
Abstract
Helicobacter pylori is a major human pathogen that plays central roles in chronic gastritis and gastric cancer. Recently, we reported that auraptene suppressed H. pylori adhesion via expression of CD74, which has been identified as a new receptor for H. pylori urease. In this study, we attempted to clarify the effects of oral feeding of auraptene on H. pylori infection and resultant inflammatory responses in C57BL/6 mice and found that it remarkably attenuated H. pylori colonization and gastritis. Biochemical analyses revealed that auraptene inhibited H. pylori–induced expression and/or production of CD74, macrophage migration inhibitory factor, interleukin-1β, and tumor necrosis factor-α in gastric mucosa, together with serum macrophage inhibitory protein-2. It is notable that treatment with this coumarin during the pretreatment period was more effective than that during posttreatment. Our results suggest that auraptene is a promising phytochemical for reducing the risk of H. pylori–induced gastritis and carcinogenesis.
Introduction
H
Adhesion of H. pylori to gastric epithelial cells is recognized as one of the essential steps in developing gastritis, which leads to promotion of a definitive virulence factor, cytotoxin-associated antigen A, through type IV secretion systems. 6 However, the adhesion mechanisms of H. pylori to gastric epithelial cells are not fully understood. Recently, Beswick et al. 7,8 suggested that the urease B subunit binds to CD74 on gastric epithelial cells and induces nuclear factor-κB activation, thereby stimulating interleukin (IL)-8 production. On the other hand, other reports have suggested that H. pylori inoculates cytotoxin-associated antigen A into gastric epithelial cells for inducing phosphorylation of mitogen-activated protein kinase kinase, Src, and SHP-2, 9,10 thereby promoting the production of IL-8, IL-1β, and tumor necrosis factor-α (TNF-α). 11 These cytokines play several crucial roles in H. pylori–associated gastritis through recruitment, activation, and infiltration of neutrophils to sites of infection, as well as chronic inflammation and gastric injury. 12 –14
Several food extracts and components have been shown to lower the risk of damage derived from H. pylori infection. Green tea catechins, such as (–)-epigallocatechin-3-gallate, strongly inhibited H. pylori urease activity in vitro and suppressed H. pylori–induced gastritis in Mongolian gerbils. 15 Also, broccoli sprouts rich in sulforaphane, which has an antibacterial effect on H. pylori by unknown mechanisms and also exhibits a chemoprotective effect, significantly decreased colonization and attenuated gastritis in H. pylori–infected mice and humans. 16,17 However, these phytochemicals have yet to be proven adequately effective for clinical use; because of their broad range of biological activities, more specific molecular targeting is indispensable to elucidate their potential efficacy. In this context, we reported that citrus auraptene markedly inhibits extracellular signal-regulated kinase 1/2 activation and thereby suppresses H. pylori adhesion to gastric cells and inflammatory responses in NCI-N87 human gastric carcinoma cells. 18 Auraptene is present in various citrus fruits, such as hassaku (Citrus hassaku) and grapefruit (Citrus paradici), and can be prepared by an established method in large quantities. 19 –21 In addition, this coumarin was shown to have anti–H. pylori urease activity in the glandular stomach of Mongolian gerbils, although mechanistic findings were not provided. 22 In the present study, we evaluated the effects of auraptene on adhesion and resultant cytokine production in H. pylori–infected C57BL/6 mice.
Materials and Methods
Chemicals
Oligonucleotide primers were synthesized by Sigma Genosys (Hokkaido, Japan). Auraptene was isolated using a method previously described. 20,23 All other chemicals were purchased from Wako Pure Chemicals (Osaka, Japan), unless specified otherwise.
Animals
Six-week-old female C57BL/6 mice were purchased from Japan SLC, Inc. (Shizuoka, Japan). The animals were housed in cages under a temperature (23±2°C)- and humidity (55±5%)-controlled environment, with a 12-h light and dark cycle, and were provided Oriental MF® sterilized with γ-rays from Oriental Yeast Co., Ltd. (Tokyo, Japan) and water ad libitum throughout the experiment. The animals were handled according to the guidelines of the Committee for Ethics in Animal Experimentation of the University of Shizuoka.
Bacterial culture
H. pylori NCTC 11637 was purchased from the American Type Culture Collection and grown on Columbia HP agar plates (Becton Dickinson, San Diego, CA, USA) at 37°C under microaerophilic conditions. After 72 h, bacterial cells were transferred to Brucella broth containing 10% fetal bovine serum, centrifuged at 4000 g for 5 min, and then resuspended in phosphate-buffered saline. Bacterial density was determined by measuring absorbance at 595 nm using a microplate reader (Thermo Fisher Scientific, Waltham, MA, USA).
Animal treatment
Following a prefeeding period of 1 week, 70 mice were randomly divided into the following eight groups (Fig. 1): untreated (Group 1), H. pylori infection alone (Group 2), 500 ppm of auraptene alone for 9 weeks (Group 3), 100 ppm (Group 4) or 500 ppm (Group 5) of auraptene before infection, 500 ppm of auraptene alone for 7 weeks (Group 6), and 100 ppm (Group 7) or 500 ppm (Group 8) of auraptene after infection, with five to 15 mice in each group. H. pylori infection was performed as previously reported by Hirayama et al. 24,25 In brief, 7-week-old mice were inoculated orally with an H. pylori suspension that contained approximately 5×107 colony-forming units (CFU)/0.2 mL using a feeding needle. Mice in groups were fed their respective diets for the entire designated experimental period. In week 8, the mice were fasted for 24 h and then euthanized, after which the stomachs and whole blood were immediately removed. After the stomachs were excised, they were opened along the greater curvature, and the intragastric contents were gently removed. To determine CFU, each stomach was cut finely and homogenized in 7 mL of sterilized saline, followed by serial dilutions with the same saline, after which 0.1-mL aliquots of the diluted homogenates were inoculated onto Helicobacter agar plates (Nissui Pharmaceutical Co., Ltd., Tokyo). The plates were incubated at 37°C under microaerophilic conditions for 7 days, and then the colonies were counted to detect H. pylori CFU. The degree of gastric mucosal injury was determined by a pathologist (F.T.) in a blind fashion and scored using the following criteria: 0, normal; 1, edematous; 2, erosion; 3, multiple erosions; and 4, hemorrhage erosions and/or ulcers larger than 1 mm in diameter.
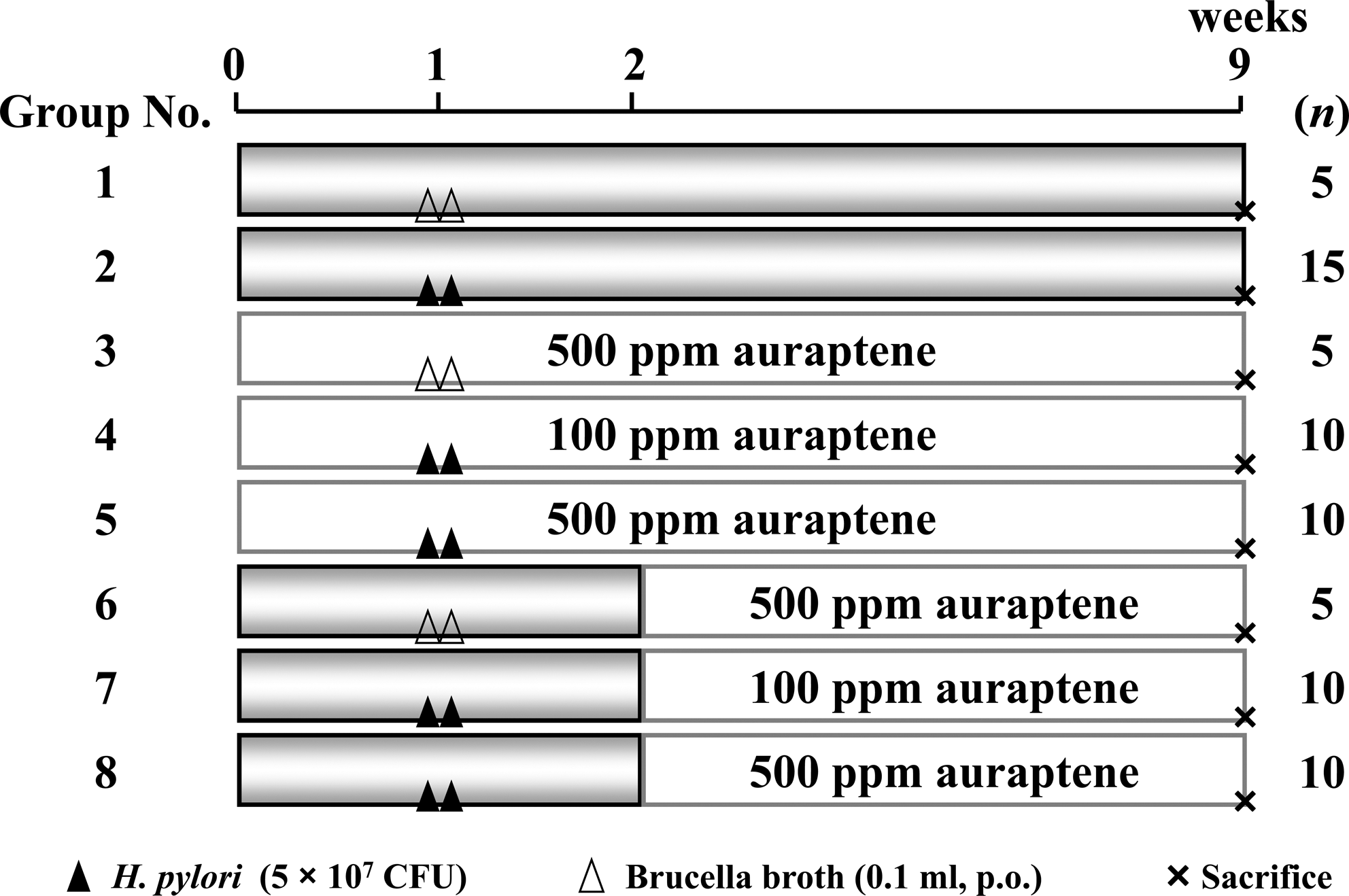
Experimental protocol for H. pylori infection model. Mice in Groups 2, 4, 5, 7, and 8 were orally administered H. pylori (5×107 colony-forming units [CFU]) twice. Mice in Groups 4 and 7 (100 ppm) and 3, 5, 6, and 8 (500A ppm) were given auraptene in the diet throughout the experimental period.
Reverse transcription–polymerase chain reaction
Total RNA was extracted from each stomach using TRIzol® reagent (Qiagen, Hilden, Germany), according to the manufacturer's instructions. The cDNA was synthesized using 1 μg of total RNA and an RNA polymerase chain reaction kit (AMV, TaKaRa Bio, Shiga, Japan). Polymerase chain reaction amplifications were performed with a thermal cycler (model PTC-100TM, MJ Research, Waltham) using the following primers and conditions: CD74, forward (5′-gTgTCTgTTTCATCgTCCCAg-3′) and reverse (5′-AAggCAgCAAATgTgTCCAgC-3′) (product size, 205 bp) for 30 cycles, with 30 sec of denaturation at 94°C, 30 sec of annealing at 65°C, and 30 sec of primer extension at 72°C; macrophage migration inhibitory factor (MIF), forward (5′-CCATgCCTATgTTCATCgTg-3′) and reverse (5′-gAACAgCggTgCAggTAAgTg-3′) (product size, 381 bp) for 35 cycles, with 60 sec of denaturation at 94°C, 60 sec of annealing at 60°C, and 60 sec of primer extension at 72°C; IL-1β, forward (5′-ATggCAACTgTTCCTgAACTCAACT-3′) and reverse (5′-AAggCAgCAAATgTgTCCAgC-3′) (product size, 586 bp) for 30 cycles, with 30 sec of denaturation at 94°C, 30 sec of annealing at 60°C, and 30 sec of primer extension at 72°C; TNF-α, forward (5′-ggCAggTCTACTTTggAgTCATTgC-3′) and reverse (5′-ACATTCgAGGCTCCAgTgAATTCgg-3′) (product size, 309 bp) for 35 cycles, with 45 sec of denaturation at 94°C, 45 sec of annealing at 65°C, and 60 sec of primer extension at 72°C; and glyceraldehyde 3-phosphate dehydrogenase, forward (5′-gTgAAggTCggAgTCAACg-3′) and reverse (5′-ggTgAAgACgCCAgTggACTC-3′) (product size, 300 bp) for 30 cycles, with 30 sec of denaturation at 94°C, 30 sec of annealing at 55°C, and 30 sec of primer extension at 72°C. Amplified cDNA was subjected to electrophoresis on 3% agarose gels and stained with SYBR® Gold (Molecular Probes, Carlsbad, CA, USA). Image analysis was performed using the NIH Image software program. Glyceraldehyde 3-phosphate dehydrogenase was used as the internal control.
Enzyme-linked immunosorbent assay
Collected serum samples without dilution were used for enzyme-linked immunosorbent assay, with measurements of mouse macrophage inflammatory protein-2 (MIP-2) (R&D Systems, Minneapolis, MN, USA) performed according to the manufacturer's instructions.
Statistical analysis
Each result is expressed as the mean±SD or SEM. One-way analysis of variance was performed, followed by Tukey's multiple comparison test to determine whether the differences among the groups were significant. Differences were considered significant at P<.05.
Results
Auraptene blocked H. pylori colonization and resultant gastric mucosal injuries
Oral feeding of auraptene (Groups 3–8) did not affect food intake, water intake, or body weight compared with the untreated control (Group 1) (data not shown). Eight weeks following infection with H. pylori, the level of H. pylori colonization in the stomachs of mice was decreased by 45–50% in Groups 4 and 5 (auraptene during pretreatment) compared with Group 2 (infection alone), whereas posttreatment treatments (Groups 7 and 8) were not effective (Fig. 2A). The gastric mucosal injury score in Group 2 (score=2.3) was significantly higher than that in Group 1 (control) (Fig. 2B). It is notable that the injury scores of the mice in Groups 5 and 6 (500 ppm of auraptene pre- and post-treatment, respectively) were reduced by 74–82%, whereas that in Group 7 (100 ppm posttreatment) was not reduced (Fig. 2C). Furthermore, ulcers (injury score=4.0) were observed in Group 2 (H. pylori infection alone) and Groups 7 and 8 (auraptene posttreatment), but not in Groups 4 and 5 (auraptene pretreatment). In addition, Groups 3 and 6 (auraptene without H. pylori) did not show H. pylori colonization or gastric mucosal injury.
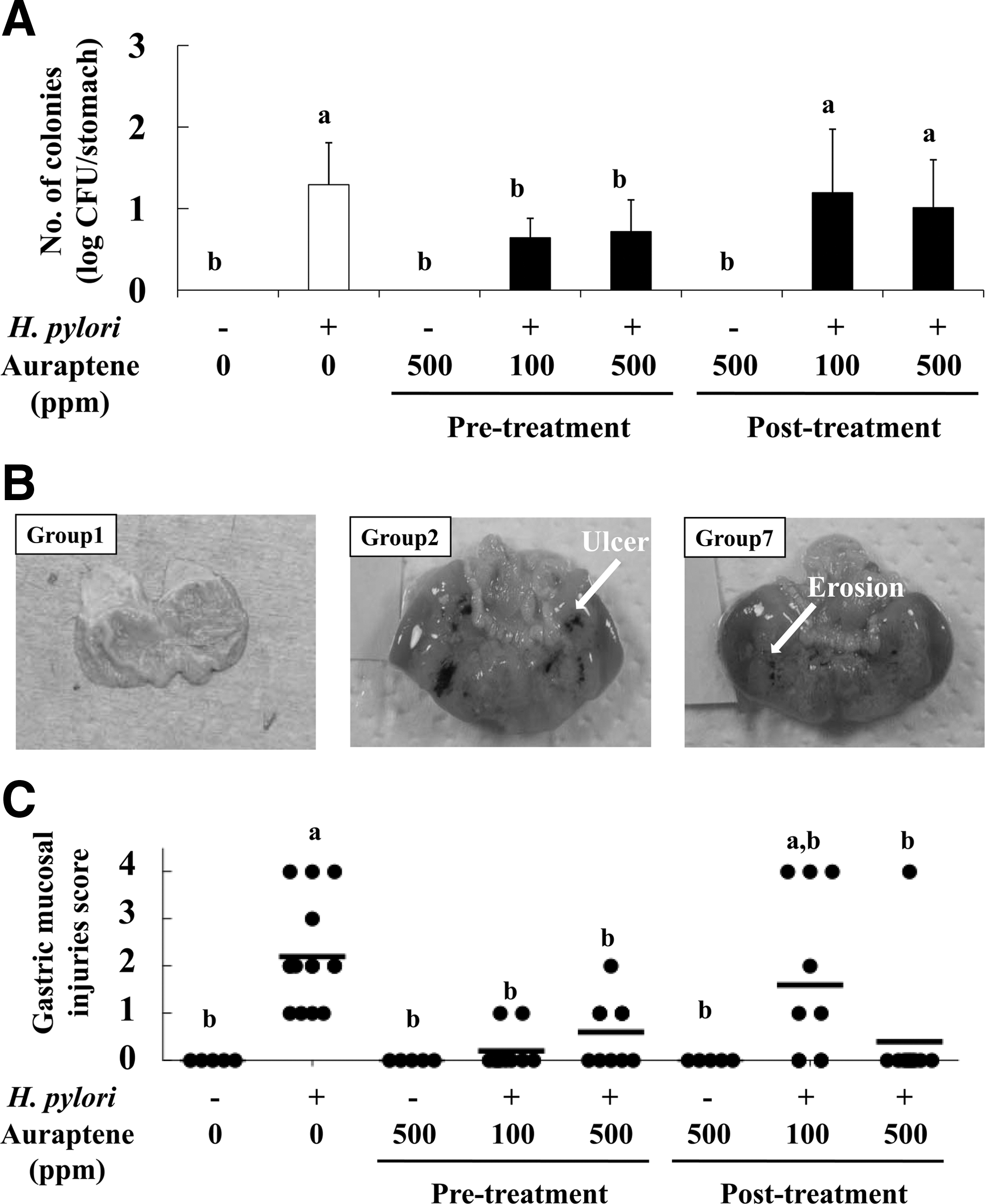
Effects of auraptene on
Auraptene attenuated expressions of CD74 and pro-inflammatory cytokines
We also investigated whether auraptene suppresses H. pylori adhesion-dependent production of pro-inflammatory cytokines. As noted above, H. pylori urease binding to CD74 results in induction and production of numerous pro-inflammatory cytokines. As shown in Figure 3A and B, the mRNA expressions of CD74 and its ligand MIF were significantly increased by 2.6- and 3.5-fold, respectively, in H. pylori–infected mice (Group 2). However treatments with auraptene during the pre- and posttreatment periods (Groups 4, 5, 7, and 8) reversed those levels to those of the controls. Similarly, H. pylori infection increased IL-1β and TNF-α expressions by 9.6- and 9.2-fold, respectively, whereas those were suppressed in Groups 4, 5, 7, and 8 (auraptene pre- and posttreatment) by 62–85% and 66–93%, respectively (Fig. 3C and D).
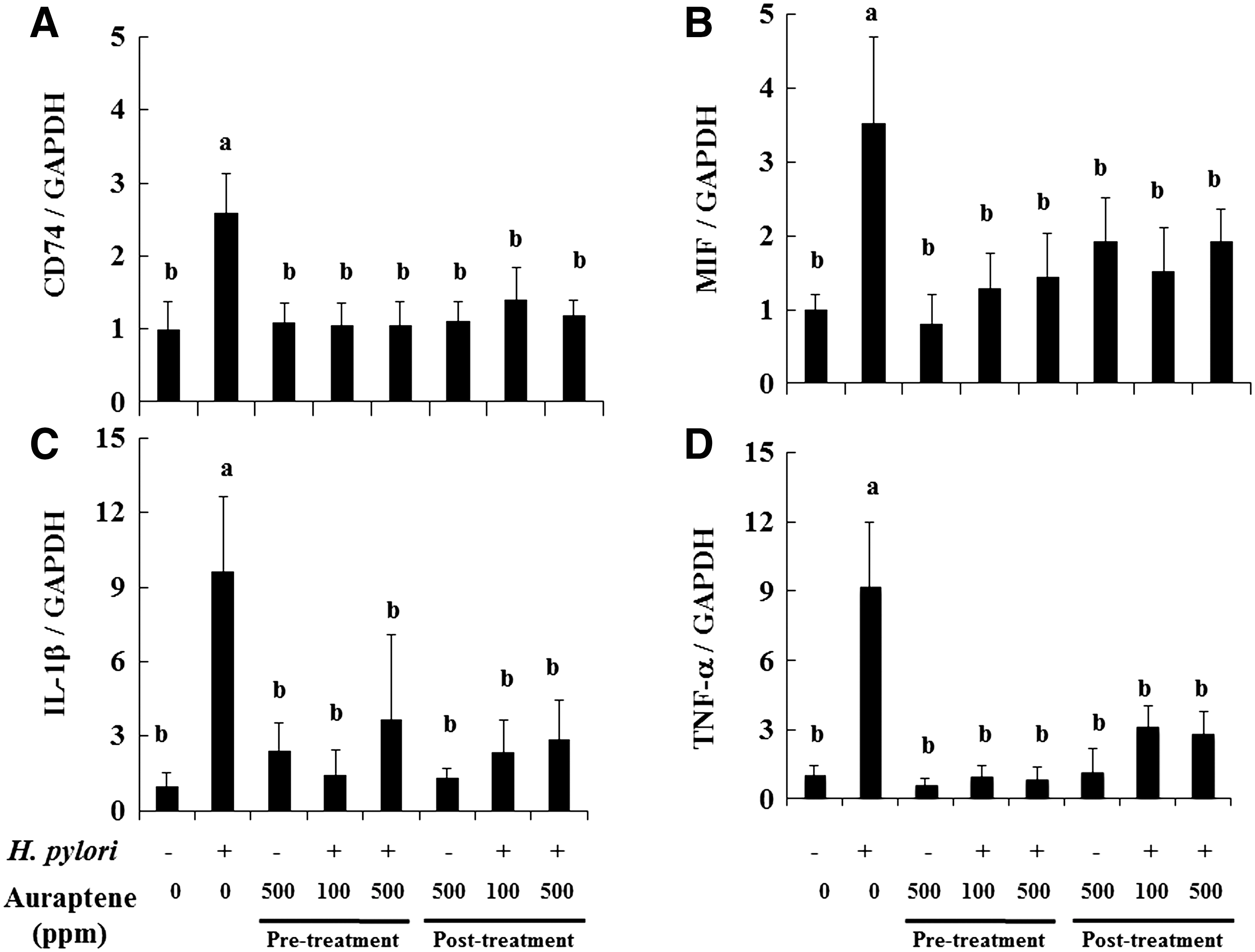
Effects of auraptene on expressions of
Auraptene decreased serum MIP-2 levels
Finally, we assessed the effects of auraptene treatment on production of the chemokine MIP-2. Mouse MIP-2, also known as CXC chemokine CXCL2, has been suggested to be a functional homolog of human IL-8 in mice. 26,27 The serum MIP-2 levels in Group 2 (H. pylori infection) were increased by 1.4-fold compared with Group 1. Although pretreatment with auraptene at 100 and 500 ppm (Groups 4 and 5, respectively) significantly inhibited its production by 19–30%, posttreatment suppressed the level of MIP-2 only in Group 8 (500 ppm of auraptene) by 19% (Fig. 4).
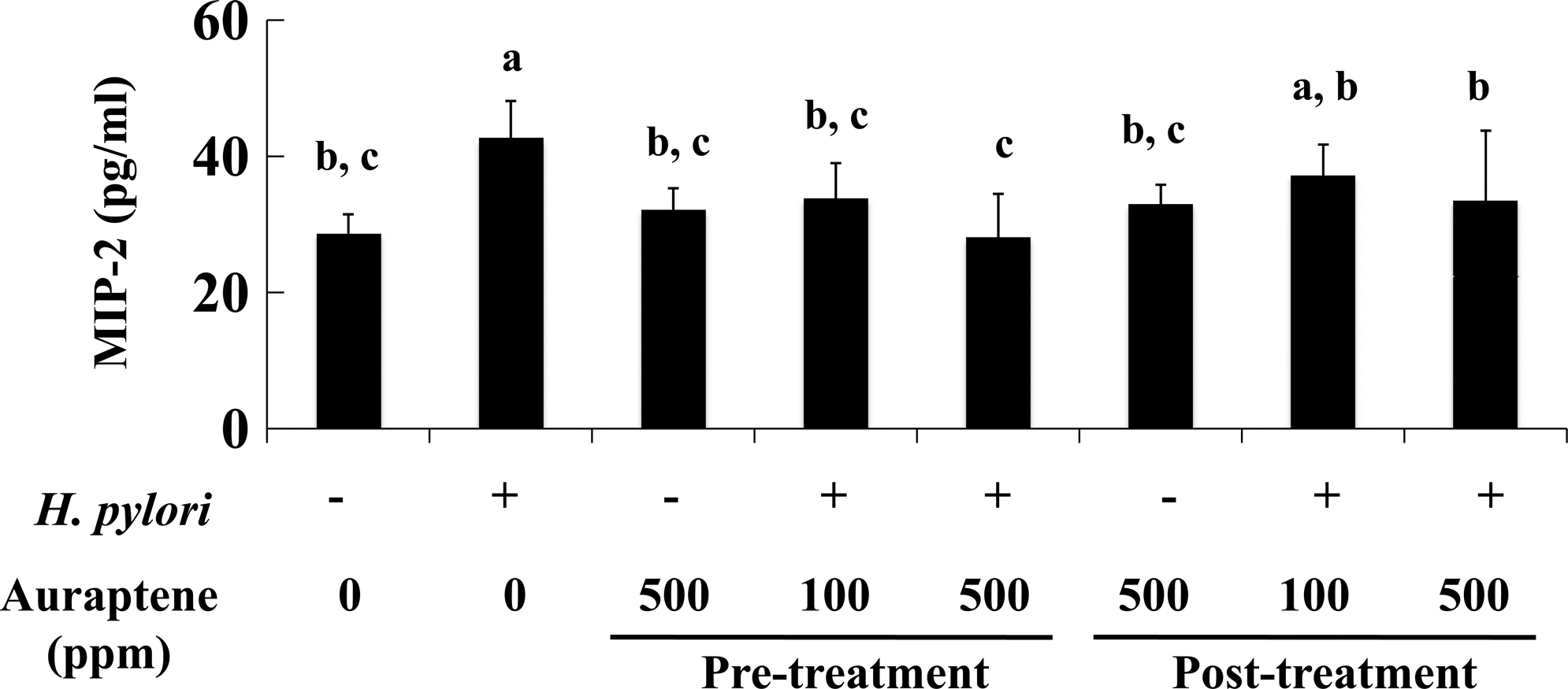
Effects of auraptene on serum macrophage inhibitory protein-2 (MIP-2) level. Enzyme-linked immunosorbent assay was used to determine MIP-2 levels, as described in Materials and Methods. Data are mean±SD values. abcBars not sharing a letter differ significantly. P<.05 by Tukey's multiple comparison test.
Discussion
Takeda et al. 22 reported that auraptene treatment during postinfection (100 and 500 ppm) was not effective for H. pylori colonization in Mongolian gerbils, which is consistent with the present findings. However, we found that it markedly inhibited H. pylori colonization when given during the pretreatment period (Fig. 2A). Previous reports have suggested that H. pylori eradication at an early stage might increase the success rate of reducing colonization and be effective for preventing gastric carcinogenesis. 28 Collectively, auraptene may be more effective for prevention of H. pylori–induced gastritis than for treatment of the disease. It is important to note that during the posttreatment period auraptene did not reduce the colonization of H. pylori (Fig. 2A), whereas it significantly decreased gastric injury (Fig. 2C) and attenuated the expression and production of pro-inflammatory mediators (Figs. 3 and 4). These conflicting results suggest that auraptene does not affect the growth of H. pylori and is lethal toward that bacterium during the posttreatment period, whereas it is capable of suppressing the biological activation of H. pylori–infected gastric cells. In contrast, auraptene given during the pretreatment period may have growth inhibitory effects and thereby down-regulate pro-inflammatory mediator production. In a recent report, we also noted that CD74 may partially contribute to H. pylori adhesion as well as pro-inflammatory events. 18 It is interesting that Wong et al. 29 found that CD74 expression was up-regulated immediately after infection. In the present study, CD74 mRNA expression was dramatically decreased by auraptene (Fig. 3A), whereas H. pylori infection was inhibited only in the pretreatment groups (Fig. 2B). Together, these results indicate that early-stage treatment targeting CD74 may be a reasonable and promising strategy for suppressing H. pylori colonization in mice.
Recently, we reported that auraptene suppressed IL-8 production in gastric cancer cells via down-regulation of extracellular signal-regulated kinase 1/2. 18 IL-8 plays a crucial role in H. pylori–associated gastritis through recruitment, activation, and infiltration of neutrophils to the sites of infection. The release of IL-8 from gastric epithelium, which is induced by TNF-α and IL-1β and directly caused by H. pylori strains, induces the expression of adhesion molecules, such as CD11b/CD18, and resultant production of reactive oxygen species. 30,31 If the various defense mechanisms fail and chronic infection becomes established, continuous up-regulation of IL-8 and activation of neutrophils could lead to mucosal damage and increased free radical formation. 32 Furthermore, MIF, the ligand of CD74, acts as a pro-inflammatory mediator, including IL-1, TNF-α, and IL-8. 33 –35 In the present study, we found that the expressions of CD74 and MIF mRNA were markedly up-regulated with H. pylori infection (Fig. 3A and B), which in turn induced the development of gastric inflammation. Once released, MIF functions as both a paracrine and autocrine activator of its receptor CD74 and then binds and activates CD74 to induce the cascade of gastric inflammation, resulting in up-regulation of IL-1β and TNF-α and production of MIP-2 (Figs. 3C and D, and 4). A previous report suggested that MIF knockout mice infected with H. pylori express not only a normal level of CD74, but are also protected against gastric inflammation. 29 Therefore, CD74 suppression by auraptene may, at least in part, contribute to down-regulation of MIF, IL-1β, and TNF-α expression, as well as MIP-2 production (Figs. 3B–D and 4).
In conclusion, our results showed that auraptene notably suppresses H. pylori colonization and resultant gastritis by reducing inflammatory cytokine expression and chemokine production. This is the first known report to describe a food phytochemical with potential for regulating CD74 expression. Although no side effects from auraptene were noted in any mice in this study, toxicity assessment in a long-term experiment is necessary.
Footnotes
Acknowledgments
The authors thank Professor Kayoko Shimoi, Dr. Hiroyuki Sakakibara, and Ms. Asako Matsui, Graduate School of Nutritional and Environmental Sciences, Institute for Environmental Sciences, University of Shizuoka, for their technical support with the experiments. This study was supported in part by a Grant-in-Aid for Scientific Research (C) (to A.M.) from the Japan Society for the Promotion of Science, by a grant from the Council for Advancement of Fruit Tree Science (to A.M.), and a Grant-in-Aid for Research Fellowships of the Japan Society for the Promotion of Science for Young Scientists (to H.S.).
Author Disclosure Statement
All authors declare no competing financial interests.