
Abstract
The aim of this study was to investigate whether Dioscorea batatas (DB) extract attenuates high-fat diet (HFD)-induced insulin resistance in the visceral adipose tissues of mice, and by what mechanism(s). Mice were fed a HFD for 4 weeks to induce the early development of insulin resistance. The DB extract was administered to mice fed a HFD by oral gavage at a dose of 100 mg/kg body weight daily for 7 weeks. Biochemical parameters in blood were measured using enzymatic kits, and the expression levels of glucose transporter 4 (GLUT4), phosphorylated (p-)S6K1, phosphorylated v-akt murine thymoma viral oncogene homolog (p-AKT), and phosphorylated extracellular regulated kinase (p-ERK) in epididymal fat tissue were determined by western blot analyses. The DB extract effectively reversed the HFD-induced elevations in plasma glucose and insulin levels, and the homeostasis model assessment for insulin resistance and oral glucose tolerance test values. The level of p-AKT protein was up-regulated, whereas the levels of p-ERK and p-S6K1 proteins were down-regulated in the adipose tissues of DB mice compared with HFD mice. Furthermore, the DB extract significantly reversed the HFD-induced decrease in the plasma membrane GLUT4 level in the adipose tissue of mice. The DB extract improved glucose metabolism in HFD-fed mice through the up-regulation of plasma membrane GLUT4 content in the visceral adipose tissue. Activation of the insulin signaling cascade leading to GLUT4 translocation was the mechanism underlying the beneficial effects of the DB extract on early-stage obesity-induced insulin resistance.
Introduction
O
Besides insulin, oral hypoglycemic drugs that are currently used to treat type 2 diabetes include sulphonylurea derivatives, biguanides, thiazolidinediones, and α-glucosidase inhibitors. 11 These agents have undesirable side effects, such as hypoglycemia, 12,13 lactic acidosis, 12,14 heart failure, 15 and gastrointestinal disorders. 11 Herbal medicines are gaining more importance in the treatment of insulin resistance, as they are free from side effects and less expensive than synthetic hypoglycemic agents. 16,17
Yam (Dioscorea batatas [DB]) belongs to the Dioscoreaceae family and is found throughout East Asia, including China, Japan, Taiwan, and Korea. The rhizome of DB is used as a traditional medicine for the treatment of asthma, abscesses, chronic diarrhea, and ulcers. 18,19 DB exhibits several useful biological features, such as anti-inflammatory activity in lipopolysaccharide-stimulated murine macrophage cells, 20 as well as antioxidative and cytoprotective properties in primary mouse splenocytes. 21 In a previous study, an ethanol extract of Dioscorea opposite (Dioscoreaceae) was reported to be effective in reducing the plasma glucose level in type 2 diabetic rats. 22 However, the blood glucose-lowering effect of DB in mice fed a high-fat diet (HFD) has not been documented. The primary aim of this study was to investigate whether DB attenuates obesity-induced insulin resistance in the visceral adipose tissues of mice, and to explore the underlying mechanisms involved in this attenuation process.
Materials and Methods
Preparation of D. batatas extract
The rhizome of DB was purchased in June 2008 from an herbalist in Korea (Gyeong-buk Province). The specimen was authenticated by Professor Wan Kyunn Whang, at the College of Pharmacy, Chung-Ang University, Korea, where a voucher specimen has been deposited (No. DA-9802). The dried rhizome of DB was homogenized to a fine powder. The powdered DB rhizome (1 kg) was soaked in a water-ethanol (1:1, v/v) solution (1:5, plant weight/solvent volume) for 24 h, with occasional shaking. After filtration through Whatman no. 1 filter paper, the filtrate was evaporated under reduced pressure at 50°C and then spray dried. The dried DB extract yield was 53 g per kg of powdered material, and it was stored at room temperature in opaque bottles.
High-performance liquid chromatography analysis of allantoin
Pure allantoin standard was purchased from Sigma-Aldrich and was used as an external standard in the high-performance liquid chromatography (HPLC) analyses. The standard solution of allantoin was prepared by dissolving 50 mg of pure allantoin in 50 mL of ethanol (1 mg/mL). A sample solution of the DB extract was prepared by centrifugation at 13,000 g for 5 min, and the supernatant was used for further investigation. Allantoin was measured using an Agilent 1200 Series HPLC equipped with a photodiode array detector, an Inertsil NH2 column (4.6 mm×250 mm, 5 μm inner diameter), and an auto-sampler. The binary mobile phase consisted of water (Solvent A) and acetonitrile (Solvent B). The gradient elution program was set to 90% B (0–15 min), 90–70% B (15–20 min), 70% B (20–50 min), 70–90% B (50–50.1 min), and 90% B (50.1–55 min). The flow rate was set at 1 mL/min, and the absorbance of the effluent was monitored at 220 nm. The injection volume was 10 μL, and the peaks were identified by comparing the retention time with that of the standard solution.
Animals and experimental protocol
Twenty-four male C57BL/6 mice (5-week-old) were purchased from Central Lab. Animal (Seoul, Korea). The mice were housed in individual cages, maintained under standard conditions with a 12-h light-dark cycle, and had free access to food and water. All mice were fed a chow diet (CD) for a period of 1 week (adaptation period), after which they were randomly divided into three groups (eight mice per group). One group was fed the CD, and the other two groups were fed a HFD (55% of kcal from fat, 24% of kcal from carbohydrates, and 21% of kcal from protein; Research Diet). After 4 weeks, one of the groups maintained on the HFD was administered the DB extract suspended in 3% hydroxypropyl methylcellulose, whereas the other group was maintained on the HFD, and the control mice were maintained on the CD. The DB extract was administered daily for 7 weeks by oral gavage at a dose of 100 mg/kg of body weight.
During the treatment period, the body weights of the mice were monitored, and fasting blood specimens were collected from the tail vein every week. At the end of the experimental period, the animals were anesthetized with ether after a 12-hour fasting period. To analyze plasma insulin and glucose levels, freshly collected blood samples were centrifuged at 2000 g for 15 min at 4°C. Epididymal adipose tissues were removed and rinsed with phosphate-buffered saline. The plasma and epididymal fat pad samples were stored at −70°C until analysis. All mice were housed in the specific pathogen-free facility of Yonsei University, Seoul, Korea, and this study was approved by the Institutional Animal Care and Use Committee of Yonsei University.
Measurement of blood glucose and insulin levels
Glucose was measured in the blood obtained from the tail vein using a hand-held glucose meter (Accu-Chek Active; Roche Diagnostics). Plasma insulin concentration was determined using an ELISA kit (AKRIN-011T; Shibayagi). Plasma glucose levels were determined enzymatically using a commercial kit (Bio-Clinical System). The insulin to glucose ratio (IGR) value was calculated and analyzed as a surrogate index of insulin resistance.
Oral glucose tolerance test
Oral glucose tolerance tests (OGTTs) were conducted 1 week before the end of oral gavage. The mice were fasted for 16 h, and a glucose tolerance test was performed between 8:00 AM and 12:00 PM. After the collection of an unchallenged sample (time 0), a solution of 50% glucose (2.0 g/kg body weight) was administered by oral gavage. During the test, blood was collected from the tail vein 30, 60, and 120 min after glucose administration. All blood glucose measurements were performed using a hand-held glucose meter.
Western blot analysis
Epididymal fat-pads from each mouse were homogenized at 4°C in extraction buffer (100 mM Tris-HCl, pH 7.4, 5 mM EDTA, 50 mM NaCl, 50 mM sodium pyrophosphate, 50 mM NaF, 100 mM orthovanadate, 1% Triton X-100, 1 mM phenylmethanesulfonylfluoride, 2 mg/mL aprotinin, 1 mg/mL pepstatin A, and 1 mg/mL leupeptin). The tissue homogenates were centrifuged (13,000 g, 20 min, 4°C), and the resulting supernatants (total lysate) were used for western blot analysis. Plasma membrane proteins were extracted from the epididymal adipose tissue using a commercially available protein extraction kit (Bio-Rad), according to the manufacturer's instructions. The total protein concentrations of whole-tissue and plasma membrane extracts were determined by the Bradford assay (Bio-Rad).
Protein samples were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto a nitrocellulose membrane (Amersham), which was incubated with primary antibodies (diluted 1:250–1000) overnight at 4°C. After the membrane had been incubated with the relative secondary antibody (Santa Cruz), immunoreactive signals were detected using a chemiluminescent detection system (Amersham) and quantified using Quantity One analysis software (Bio-Rad). Antibodies to the following proteins were purchased from the indicated sources: ERK, phosphorylated (p-)ERK (Thr202/Tyr204), S6K1, p-S6K1 (Thr389), AKT, p-AKT (Ser473), and GAPDH were obtained from Cell Signaling; GLUT4 was obtained from Santa Cruz; Caveolin was obtained from BD Transduction Laboratories.
Statistical analysis
All data are presented as the mean±standard error of the mean (SEM) of eight mice. The western blot data is presented as the mean±SEM of the protein samples analyzed in triplicate. Data regarding changes in body weight and plasma glucose level were analyzed using multivariate analysis of variance. Comparisons among groups were made using one-way analysis of variance with Duncan's multiple-range tests or the Kruskall–Wallis test followed by the Mann–Whitney U test. A P value of <.05 was considered statistically significant. Statistical analyses of the data were performed using SPSS statistical software (Version 12.0).
Results
Phytochemical analysis
Phytochemical analysis of the DB rhizome extract revealed the presence of allantoin (Fig. 1). Diosgenin and dioscin were not detected in the DB extract used in the present study (data not shown). The HPLC chromatogram of the standard allantoin solution showed an absorption peak with a retention time of 13.642 min, which was also present in the HPLC chromatogram of the DB extract, indicating the presence of allantoin (Fig. 1). The HPLC analysis showed high amounts of allantoin (2.33%, w/w) in the dried DB extract.
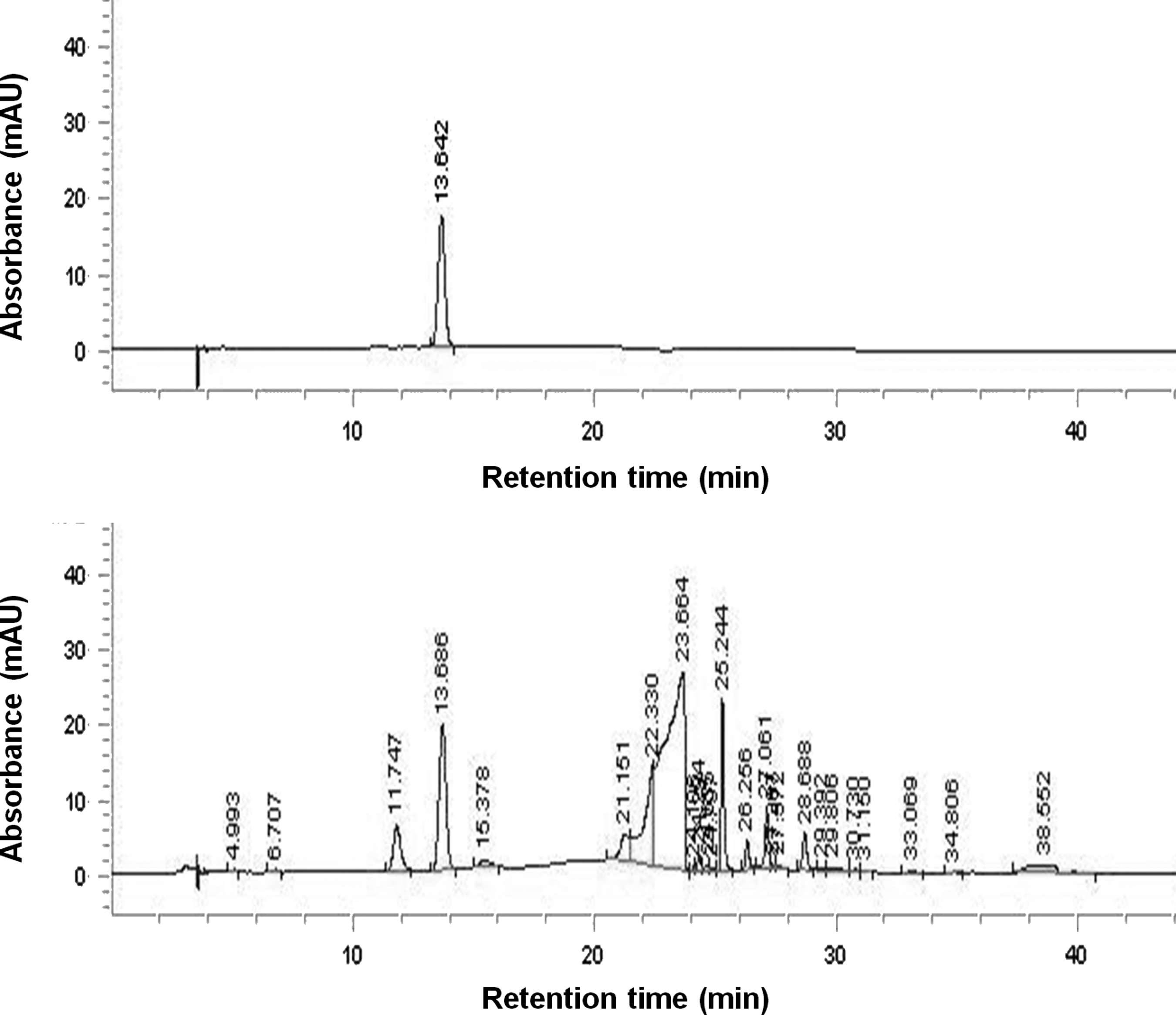
HPLC chromatogram of DB extract.
Body weight and blood biochemistries
DB treatment did not affect the body weight of HFD-fed mice during the experimental period (Fig. 2A). Fasting blood glucose levels, which were measured every week, were significantly reduced in mice administered the DB extract compared with those of the HFD mice (Fig. 2B). At the end of the 7 weeks of treatment, the mice administered the DB extract showed a significant decrease (−33%) in blood glucose levels compared with the HFD mice (Fig. 2C). Similarly, the DB extract effectively reversed (−24%, P<.05) the HFD-induced elevation in plasma insulin level (Fig. 2D). Insulin sensitivity was also significantly increased by the administration of the DB extract as indicated by a 37% reduction in IGR (Fig. 2E).
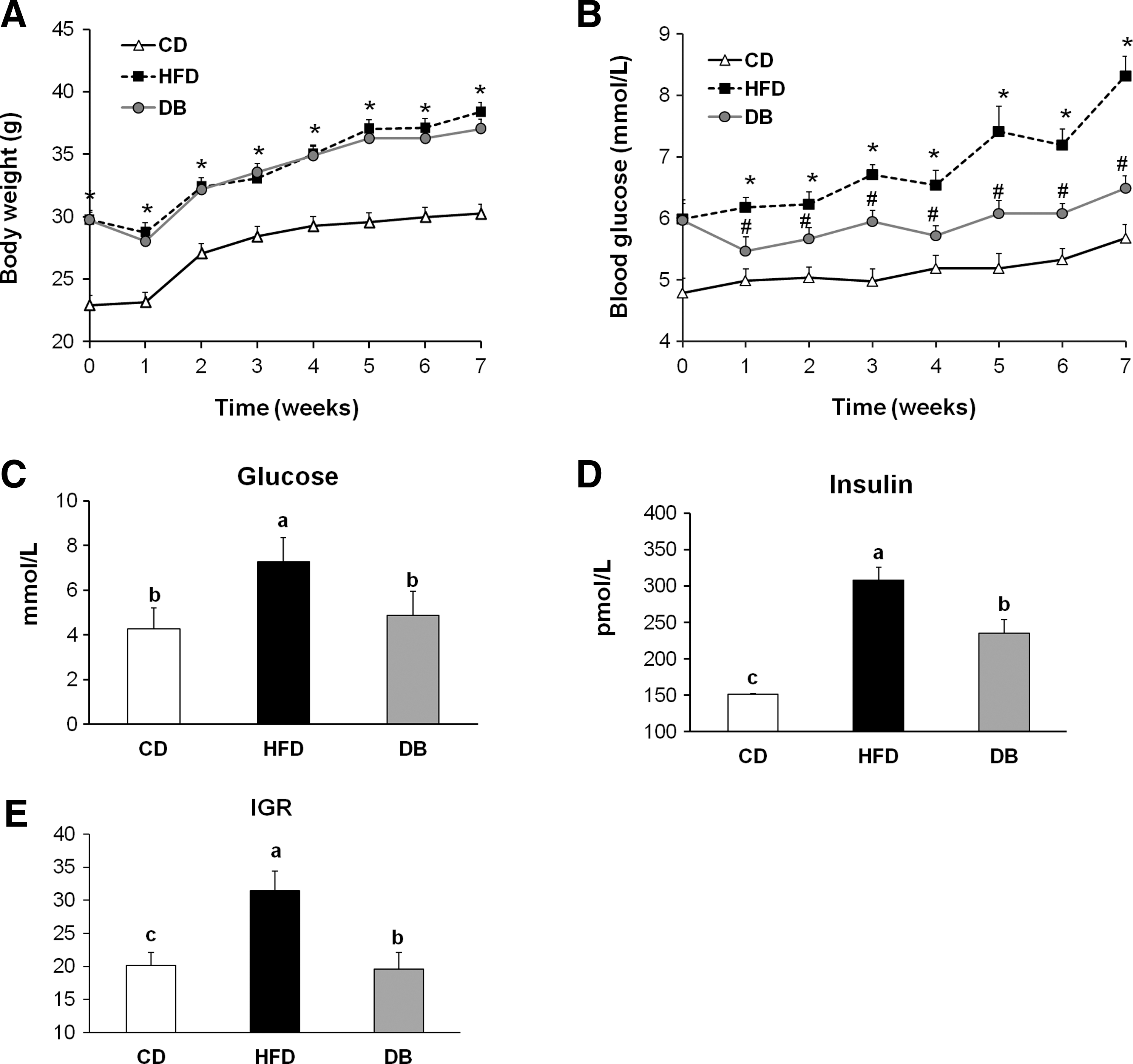
The effects of the DB extract administration on body weight and blood biomarkers of insulin resistance in mice fed the HFD.
Oral glucose tolerance test
After 6 weeks of treatment, the mice were subjected to an OGTT. The DB mice had significantly lower blood glucose concentrations at 0 (−16%), 30 (−12%), 60 (−16%), and 120 (−11%) minutes during the OGTT compared with the HFD mice. The glucose area under the curve observed for the DB mice was significantly lower (−13%) than the value for the HFD mice, and was comparable to that for the CD mice (Fig. 3).
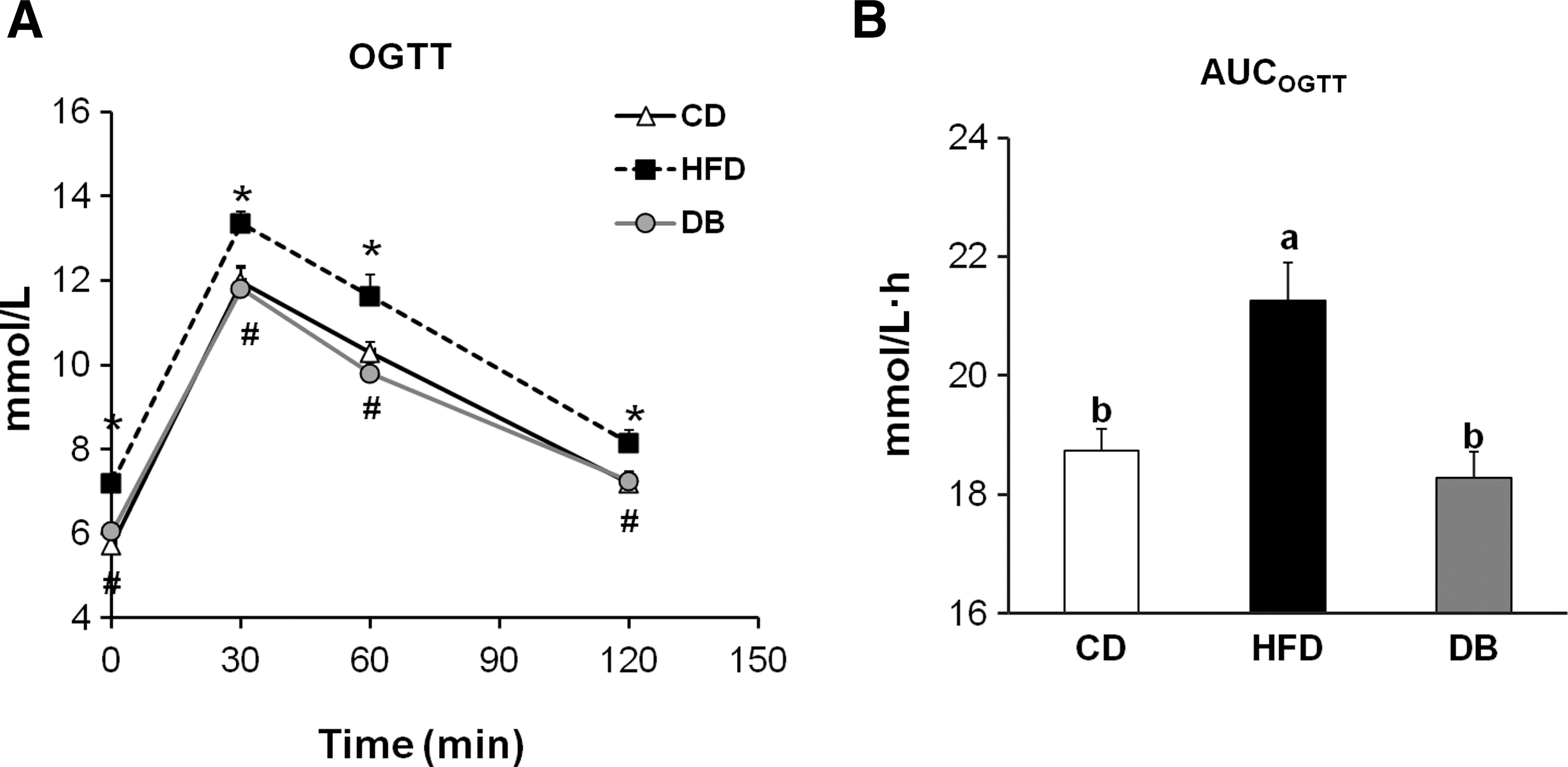
The effect of the DB extract on oral glucose tolerance in mice fed the HFD. Mice were fasted for 16 h and then given a 50% glucose solution (2.0 g/kg body weight) by oral gavage.
GLUT4 protein concentration
We investigated whether the DB treatment influenced the translocation of GLUT4 from the cytosol to the plasma membrane in the adipose tissue of HFD-fed mice. Results from the western blot analysis of epididymal adipose tissue revealed that although the total GLUT4 protein concentration among the experimental groups was similar (Fig. 4A), the level of GLUT4 protein in the plasma membrane fraction was significantly reduced in the HFD mice (vs. the CD mice) and was elevated in the DB mice (vs. the HFD mice; Fig. 4B).
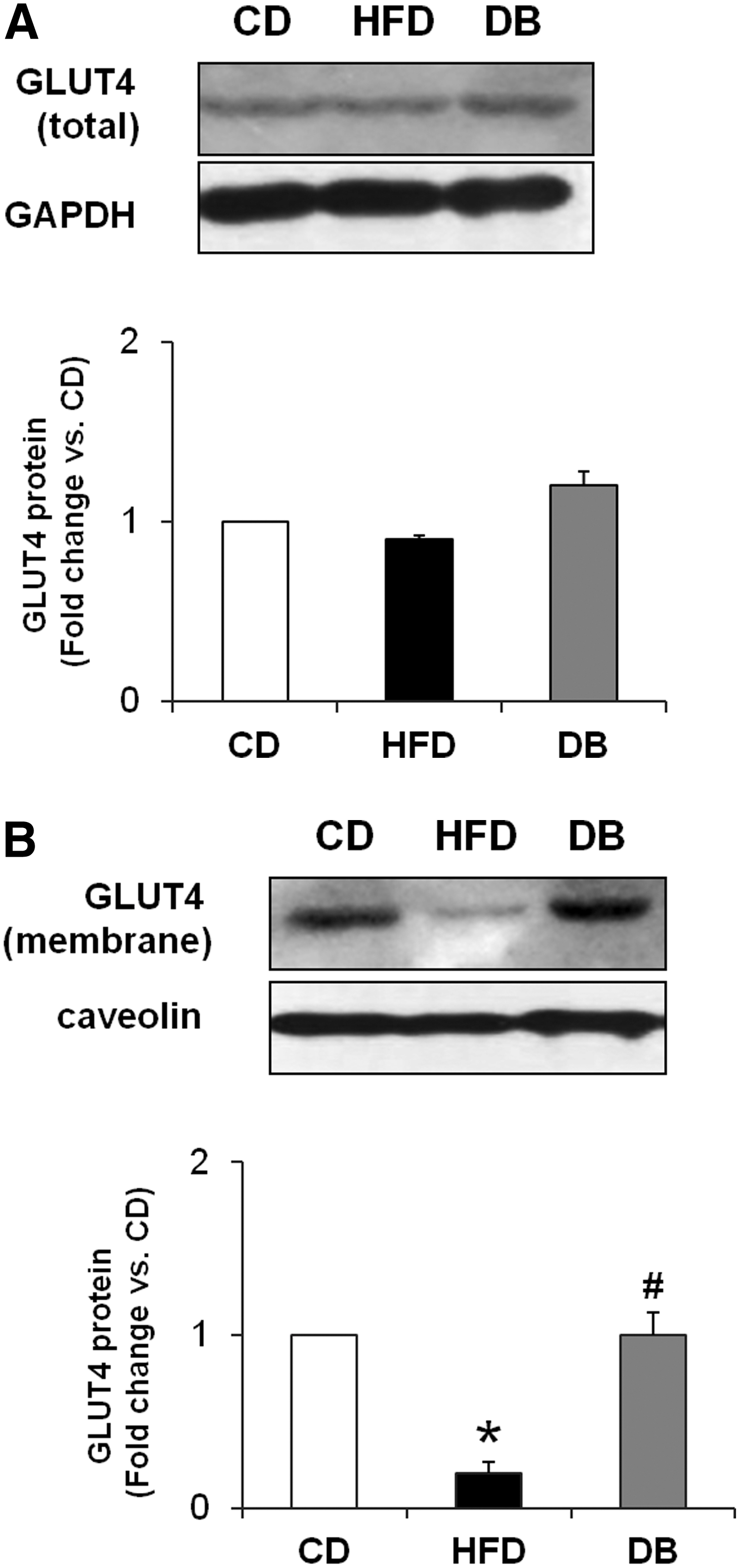
The effect of the DB extract on GLUT4 translocation in the visceral adipose tissue of mice fed the HFD.
Expression of insulin signaling molecules
Western blot analyses of proteins isolated from epididymal adipose tissues revealed that the p-AKT (Ser473) protein expression was significantly decreased, whereas the p-ERK (Thr202/Tyr204) protein level was significantly increased in the HFD mice as compared with the CD mice. The DB extract significantly reversed the HFD-induced changes in the expression of p-AKT and p-ERK (Fig. 5A, B). In addition, the DB extract significantly reversed the HFD-induced up-regulation of phosphorylated S6K1 (Thr389) protein, a downstream target of mTOR, in the visceral adipose tissue of mice (Fig. 5C).
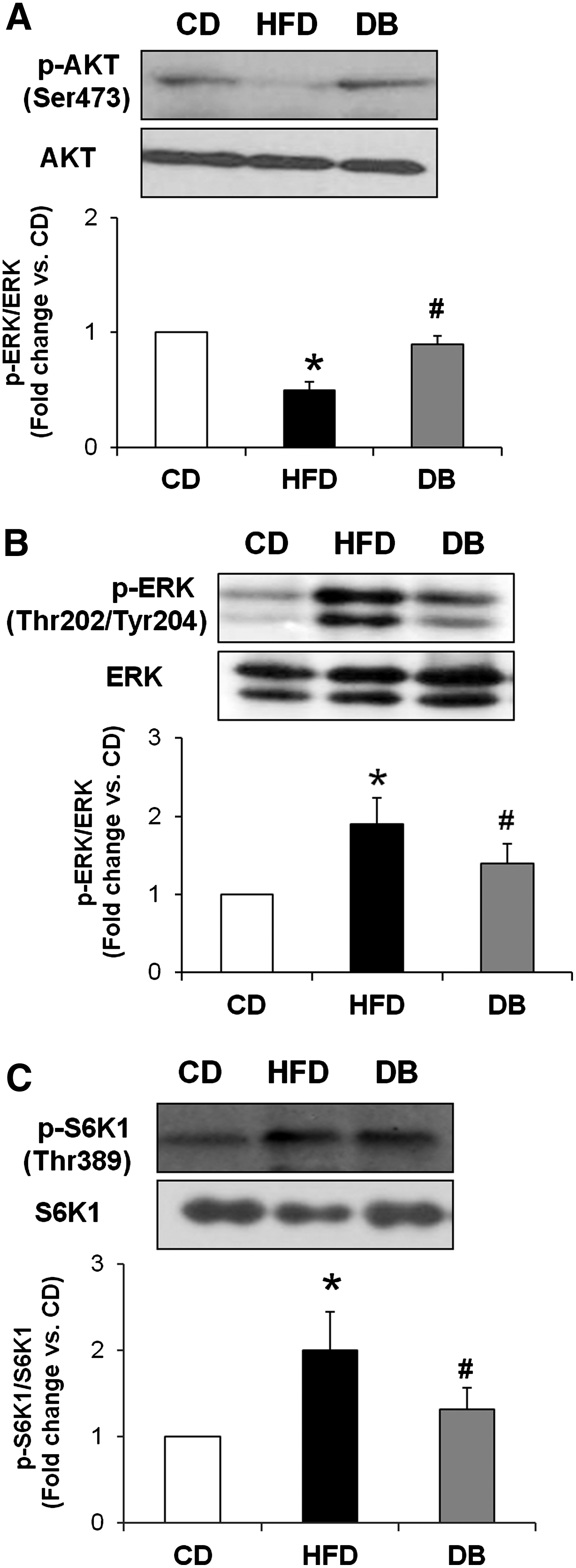
The effect of the DB extract on insulin signaling molecules in the visceral adipose tissue of mice fed an HFD. The proteins in the whole tissue extract (80 μg/lane) from epididymal adipose tissue were analyzed by western blotting using the following antibodies:
Discussion
HFD feeding, a commonly studied model of insulin resistance in rodents, rapidly causes progressive metabolic maladies. 23,24 Insulin resistance in adipose tissue, liver, and muscle, and hyperglycemia and hyperinsulinemia are observed as significantly robust phenotypes as early as 1–4 weeks of feeding with an HFD. 23,25 –28 We investigated the role of the DB extract in the early development of insulin resistance using a 4-week HFD feeding model. In our preliminary studies, oral administration of the DB extract at a dose of 100 mg/kg body weight for 7 weeks was shown to reduce fasting blood glucose levels in genetically obese ob/ob and db/db mice (data not shown). Based on these observations, the DB extract was administered to the mice at a dose of 100 mg/kg body weight in the present study. Animals in CD, HFD, and DB groups failed to attain increases in body weight during the first week of oral administration, but exhibited increased body weight thereafter. This might be attributable to the mice being stressed by oral administration during the first week, which was overcome eventually. The same tendency to decrease body weight was observed during the first week of oral administration in reports by other investigators. 29,30 Although there was no significant change in body weight in mice treated with the DB extract compared with the HFD-fed mice at the end of the study, there were significant decreases in fasting plasma glucose and insulin levels. These results are consistent with a previous report showing that orally administered D. opposite (Dioscoreaceae) at doses of 75–300 mg/kg for 2 weeks decreased fasting plasma glucose and insulin levels in dexamethasone-induced diabetic rats. 22 In the present study, fasting plasma glucose concentrations increased steadily from 0 to 7 weeks in CD-fed mice, although the difference between 0 and 7 weeks was not statistically significant. This tendency of increased fasting glucose level in animals fed a normal diet as they become older was evident in the previous reports as well. 31 –33
In obese states, adipocytes release cytokines, adipokines, and free fatty acids, which can act in a paracrine or autocrine fashion to amplify the proinflammatory state within adipose tissue and cause localized insulin resistance. Adipose tissue also serves as an endocrine organ whereby cytokines, adipokines, and free fatty acids travel to the liver and muscle and may lead to systemic insulin resistance. 2 The importance of white adipose tissue in maintaining peripheral insulin sensitivity has been previously demonstrated; adipose tissue-specific reduction of GLUT4 in mice resulted in impaired glucose tolerance and peripheral insulin resistance. 34 In addition, impairment of the glucose transport pathway in adipose tissue has been demonstrated during insulin resistance and prediabetes. 35 –37 To explore the mechanisms by which the DB extract improves insulin sensitivity, we focused on the possibility that the DB extract could increase GLUT4 content in the plasma membrane of epididymal fat depots. The present study demonstrates that the DB extract significantly reversed the HFD-induced decreases in GLUT4 translocation to the plasma membrane of the visceral adipose tissue in mice. This finding is supported by a previous report showing that the DB extract increased the GLUT4 mRNA level in insulin-treated 3T3-L1 adipocytes. 22
In adipose tissue, activation of p-ERK and p-AKT, the two major downstream kinases activated by insulin, is considered an essential step in the translocation of GLUT4 from the cytosol to the plasma membrane on insulin stimulation. Insulin resistance in the visceral adipose tissue of HFD mice was characterized by elevation of ERK phosphorylation and attenuation of AKT phosphorylation. A previous study demonstrated that AKT interacted with Raf and phosphorylated this protein at a highly conserved serine residue in its regulatory domain in vivo. 38 This phosphorylation of Raf by the AKT inhibited activation of the Raf-MEK-ERK signaling pathway. 38 Under conditions of nutrient overload associated with the obese state, constitutive activations of ERK and S6K1 lead to desensitization of insulin signaling through IRS-1 serine phosphorylation, leading to insulin resistance. 39 Several studies reported that S6K1 and ERK phosphorylations were highly elevated in animals fed a HFD, whereas AKT activity was blunted in adipose tissue. 10,40 In DB extract-treated mice, p-ERK and p-S6K1 protein expressions were down-regulated, whereas p-AKT protein expression was up-regulated compared with that in HFD mice. Based on these observations, we speculated that the DB extract could attenuate insulin resistance in mice fed an HFD, probably through the suppression of insulin signaling via the mTOR/S6K1 pathway.
Allantoin is recognized as an abundant and active component in yam (Dioscorea spp.). 41 In the present study, HPLC analysis of the DB extract revealed a high amount of allantoin (2.33%, w/w). A recent study reported that allantoin increased glucose uptake directly in the isolated skeletal muscle of streptozotocin-induced diabetic rats through the activation of GLUT4 translocation. 42 In addition, plasma glucose levels of streptozotocin-induced diabetic rats were reduced after repeated intravenous injections of allantoin (0.5 mg/kg, three times a day) for 3 days. 42 Although it is possible that the allantoin in the DB extract was responsible for the observed effects in the present study, further studies are needed to determine whether the protective effects of allantoin against HFD-induced insulin resistance are manifested in the visceral adipose tissue of mice as well.
The results of this study revealed that the DB extract improved glucose metabolism through the up-regulation of GLUT4 content in the plasma membrane of visceral adipose tissue in HFD-fed mice. Activation of the insulin signaling cascade leading to GLUT4 translocation was the underlying mechanism for the beneficial effects of the DB extract in the early stage of obesity-induced insulin resistance. Therefore, dietary supplementation with this extract, if validated in human studies, may provide a herbal adjunctive therapy for the treatment of diet-induced insulin resistance.
Footnotes
Acknowledgment
This research was supported by the Industrialization Support Program for Bio-technology of Agriculture and Forestry (810002031SB110), Ministry for Food, Agriculture, Forestry, and Fisheries, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.