
Abstract
To investigate neuronal cell protective effects of an ethyl acetate fraction from chestnut inner skin, in vitro assays, including 2′,7′-dichlorofluorescein diacetate, 3-[4,5-dimethythiazol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT), and lactate dehydrogenase (LDH), were performed. Intracellular accumulation of reactive oxygen species resulting from hydrogen peroxide (H2O2) treatment of PC12 cells was significantly reduced when ethyl acetate fractions were present in the medium compared to PC12 cells treated with H2O2 only. In a cell viability assay using MTT, the ethyl acetate fraction protected against H2O2-induced neurotoxicity, and inhibited LDH release into the medium. In addition, the ethyl acetate fraction improved in vivo cognitive ability against amyloid β-peptide (Aβ)–induced neuronal deficit. High-performance liquid chromatography analyses showed that gallic acid, catechin, and epicatechin were predominant phenolics in the ethyl acetate fraction. Consequently, the results suggest that chestnut inner skin, including above phenolics, could ameliorate Aβ-induced learning and memory deficiency, and be utilized as effective substances for neurodegenerative disorders, notably Alzheimer's disease.
Introduction
O
Naturally occurring antioxidants have been reported to play a major role in ameliorating oxidative damage induced by free radicals. Recently, natural foods and food-derived components, such as antioxidative vitamins and phenolic phytochemicals, have received a great deal of attention because they are safe and not perceived as medicine; some of these are known to function as chemopreventive agents against oxidative damage. 9 Chestnut inner skin is also rich in carbohydrates and is a good source of essential fatty acids and minerals. 10 In addition, they have several vitamins and appreciable levels of fiber. 11 Some researchers also reported that chestnut inner skin contains gallic acid, epicatechin, and catechin as its main phenolics. 12 Although it has already been demonstrated that chestnut fruits and leaves contain phenolics and antioxidant activities, 13 little is known about their protective effect on in vivo antiamnesic effects. Therefore, in vitro neuronal cell protective effects of the ethyl acetate fraction from chestnut inner skin against H2O2-induced cell death in PC12 cells were examined. In addition, its effects on Aβ-induced cognitive deficits in mice using in vivo learning and memory tests such as the Y-maze test were evaluated. According to the oxidative stress theory of AD, Aβ generates free radicals in neuronal cell membrane bilayers and then causes cellular dysfunction through lipid peroxidation. 14 –16 Therefore, cognitive dysfunction caused by neurotoxic Aβ has a direct correlation between oxidative stress-induced neurocitoxicity and neurodegeneration. Consequently, the present study was undertaken to investigate the antiamnesic effects of the ethyl acetate fraction from chestnut inner skin on Aβ-induced cognitive dysfunction in mice.
Materials and Methods
Chemicals
2′,7′-Dichlorofluorecein diacetate (DCF-DA), vitamin C, H2O2, dimethyl sulfoxide, HEPES, sodium bicarbonate, penicillin, streptomycin, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay kit, lactate dehydrogenase (LDH) assay kit, Aβ25–35, and all other materials used were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Neuronal cell culture
PC12 cells (KCLB 21721; Korea Cell Line Bank, Seoul, Korea), which were derived from a transplantable rat pheochromocytoma, were propagated in an RPMI 1640 medium (Gibco BRL, Grand Island, NY, USA) containing 10% fetal bovine serum, 25 mM HEPES, 25 mM sodium bicarbonate, 50 units/mL penicillin, and 100 μg/mL streptomycin.
Animals and in vivo experimental design
Institute of cancer research mice (male, 4-week old) were obtained from Samtako (Osan, Korea). The mice were housed two per cage in a room maintained with a 12-h light–dark cycle, 55% humidity, and 23°C–25°C temperature. The ethyl acetate fraction of chestnut inner skin was mixed in drinking water at concentrations of 5, 10, and 20 mg/kg of body weight. All experimental procedures were approved by the guidelines established by the Animal Care and Use Committee of Gyeongsang National University. Subsequently, Aβ25–35 was administered via intracerebroventricular (ICV) injection. Control animals were injected with Aβ25–35 (Fig. 1). Aβ was dissolved in 0.85% sodium chloride solution (w/v) and injected intracerebroventricularly into each mouse with a 25-μL Hamilton microsyringe fitted with a 26-gauge needle that was inserted to a depth of 2.5 mm. The injection volume was 5 μL (dose 410 pmol/mouse). 17

In vivo experimental design for amyloid β-peptide (Aβ)-induced amnesic mouse model.
Plant materials and extraction
Chestnut inner skin was purchased from (Altori Co., Ltd. Jinju, Korea) in September 2010. The samples were washed with running tap water before being chopped into pieces. They were then oven-dried at 40°C for 2 days, and ground to a powder. The ethyl acetate fraction from the water extract of chestnut inner skin was obtained as follows. Powder of chestnut inner skin (100 g) was suspended and extracted with 500 mL of water at 70°C for 2 h. Water extracts were filtered through Whatman No. 2 filter paper (Whatman International Limited, Kent, England, United Kingdom), and evaporated to dryness. The dry materials were redissolved in 500 mL of distilled water. The solution was consecutively partitioned in a separatory funnel with the equivalent amount of ethyl acetate. The fraction was concentrated in a vacuum evaporator (N-N series; Eyela Co., Tokyo, Japan) at 40°C. This sample was stored at −20°C until use.
Measurement of intracellular oxidative stress
Levels of intracellular ROS were determined by DCF-DA assay. 18 In brief, cells (104 cells/well on 96-well) were treated for 48 h with the indicated concentrations of the ethyl acetate extract of chestnut inner skin or vitamin C. The cells were then treated with or without 200 μM H2O2 for 2 h. At the end of the treatment, cells were incubated in the presence of 50 μM DCF-DA in phosphate-buffered saline. Fluorescence was then quantified using a TECAN SER-NR 94572 fluorometer (San Jose, CA, USA) using 485-nm excitation and 530-nm emission filters.
Protection of neuronal cells against oxidative stress
MTT reduction assay was determined using the in vitro toxicology assay kit (TOX-1; Sigma). Neuronal PC12 cells were plated at a density of 106 cells per well on 96-well plates in 100 μL of RPMI. The cells were preincubated with the ethyl acetate extract of chestnut inner skin for 48 h before the 200 μM H2O2 was added. The cells were treated with or without H2O2 for 2 h. The amount of MTT formazan product was determined by measuring absorbance using a microplate reader (680; Bio-Rad, Tokyo) at a test wavelength of 570 nm and a reference wavelength of 690 nm. 19
Neuronal PC12 cells were precipitated by centrifugation at 250 g for 4 min at room temperature; 100 μL of the supernatants was transferred into new wells, and LDH was determined using the in vitro toxicology assay kit (TOX-7; Sigma). Damage of the plasma membrane was evaluated by measuring the amount of the intracellular enzyme LDH released into the medium. 19
Y-maze test
Recording spontaneous alternation behavior in the Y-maze test was used to assess the immediate working memory performance. The Y-maze test was performed 2 days after the Aβ injection. The maze was made of black-painted plastic, and each arm of the maze was 33-cm long, 15-cm high, and 10-cm wide and was positioned at an equal angle. Each mouse was placed at the end of one arm and allowed to move freely through the maze for 8 min. The series of arm entries was recorded visually, and arm entry was considered to have been completed only when the hind paws of the mouse were placed completely in the arm of the maze. Alternation is defined as successive entries into the three arms in overlapping triplet sets. The percentage alternation was calculated as the ratio of actual to possible alternation (defined as the total number of arm entries minus two), multiplied by 100. 20
Determination of total phenolics
The total phenolics were determined by a spectrophotometric method. 12 The standard curve for total phenolics was made using the gallic acid standard solution (0–100 mg/L). The total phenolics in aqueous extract of propolis were expressed as milligrams of gallic acid equivalents per 1 g of sample.
Quantification by HPLC
Phenolics in ethyl acetate fraction from chestnut inner skin were measured at 280 nm by a photo diode array detector (U 3000 series; Dionex, Sunnyvale, CA). Separation was achieved with a Shiseido C18 column (250 mm×4.6 mm id, 5 μm; Shiseido Co., Tokyo). The elution solvents were (A) 0.01 M potassium phosphate buffer adjusted to pH 3.0 by phosphoric acid and (B) methanol. The solvent gradient elution program used was as follows: initial 90% (A), hold for 9.5 min; linear gradient to 68% (A) in 3.5 min; linear gradient to 67% (A) in 17 min; linear gradient to 20% (A) in 1 min; linear gradient to 90% (A) 1 min, and hold for 10 min. The flow rate was 1.5 mL/min. Phenolics were identified by comparison of their retention time (RT) and UV spectra with those of known standards, and quantified by peak areas from the chromatograms. All analyses were run in triplicate, and mean values were calculated. Content of phenolics was expressed in mg/100 g extract.
Statistical analysis
All data were expressed as mean±SD. The data were analyzed by Duncan's new multiple-range test of SAS (Cary, NC, USA). The statistical significance of differences among groups was calculated by a one-way analysis of variance.
Results and Discussion
Measurement of intracellular oxidative stress
To examine intracellular accumulation of ROS, we used a DCF-DA probe that is freely permeable to the cell membrane. In PC12 cells, the compound is hydrolyzed by the cellular esterase to DCF, which interacts with peroxide-forming fluorescent DCF. Exposure of PC12 cells to H2O2 for 2 h resulted in a 130.66% increase of ROS levels compared to control (Fig. 2). Intracellular ROS accumulation resulting from H2O2 treatment was significantly decreased when the ethyl acetate extract of chestnut inner skin was present in the medium compared to PC12 cells treated with H2O2 only. Vitamin C, a naturally occurring major nutrient, has antioxidant activity. At the level of 200 μM vitamin C (101.89%) as a positive control, PC12 cells had significantly lower oxidative stress than PC12 cells treated with H2O2 only. Oxidative stress in AD may result from aging, energy deficiency, inflammation, or excessive production of Aβ. Aβ protein can induce cell death through a mechanism involving H2O2 production. 16 In this respect, this result suggests that the ethyl acetate extract of chestnut inner skin with antioxidant activity may decrease the oxidative stress-induced risk by AD.
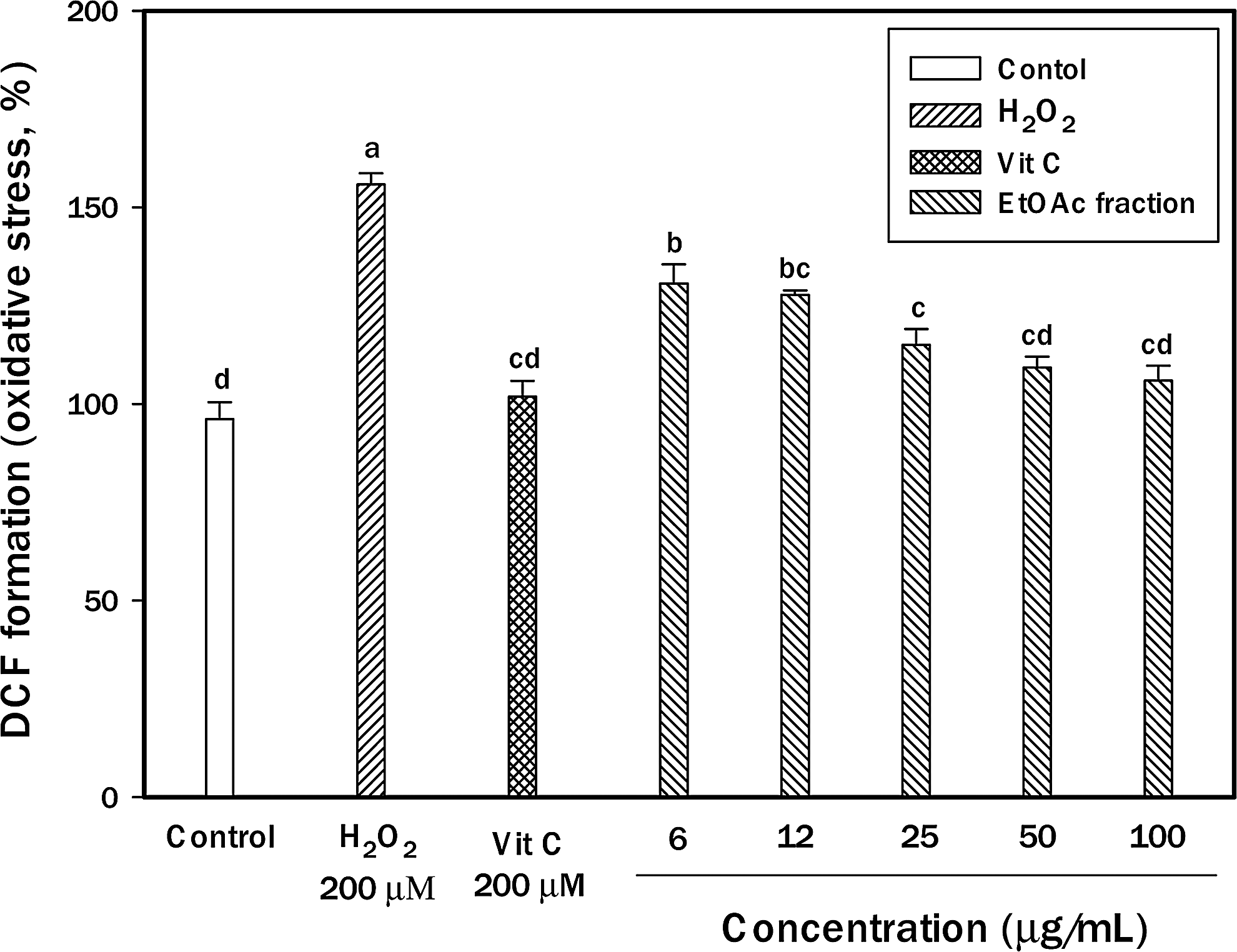
Protective effect of ethyl acetate fraction from chestnut inner skin against cellular oxidative stress. PC12 cells were pretreated for 48 h with various concentrations of the extract fraction. After 48 h, the cells were treated with 200 μM hydrogen peroxide (H2O2) for 3 h. The increase of 2′,7′-dichlorofluorecein diacetate (DCF-DA) fluorescence was measured by a fluorescence microplate reader. Results are presented as mean±SD of three independent experiments in triplicate. abcdDifferent letters indicate significant differences at P<.05.
Neuronal cell protective effects on oxidative stress
H2O2 has been reported to induce apoptosis in cells of the central nervous system. In this study, the ethyl acetate fraction of chestnut inner skin was selected to investigate the neuroprotective effects against H2O2-induced damage because of strong antioxidant activity. Protective effects on H2O2-induced neuronal cells damage were confirmed by MTT assay. H2O2 caused a decrease in cell viability (32.87%), but pretreatment of PC12 cell with increasing concentrations of the ethyl acetate fraction of chestnut inner skin inhibited oxidative stress-induced cytotoxicity (Fig. 3A). Neuronal cell protective effects of the fraction at 100 μg/mL on oxidative injury were similar to those of 200 μM vitamin C. This study demonstrated that PC12 cell cytotoxicity through oxidative stress was suppressed by pretreatment with the ethyl acetate fraction obtained from chestnut inner skin. MTT dye reduction assay is based on the catalytic activity of some metabolic enzymes in intact mitochondria. Mitochondria may be one of the most sensitive primary targets of oxidative injury in neuronal cells. These results suggest that PC12 cell protection by the ethyl acetate fraction of chestnut inner skin is partially due to the mechanisms with mitochondrial protection.
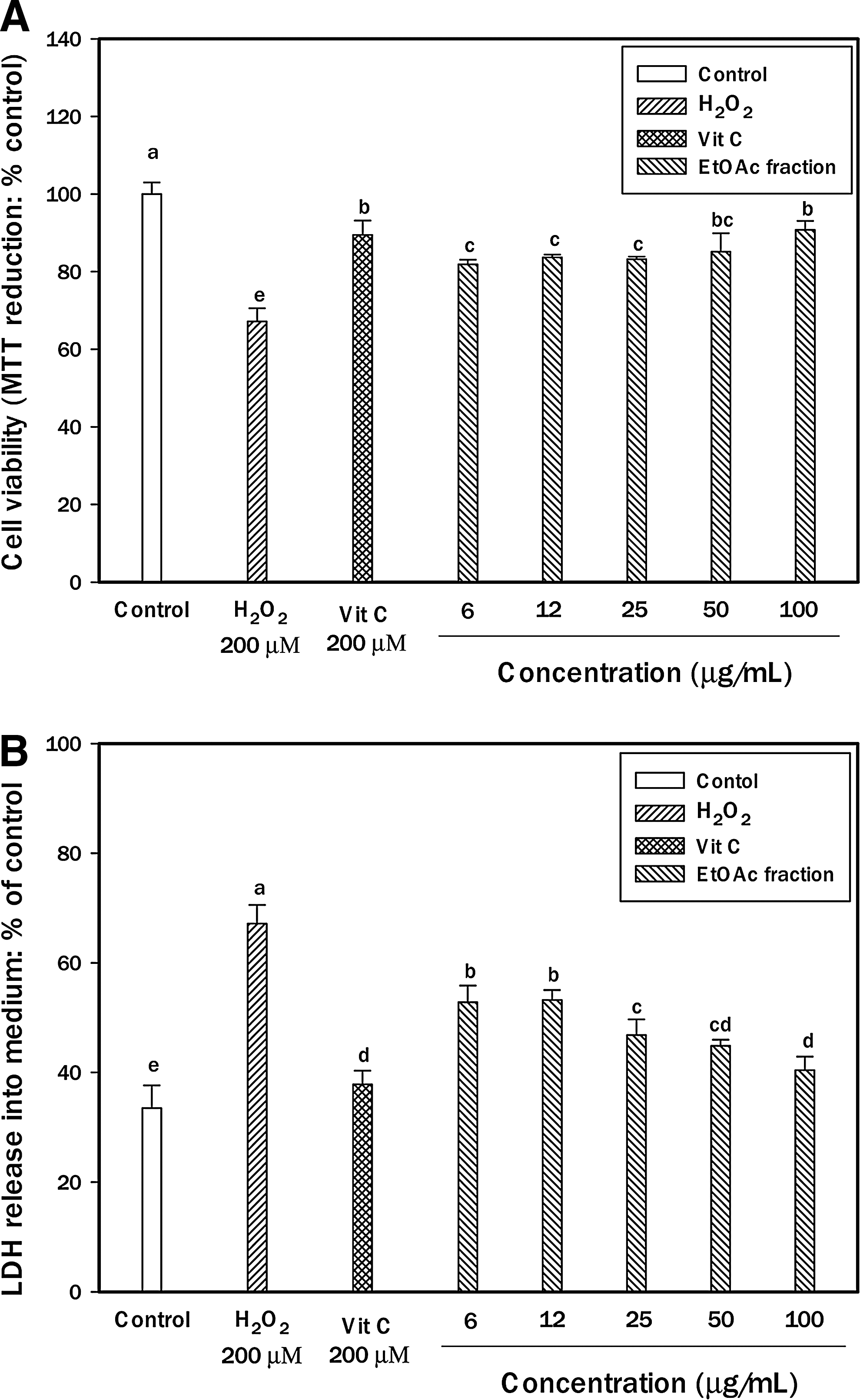
Neuroprotective effect of ethyl acetate fraction from chestnut inner skin on H2O2-induced cytotoxicity in PC12 cells.
Protective effect on H2O2-induced membrane damage
The neuronal membrane contains polyunsaturated fatty acids, which are vulnerable to oxidative stress induced by ROS such as H2O2. Lipid peroxidation can alter the fluidity of the plasma membrane. LDH assay provided an estimate of the percentage of surviving PC12 cells. The LDH assay was used to further investigate the neuroprotective effect of the ethyl acetate fraction of chestnut inner skin. After the treatment of PC12 cells with 200 μM H2O2 for 3 h, the absorbance values at 490 nm increased. With oxidative treatment with H2O2 for 3 h, the amounts of LDH release of PC12 cells increased by 33.64% compared to that of the control without treatment. Compared with their effects on the corresponding H2O2 treatment group, the ethyl acetate fraction of chestnut inner skin was dose-dependently decreased. Especially, the fraction attenuated LDH release significantly to 26.70% of 200 μM H2O2 at 100 μg/mL. In addition, the fraction showed that the pattern of neuronal cell protection was similar to MTT assay. Above results showed that the fraction could protect PC12 cells against the lesion induced by H2O2 (Fig. 3B). Therefore, these data suggest that PC12 cell protection by the ethyl acetate fraction of chestnut inner skin is partially due to its protection of mitochondrial and cell membranes during H2O2-induced neurotoxicity.
Effect of ethyl acetate fraction obtained from chestnut inner skin on Y-maze test
The Y-maze test was carried out 2 days after the Aβ injection. The control group was injected with the nontoxic reverse fragment as Aβ35–25. The Aβ25–35 group was injected with 410 pmol of Aβ25–35 per mouse. Aβ was dissolved in a 0.85% (w/v) sodium chloride solution. In Figure 4A, the administration of Aβ significantly impaired the spatial working memory (83.49%) compared with that of the control group (100%). Pretreatment with the sample increased spontaneous alternation behavior in the Aβ-injected mice (5 mg/kg of ethyl acetate fraction: E5, 90.80%; 10 mg/kg of ethyl acetate fraction: E10, 98.26%; 20 mg/kg of ethyl acetate fraction: E20, 103.98%). In contrast, the number of arm entries did not change among all the experimental groups (Fig. 4B). ICV injection of Aβ induced memory deficits. 21 Therefore, this method of Aβ exposure is a useful in vivo model for Aβ toxicity and suggests that ethyl acetate fraction of chestnut inner skin exhibits protective effect against learning and memory deficits in AD mice.
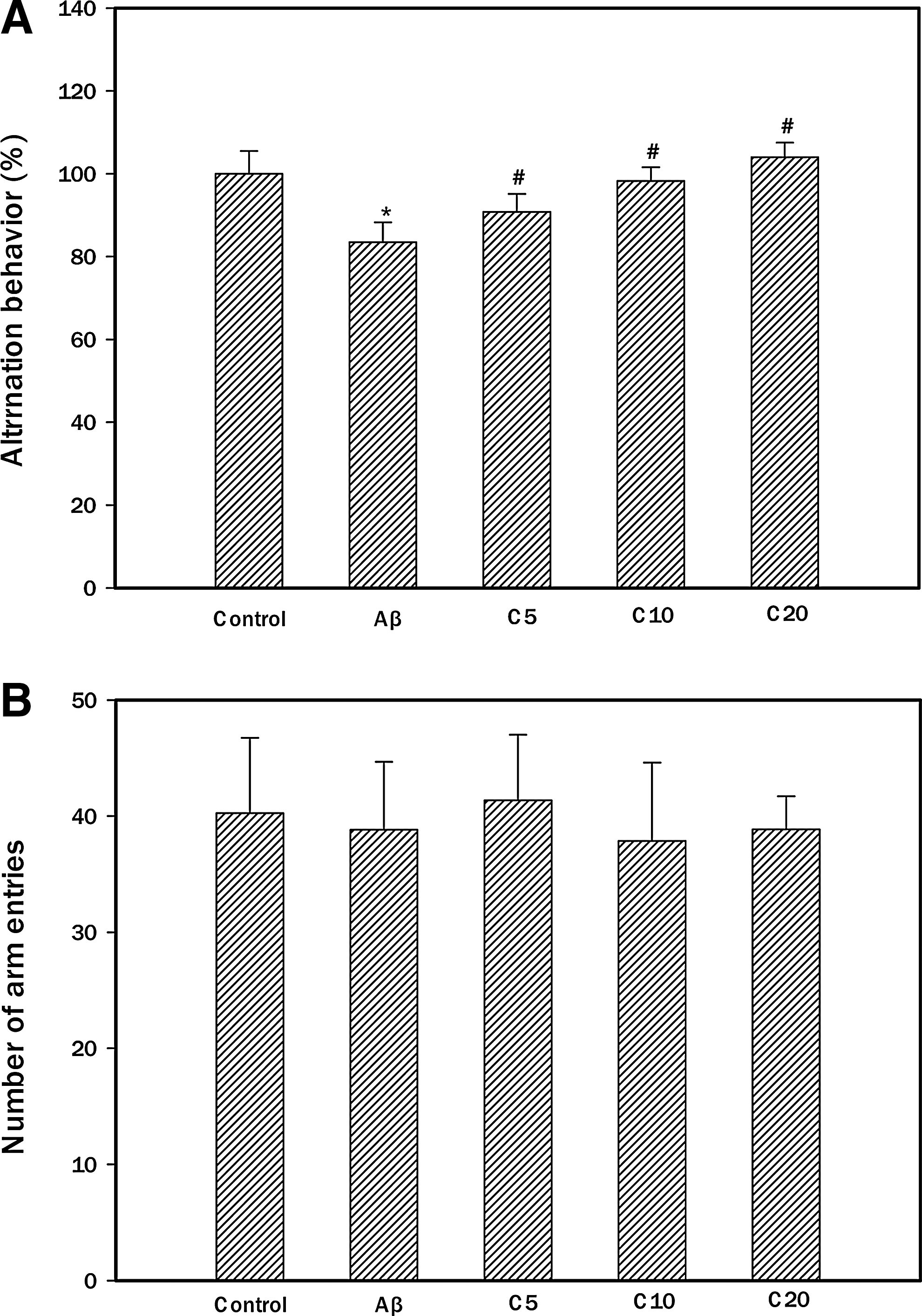
Effect of ethyl acetate fraction from chestnut inner skin against spontaneous alteration behavior in Aβ-induced amnesic mice. The spontaneous alteration behavior
Phenolic composition of ethyl acetate fraction from chestnut inner skin
Phenolics are secondary plant metabolites that are present in natural products. Many of the phenolics have been shown to contain high levels of antioxidant activities. 22 To identify the major neuroprotective phenolics, the ethyl acetate fraction was subjected to further analysis by high-performance liquid chromatography (HPLC), and the fraction showed important phenolics (Table 1). By comparing the RTs and UV spectra of these compounds with those of standards, three phenolics (gallic acid, catechin, and epicatechin) were identified (Fig. 5). Furthermore, the HPLC results indicated that gallic acid (10.90 mg/100 g) was the predominant phenolic in the ethyl acetate fraction, followed by catechin and epicatechin (Table 1). Consequently, the physiological activities determined in ethyl acetate fraction from chestnut inner skin could be attributed to three phenolics, including gallic acid, catechin, and epicatechin. ROS is a key factor involved in the Aβ toxicity. Aβ insertion into the neuronal membrane bilayer generates oxygen-dependent free radicals, causing the lipid peroxidation and protein oxidation. The loss of membrane integrity leads to cellular dysfunction, such as loss of Ca2+ homeostasis, disruption of signal pathways, and activation of nuclear transcription factor and apoptotic pathway. 21 The neuronal apoptosis is the ultimate consequence of these cellular dysfunctions. 23 Some phytochemicals are suggested to have efficacy in the management of oxidative stress-induced cellular dysfunctions such as AD. Many phytochemicals are also known to be potential chemopreventive agents. 24 Therefore, current results suggest that gallic acid, catechin, and epicatechin of the ethyl acetate fraction obtained from chestnut inner skin should mitigate Aβ-induced oxidative stress in neuronal cells, based on their abilities to protect neuronal cell membranes and mitochondrial dysfunction. Also, there are critical criteria for selecting herbal drugs for the treatment of amnesia, including high bioavailability, ability to cross the blood–brain barrier, and minimal toxicity. 24 Based on the studies of Manach et al., 25 the bioavailability of different types of flavonoids is as follows (from most to least bioavailable): isoflavones, gallic acid, catechins, flavones, and quercetin glucosides. In conclusion, the ethyl acetate fraction obtained from chestnut inner skin inhibits neuronal cell death caused by Aβ-induced oxidative stress, and improves Aβ-induced learning and memory deficits.
Values represented the mean±SD (n=3), presented in mg/100 g dry weight.
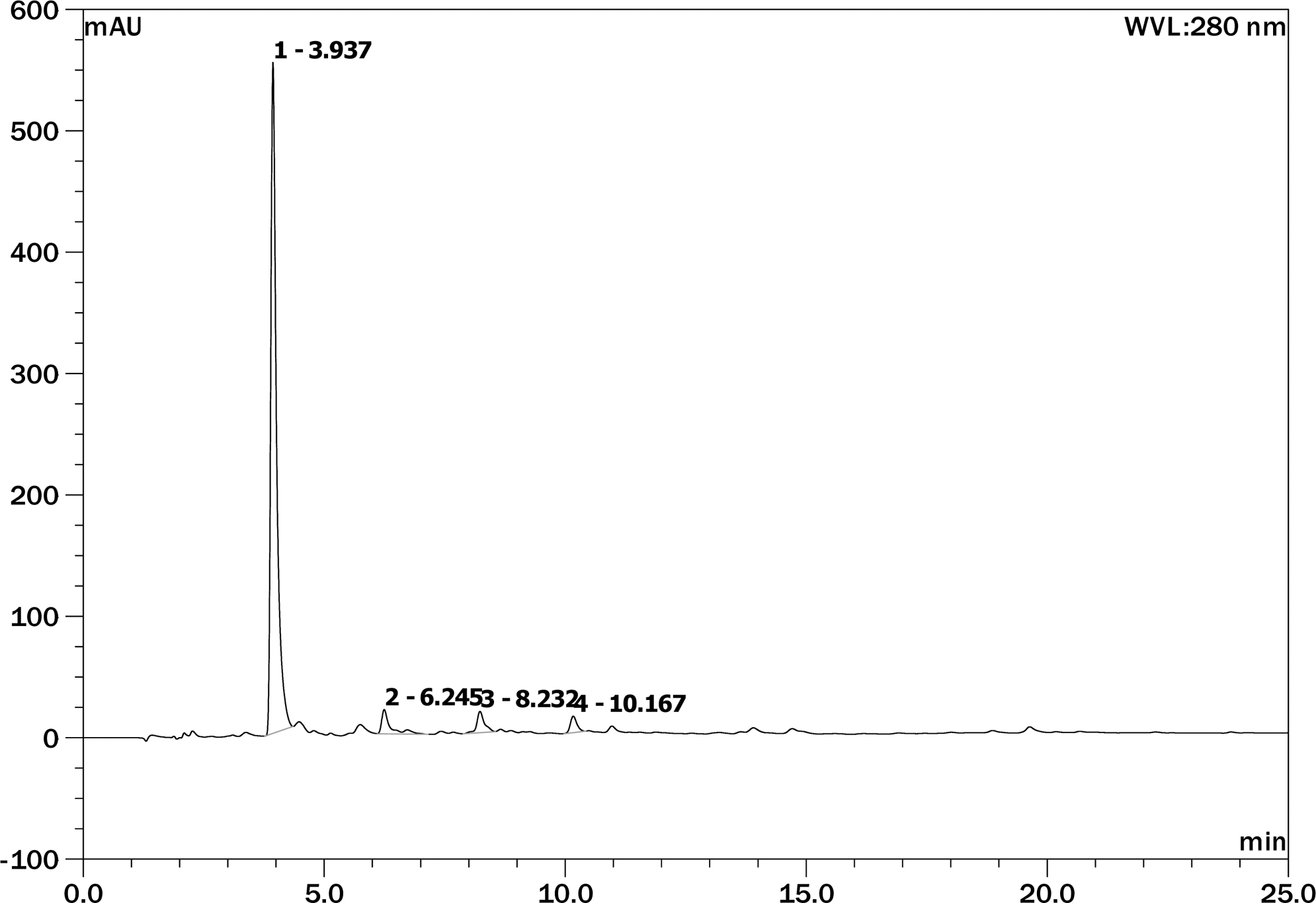
HPLC chromatogram of ethyl acetate fraction from chestnut inner skin. Retention time (min): gallic acid, 3.937; epicatechin, 8.232; catechin, 10.167.
Footnotes
Acknowledgments
This work was supported by the Korea Research Foundation Grant funded by the Korean Government (KRF 2008-521-F00074, KRF-2011-0021664), and the R&D Program (2011) of Korea Forest Research Institute. J.H.J. was supported by BKH program.
Author Disclosure Statement
No competing financial interests exist.