Introduction
C
Antioxidant enzymes, the main scavengers of free radicals, are modulated during carcinogenesis or after tumor formation. Changes in these enzymes may result in malignancy due to free radical damage, and restoration of their activities could help to reverse malignancy by scavenging free radicals and preventing further malignancy. 4 Cellular antioxidant systems can be divided into two major groups: enzymatic and nonenzymatic. Enzymatic scavengers include superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), glutathione reductase (GR), and glutathione S-transferase (GST). Reduced glutathione (GSH), vitamin C, and vitamin E are the nonenzymatic antioxidants involved in detoxification of the toxic ROS produced during malignancy. 5
The 1,2-dimethylhydrazine (DMH)-induced colon cancer model has been well characterized and bears many of the same cell kinetics and histopathological and molecular characteristics of tumorigenesis as human colon cancer, as well as being morphologically similar to it. 6 A high fat diet is thought to act, at least in part, by increasing the intracolonic concentrations of bile acids and fatty acids. 7 Increased intraluminal bile acids as a potential consequence of high fat, high meat/low fiber diets are believed to be a major causative factor in the formation of colon cancer. In animal studies, dietary fat appears to act as a promoter rather than as an initiator of colon carcinogenesis. 8 Antioxidant enzymes, the main scavengers of free radicals, are modulated during carcinogenesis or after tumor formation. As changes in these enzymes result in malignancy, the recovery of decreased activities of the enzymes could help to reverse the malignancy. 4 The chemopreventive mechanism in experimentally induced colon carcinogenesis is thought to be as a result of antioxidant activity, by the modulation of the enzymes involved in metabolic activation and excretion of carcinogens. 9
β-Sitosterol is the second major phytosterol, having a structure similar to that of cholesterol, and is present in almost all plant foods, especially plant oils, seeds and nuts, cereals, and legumes. β-Sitosterol inhibits the growth of several specific types of tumor cells in vitro. β-Sitosterol isolated from various plants promotes apoptosis by increasing Fas levels and caspase-8 activity, 10 phosphorylation of extracellular-signal regulating kinase and p38 mitogen-activated protein kinase, 11 inhibition of cancer cell proliferation (even at low concentrations with no cytotoxic effect to noncancerous cells), 12 modulation of antioxidant enzyme levels in pathogenesis, 13 arrest of cells at the G2/M phase in prostate cancer cells, 14 and decreasing free radical generation in vitro. 15 A large number of medicinal plants and their purified constituents have beneficial therapeutic potential. We previously reported that β-sitosterol exhibits chemopreventive potential in DMH-induced animals because of its in vitro antioxidant activity and decrease in numbers of aberrant crypts in DMH-induced animals. 16 These results suggest that β-sitosterol may prevent cancer development. We therefore examined the effect of β-sitosterol supplementation on DMH-induced rat colon carcinogenesis using tissue lipid peroxidation and antioxidant levels as biomarkers.
Materials and Methods
Chemicals
In this study, extraction, separation, and purification of β-sitosterol were performed as reported previously 16 from Asclepias curassavica. DMH, hematoxylin, and eosin were purchased from Sigma Chemical Co. (Bangalore, India). All other chemicals, including solvents, were of high purity and analytical grade marketed by Himedia Chemicals (Mumbai, India).
Animals
Experiments were carried out in male albino Wistar rats, 5 weeks old, obtained from the Central Animal House, Kings Institute, Chennai, Tamil Nadu, India. In total, 60 animals were divided into six groups of 10 animals each. The animals were cared for in compliance with the principles and guidelines of the Ethical Committee for Animal Care and Institutional Animal Ethical Committee in accordance with the Indian National Law on Animal Care and Use (Regulation Number 833/a/2 004/CPC SEA). The animals were housed four per cage in polypropylene cages with a wire mesh top and a hygienic bed of husk in a specific pathogen-free animal room under controlled conditions of 12-h light/12-h dark cycle, temperature of 24±2°C, and relative humidity of 50±10%. The rats were held in quarantine for 1 week and had access to food and tap water ad libitum. Commercial pellet diet containing 4.2% fat (Hindustan Lever Ltd., Mumbai) was powdered and mixed with 15.8% peanut oil, making a total of 20% fat in the diet (Table 1). This modified powdered pellet diet was fed to rats in all groups throughout the experimental period of 16 weeks.
Administration of carcinogen
The animals in Groups III–VI received subcutaneous injections of DMH at a dose of 20 mg/kg of body weight once a week for the first four consecutive weeks. Prior to subcutaneous injection, DMH was dissolved in 1 mM EDTA; the pH was adjusted to 6.5 with 1 mM sodium hydroxide to ensure the pH and stability of the chemical. The solution was used immediately after preparation.
Initial body weights of all animals were between 80 and 120 g. The animal weights were recorded once a week throughout the experimental period and prior to sacrifice.
Treatment of animals with β-sitosterol
Group I rats received modified pellet diet along with intragastric intubation of 0.1% carboxymethyl cellulose (CMC) (1.0 mL) throughout the experimental period.
Group II rats received modified pellet diet plus 20 mg/kg of body weight β-sitosterol suspended in 0.1% CMC (1.0 mL) p.o. every day throughout the experimental period.
Group III rats were administered 20 mg/kg of body weight DMH (carcinogen) s.c. once a week for four consecutive weeks and kept without any treatment for the next 12 weeks.
Group IV animals were treated as in Group III along with β-sitosterol (5 mg/kg of body weight, p.o.) suspended in 0.1% CMC supplemented throughout the entire experimental period of 16 weeks.
Group V animals were treated as in Group III along with β-sitosterol (10 mg/kg of body weight, p.o.) suspended in 0.1% CMC supplemented throughout the entire experimental period of 16 weeks.
Group VI animals were treated as in Group III along with β-sitosterol (20 mg/kg of body weight, p.o.) suspended in 0.1% CMC supplemented throughout the entire experimental period of 16 weeks.
At the end of 16 weeks all the animals were sacrificed under anesthesia (intraperitoneal administration of ketamine hydrochloride, 30 mg/kg of body weight), by cervical dislocation between 8 a.m. and 10 a.m. after an overnight fast. Tissue samples were immediately transferred to ice-cold containers, weighed, and homogenized using appropriate buffer in a tissue homogenizer. Tumor and normal colon tissues were fixed in 10% formalin and were stained with hematoxylin and eosin for histopathological investigation. Total lipid peroxidation levels, as evidenced by the formation of thiobarbituric acid–reactive substances (TBARS), were estimated by the method of Yagi 17 and GSH by the method of Ellman. 18 Diene conjugates (DCs) were estimated by the method of Recknagel and Glende, 19 and lipid hydroperoxides (LOOHs) were estimated by the method of Jiang et al. 20 The activities of GPx and GST were estimated by the method of Rotruck et al. 21 and Habig et al., 22 respectively. SOD was assayed by the method of Kakkar et al., 23 and CAT was assayed by the method of Sinha. 24 The nonenzymatic antioxidants, vitamins C and E, in hepatic and colon tissue were determined by the method of Omaye et al. 25 and Desai, 26 respectively. Tissue protein was determined by the method of Lowry et al. 27
Statistical analysis
The statistical significance of the data was determined using one-way analysis of variance, and significant difference among treatment groups were evaluated by Duncan's Multiple Range Test. The results were considered statistically significant at P<.05. All statistical analyses were made using the SPSS version 11.0 software package (SPSS, Tokyo, Japan).
Results
The lipid peroxidation levels of hepatic tissue are given in Table 2 for the experimental and control animals. Lipid peroxidation indicated by DCs, LOOHs, and TBARS was significantly higher in hepatic tissues of DMH-induced animal models compared with control animals. Supplementation of β-sitosterol at 20 mg/kg of body weight for the entire period of study significantly reduced the levels of lipid peroxidation markers in hepatic tissues of DMH-induced animals.
Data are mean±SD values of 10 rats in each group.
Values not sharing a common superscript letter in a column are significantly different at the P<.05 level by Duncan's Multiple Range Test.
P<.01, values significantly different compared with the 1,2-dimethylhydrazine alone–treated group (Group III).
DCs, diene conjugates; LOOHs, lipid hydroperoxides; TBARS, thiobarbituric acid–reactive substances.
Tables 3 and 4 show the levels of lipid peroxidation and antioxidant status in proximal colon, distal colon, and intestine of control and DMH-induced animals. The colonic lipid peroxidation markers were found to be significantly decreased in experimental animals induced with DMH. Supplementation with β-sitosterol at the dose of 20 mg/kg of body weight for the entire period of study significantly increased the levels of lipid peroxidation markers to near normal in colonic tissues of DMH-induced animals.
Data are mean±SD values of 10 rats in each group.
P<.05, values not sharing a common superscript letter are significantly different.
P<.01, values significantly different compared with the 1,2-dimethylhydrazine alone–treated group (Group III).
Data are mean±SD values of 10 rats in each group. Superoxide dismutase (SOD) activity is given in amount of enzyme required for 50% inhibition of nitro blue tetrazolium reduction/min/mg of protein. Catalase (CAT) activity is given in μmol of H2O2 utilized/min/mg of protein. Glutathione reductase (GR) activity is given in μmol of NADPH oxidized/min/mg of protein. Glutathione peroxidase (GPx) activity is given in μmol of reduced glutathione utilized/min/mg of protein. Glutathione S-transferase (GST) activity is given in μmol of 1-chloro-2,4-dinitrobenzene–reduced glutathione conjugate formed/min/mg of protein.
P<.05, values not sharing a common superscript letter are significantly different.
P<.01, values significantly different compared with the 1,2-dimethylhydrazine alone–treated group (Group III).
Table 5 shows the enzymatic antioxidant status of control and DMH-induced animals in hepatic tissue. The activities of the enzymatic antioxidants SOD, CAT, GR, GPx, and GST in hepatic tissue were significantly lowered in DMH-induced animals compared with the control animals. The antioxidant enzyme levels in experimental animals were restored to near normal after supplementation with β-sitosterol at 20 mg/kg of body weight. The levels of nonenzymatic antioxidants in colon and liver of control and DMH-induced animals are given in Figure 1. The levels of hepatic nonenzymatic antioxidants (GSH, vitamin C, and vitamin E) and colonic GSH were significantly decreased, whereas activities of the colonic nonenzymatic antioxidants (vitamins C and E) were found to be increased in DMH-induced experimental animals. Treatment with β-sitosterol at a dose of 20 mg/kg of body weight significantly restored the levels of nonenzymatic antioxidants to near normal.
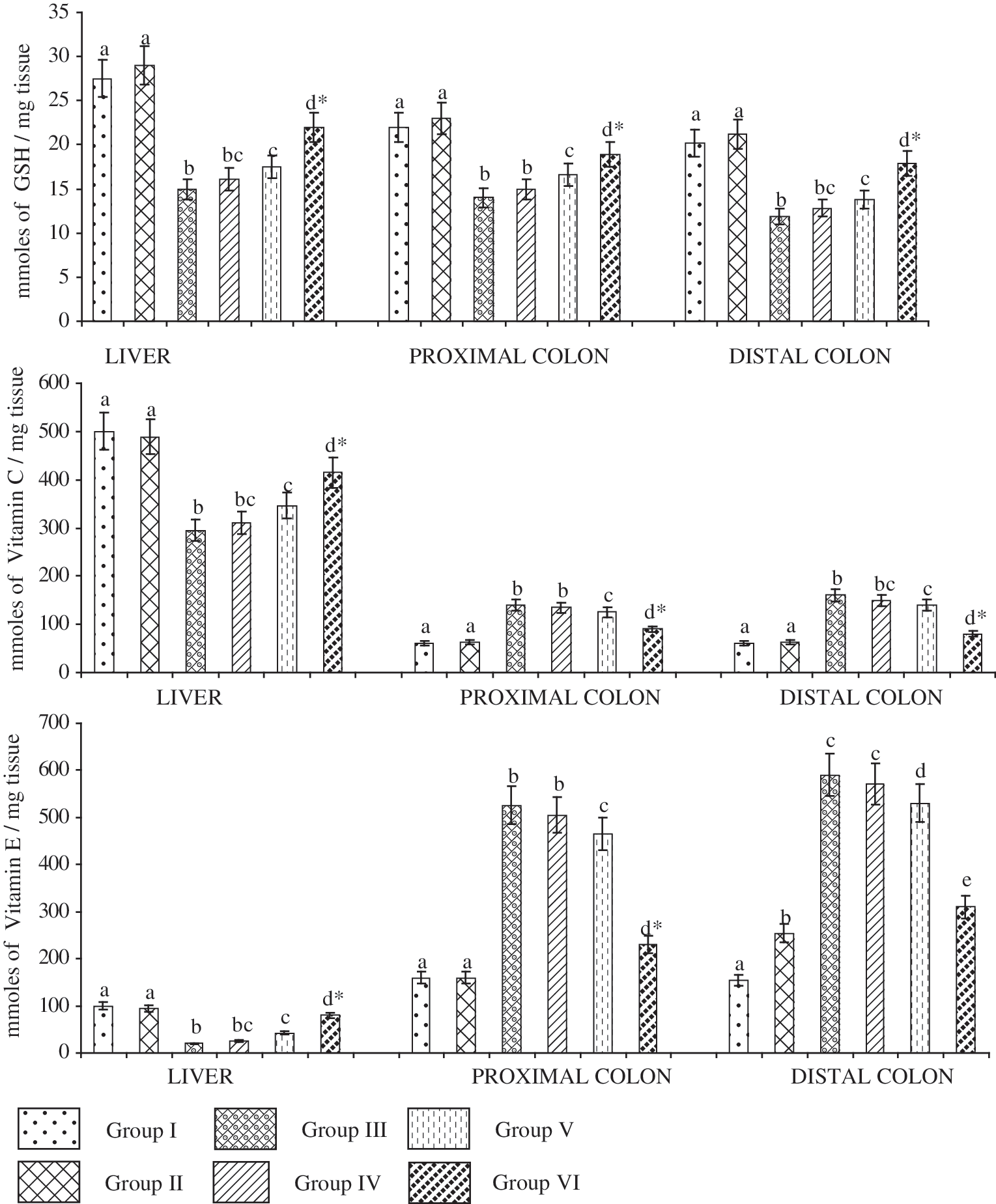
Effect of β-sitosterol on (top) GSH, (middle) vitamin C, and (bottom) vitamin E activity in the liver and colon of control and experimental rats. Data are presented as means±SD of 10 rats in each group. abcdValues not sharing a common superscript letter are significantly different (P<.05). *Values are significantly different (P<.01) as compared to 1,2-dimethylhydrazine alone–treated group (Group III).
Data are mean±SD values of 10 rats in each group. SOD activity is given in amount of enzyme required for 50% inhibition of nitro blue tetrazolium reduction/min/mg of protein. CAT activity is given in μmol of H2O2 utilized/min/mg of protein. GR activity is given in μmol of NADPH oxidized/min/mg of protein. GPx activity is given in μmol of reduced glutathione utilized/min/mg of protein. Glutathione S-transferase (GST) activity is given in μmol of 1-chloro-2,4-dinitrobenzene–reduced glutathione conjugate formed/min/mg of protein.
P<.05, values not sharing a common superscript letter are significantly different.
P<.01, values significantly different compared with the 1,2-dimethylhydrazine alone–treated group (Group III).
Figure 2 shows the histopathological alterations in normal and DMH-induced animals. Treatment with 20 mg/kg of body weight β-sitosterol was able to reverse damage in the DMH-induced animals to near normal. The animals treated with DMH alone showed well-differentiated signs of dysplasia.
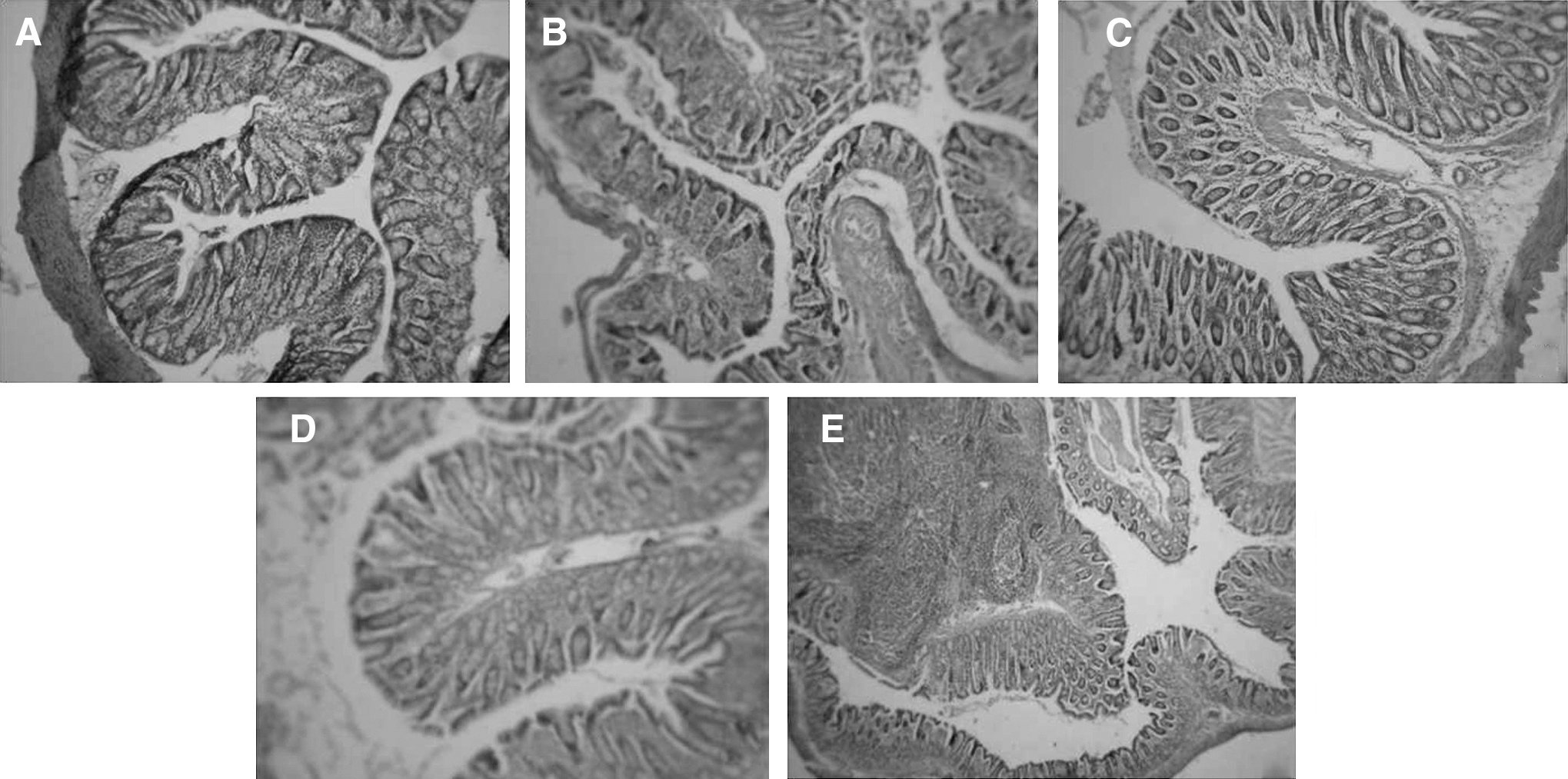
Histopathological alterations in experimental and control animals treated with β-sitosterol. For all panels, magnification ×20.
Discussion
Phytosterols cannot be synthesized by humans, and hence dietary consumption is the only source of tissue and plasma phytosterols. Because of decreased absorption from the intestine, low plasma levels are observed in healthy individuals. 28 Generally it has been suggested that in humans and other mammals only about 5% of the ingested plant sterols are absorbed. 28,29 The absorption rate of β-sitosterol was found to be 4.2% in healthy individuals. 30 The most commonly occurring phytosterols in the human diet are β-sitosterol, campesterol, and stigmasterol, which account for approximately 65%, 30%, and 3% of dietary phytosterols, respectively. 31 Because of the lower absorption rate in the intestine only 4.2% of the ingested β-sitosterol is absorbed and found in circulation. 30,32,33 The doses of 5, 10, and 20 mg/kg of body weight β-sitosterol used in this present study were selected based on our preliminary dose fixation study. Human equivalent dose is the dose in humans expected to provide the same degree of effect as that observed in animals at a given dose. 34 The maximum dose (20 mg/kg of body weight) selected for the study in rats is equivalent to 3.24 mg/kg of body weight in a healthy adult. However, this human equivalent dose of 3.24 mg/kg lies in the physiological range for humans. β-Sitosterol can be considered as an effective supplement for human well-being to prevent colon carcinogenesis. Almost 90% of plants contain β-sitosterol, and so consumption of green vegetables would be sufficient to obtain the effective concentration. Table 6 summarizes the percentage of β-sitosterol in plant foods.
Source:
Oxidative stress occurs as a result of an imbalance between free radical generation and the antioxidant levels. ROS are part of normal metabolic processes, 36 and lowered antioxidant status is associated with increased risk of cancer. Oxidative injury in cells may induce gene mutation and promote carcinogenesis. 37 On the other hand, cancer cells are under high levels of oxidative stress compared with normal cells. 38 Increased production of free radicals in cells leads to alteration of cellular components and causes damage to cells, eventually leading to death. It is known that the levels of free radical molecules are controlled by various cellular defense mechanisms consisting of enzymatic (CAT, SOD, GPx, GR, and GST) and nonenzymatic (vitamins E and C and GSH) components. 3 The cell damage and the subsequent lack of cellular repair processes due to the constant oxidative damage have been associated with carcinogenesis. 39,40
In our present study β-sitosterol supplementation in experimental and control animals did not show any toxic symptoms. Rats induced with DMH showed a decreased gain in body weight compared with control rats. We observed a steady gain in the body weight of animals treated with DMH and supplemented with β-sitosterol. All the rats consumed food and water ad libitum, and there was no difference in the food and water intake among the experimental and control animals (data not shown). All the experimental animals were alive at the end of the experimental period.
DMH, a potent carcinogen with selectivity for the colon, can produce colon cancer in experimental rats. DMH is metabolized to a methyl free radical and generates hydroxyl radicals or hydrogen peroxide responsible for lipid peroxidation and DNA damage. 41 The magnitude of this damage (called oxidative stress or oxidative damage) depends not only on ROS levels but also on the defense mechanisms against them mediated by various cellular antioxidants. Disruption of this delicate oxidant/antioxidant balance seems to play a causative role in carcinogenesis. High levels of oxidative stress result in peroxidation of membrane lipids with the generation of peroxides that can decompose to multiple mutagenic aldehyde products. An enhanced lipid peroxidation is considered to be mutagenic and carcinogenic. 42 In our present study, lipid peroxidation as evidenced by the increased levels of TBARS, LOOHs, and DCs was significantly enhanced in the liver of DMH-treated rats, which could be attributed to DMH-induced oxidative stress and production of reactive oxygen metabolites. On the other hand, the levels of TBARS, LOOHs, and DCs were significantly decreased in the colonic tissues of DMH-treated rats. One reason for the increased lipid peroxidation in liver and the decrease in the colon of DMH-induced animals may be due to poor enzymatic and nonenzymatic antioxidant defense systems. This may also be due to excessive generation of lipid peroxidation products in tumor tissue and subsequent release of these products into the circulation. An increase of lipid peroxidation level in proliferating cells leads to an increase in the circulating lipid peroxides in DMH-induced animals. 43 Lowered levels of lipid peroxidation may be due to an inverse relationship between the levels of cellular lipid peroxidation and rates of cell proliferation and/or the extent of differentiation. 44 Thus, it appears likely that the levels of lipid peroxidation are inversely related to the rate of cellular growth irrespective of whether the cells are malignant or not. It is possible that although the changes in lipid peroxidation status may reflect cell growth rate rather than malignancy in itself, important changes will nevertheless occur at some earlier stage during the progression of normal cells to malignancy. β-Sitosterol scavenges the lipid peroxidation products in DMH-induced animals by virtue of its antioxidant ability. 16
Some chemopreventive drugs are thought to exhibit their anticancer ability mainly because of their antioxidant capacity by scavenging the free radicals produced in cancer. 9 Antioxidant enzymes in cancer cells are found to vary in comparison with the levels in normal cells. These antioxidant enzymes and scavengers of ROS protect the cellular components from free radical induced damage. The enzymatic antioxidants SOD, CAT, GR, GPx, and GST play a vital role in the defense against toxic free radicals produced in cancer cells. DMH-induced animals exhibit decreased levels of these antioxidant enzymes, and treatment with antioxidant molecules enhances the activities of the enzymes. 45 A decline in these enzymes may facilitate the initiation of oxidative processes, which would lead to the elevation of ROS levels and consequently may account for increases in the levels of oxidized DNA bases, which lead to mutagenesis and carcinogenesis. The results of the current study revealed decreased activities of enzymatic antioxidants in liver and colonic tissue of DMH-induced animals. The reduced activities of antioxidant enzymes could be due to increased production of free radicals that exceeds the scavenging capacity of the antioxidant systems in cancer. Therefore, activities of antioxidant enzymes are needed at a higher level to protect cells against toxic free radical damage. β-Sitosterol supplementation at 20 mg/kg of body weight significantly increased the activities of antioxidants in liver and colonic tissues of DMH-induced animals.
GSH is a major nonenzymatic antioxidant and plays a second line of defense against free radical damage. In our study, GSH levels in the colonic and liver tissues were significantly reduced in DMH-induced animals. However, supplementation of β-sitosterol at 20 mg/kg of body weight significantly increased the concentration of hepatic and colonic GSH levels. The nonenzymatic antioxidant levels were elevated in DMH-induced animals compared with the normal tissues in our study, which correlates with previous studies. 46,47 Superoxide anions generated in the tumor tissues oxidize ascorbic acid to dehydroascorbate, the transportable form of vitamin C, and this is trapped in tumor tissues by reduction to the nontransportable forms. Increased vitamin E levels in cancer tissues may be due to decreased polyunsaturated fatty acids, which might contribute to the low rate of lipid peroxidation. 48 Upon β-sitosterol supplementation, the levels of vitamin C were decreased, which could be attributed to the decreased uptake of vitamin C and vitamin E and may also be correlated with decreased resistance of transformed cells to lipid peroxidation.
The DMH-induced animals showed irregular glands with stratified epithelium and hyperchromatic nuclei, which are reported to be present in cancer tissues. The animals treated with β-sitosterol exhibited a pattern of normal cells with very few dysplastic changes. Hyperplasia was assessed by counting the number of cells per crypt column (crypt height). The number of cells in the crypt column was decreased by β-sitosterol supplementation. In addition, the decrease in crypt height might be correlated with a significant decrease in hyperplastic lesions. 49 Treatment with 20 mg/kg of body weight β-sitosterol in DMH-induced animals restored the colonic epithelial cells to near normal, indicating the anticancer potential towards experimental colon carcinogenesis. In our previous study, we have reported that β-sitosterol suppressed the expression of β-catenin and proliferating cell nuclear antigen in COLO 320 DM cells and reduced the incidence of aberrant crypt foci in DMH-induced animals because of its antioxidant property. 16 Chemopreventive ability of a drug is attributed to its ability to scavenge free radical, which play an important role in pathogenesis. We have previously reported the anticancer potential of β-sitosterol may be due to its antioxidant and antiproliferative ability in DMH-induced animals. 39 Results of our present study support the anticancer potential of β-sitosterol. The cancer chemopreventive potential of β-sitosterol in DMH-induced animals may be due to its antiproliferative and ability to suppress the β-catenin and proliferating cell nuclear antigen mutation in DMH-induced experimental carcinogenesis.
Conclusion
In conclusion, the results of the present study suggest that β-sitosterol exhibits chemopreventive potential in DMH-induced experimental colonic preneoplastic progression in rats by its radical scavenging ability. Hence, the study determined that the effective dose for long-term studies to be 20 mg/kg of body weight in DMH-induced experimental colon carcinogenesis.
Footnotes
Acknowledgment
This work was supported by the Indian Council of Medical Research, New Delhi, India (Senior Research Fellowship to A.A.B., Project Number 2005-00930).
Author Disclosure Statement
No competing financial interests exist.