
Abstract
In a previous study, we demonstrated the anti-obesity and hypolipidemic effects of Boussingaulti gracilis Miers var. pseudobaselloides Bailey in high-fat diet-induced obese rats. The present study investigated the molecular mechanisms by which B. gracilis Miers var. pseudobaselloides Bailey ethanol extract (BGE) conferred antidifferentiation and anti-adipogenic effects in the 3T3-L1 preadipocyte differentiation model. BGE treatment significantly and dose-dependently suppressed lipid accumulation and down-regulated the expression of major transcription factors involved in adipogenesis, such as peroxisome proliferator-activated receptor-γ, CCAAT/enhancer binding protein α, sterol regulatory element-binding proteins, and their target genes. It is important that treatment with BGE increased phosphorylation of AMP-activated protein kinase (AMPK), which is one of the rate-limiting enzymes in the fatty acid synthesis pathway, and its direct downstream protein, acetyl-coenzyme A carboxylase. These results suggest that BGE may exert anti-adipogenic effects through regulation of AMPK activity and expression of genes involved in lipogenesis.
Introduction
O
Current medications for the treatment of obesity include mixed noradrenergic–serotenergic agents (sibutramine) and absorption-reducing agents (orlistat). 4 –6 However, their use is limited by their side effects. Sibutramine is known to increase blood pressure and may cause cardiac arrhythmias, constipation, and headache with minimum body weight loss. 4 Furthermore, rimonabant, which is the first selective cannabinoid receptor CB1 blocker, has been officially withdrawn from the market because of its serious psychiatric side effects. 5 It has also been reported that orlistat, the most commonly used medication to treat obesity, causes steatorrhea and deficiencies in lipid-soluble vitamins and essential fatty acids. 6 Thus, interest in natural products for the treatment of obesity is growing rapidly. Natural products derived from medicinal plants have been used as dietary supplements for body weight management in many countries. 4 For example, resveratrol, a root extract from Polygonum cuspidatum, quercetin, rutin, and catechin substantially reduce body weight gain and fat deposition in vivo and in vitro. 7 –11
Boussingaultia gracilis Miers var. pseudobaselloides Bailey is a South American species of ornamental succulent vine. The fresh leaves of Madeira vine are frequently used as vegetables in Taiwan and as folk medicine as a hypoglycemic agent, an anti-inflammatory analgesic, and a stomachics. 12 Recent studies have demonstrated that B. gracilis Miers var. pseudobaselloides Bailey ethanol extracts (BGEs) have inhibitory activities against spasmogen-induced contractions of the isolated gastric fundus 13 and ethanol-induced gastric lesions 14 in rats. In addition, hot water extracts of B. gracilis Miers var. pseudobaselloides Bailey have antiviral activities against herpes simplex virus and adenovirus type 3. 15 It has also been reported that the butanolic fraction from B. gracilis Miers var. pseudobaselloides Bailey exhibited antidiabetic effects in alloxan-induced mice. 16
Previously, we have shown that BGE has anti-obesity and hypolipidemic effects in an animal model. 17 BGE treatment effectively rescues biochemical parameters, such as serum lipid and adiponectin levels, and expression of transcription factors related to lipid and glucose metabolism from obese rats. However, the molecular mechanism of obesity inhibition by BGE remains unclear. Intrigued by our previous results, we further investigated the effect of BGE on the differentiation of 3T3-L1 cells and sought to understand the underlying cellular mechanism of BGE. In the current study, we conducted biochemical assays to determine the effect of BGE on lipid accumulation in 3T3-L1 preadipocytes. Reverse transcription (RT)–polymerase chain reaction (PCR) was used to explore whether BGE treatment affects the expression of genes related to lipid metabolism in the process of 3T3-L1 differentiation. Furthermore, we also demonstrate the critical effect of BGE on AMP-activated protein kinase (AMPK) activity, which is a key enzyme governing lipid metabolism.
Materials and Methods
Preparation of crude extract
BGE was obtained from the Pharmaceutical Analysis Laboratory of the School of Pharmacy, Shenyang Pharmaceutical University, Shenyang, China. In brief, chopped dried leaves of B. gracilis Miers var. pseudobaselloides Bailey were extracted three times with a 10-fold volume of 70% fermentation ethanol at 85°C for 4 h. The resulting extract was filtered and then lyophilized to yield 22.5% (of the initial amount) powder, and the extracts were stored in a refrigerator (–30°C) for functional studies. The extract powder contained 29.3% crude protein, 2.5% crude fat, and 13.0% fiber, as well as 53.5 mg/g total flavonoid. The extract was analyzed by the Pharmaceutical Analysis Laboratory of the School of Pharmacy, Shenyang Pharmaceutical University. 17
Materials
Dulbecco's modified Eagle's medium (DMEM) (catalog number SH30243.01), bovine calf serum (catalog number SH30626.03), fetal bovine serum (catalog number SH30396.03), and penicillin–streptomycin solution (catalog number SV30010) were purchased from HyClone Laboratories, Inc. (Logan, UT, USA). Protein assay reagent (protein assay dye reagent concentrate; catalog number 500-0006) was purchased from Bio-Rad (Hercules, CA, USA). Antibodies against phospho-AMPK (Thr172), acetyl-coenzyme A (CoA) carboxylase (ACC), and phospho-ACC (Ser79) were purchased from Cell Signaling Technology (Beverly, MA, USA). Antibodies against β-actin and AMPK were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Immobilon®-P transfer membranes were purchased from Millipore Corp. (Billerica, MA, USA). The easy-spin total RNA extraction kit was purchased from iNtRON (Seongnam, Korea). RT-PCR primers were synthesized by Bioneer (Daejeon, Korea). 6-[4-(2-Piperidin-1-yl-ethoxy)-phenyl]-3-pyridin-4-ylpyrrazolo[1,5-a]-pyrimidine (compound C) was obtained from Calbiochem (Billerica, MA, USA).
Cell culture and sample treatment
3T3-L1 preadipocyte cells from the American Type Culture Collection (Manassas, VA, USA) were cultured at 37°C in a humidified atmosphere of 5% CO2 in DMEM containing 10% bovine calf serum and 1% penicillin–streptomycin solution. To induce differentiation, postconfluent 3T3-L1 preadipocytes (Day 0) were stimulated for 48 h with differentiation medium (DMEM containing 10% fetal bovine serum, 0.5 mM 3-isobutyl-1-methylxanthine, 1 μM dexamethasone, and 5 μg/mL insulin). After stimulation, cells were maintained in DMEM supplemented with 10% fetal bovine serum and 5 μg/mL insulin. Cells were treated with various concentrations of BGE (10, 50, and 100 μg/mL) and compound C (2 μM) from Day 0.
Detection of lipid accumulation
3T3-L1 adipocytes (3×104 cells per well) were seeded into six-well culture plates. Two days after reaching confluence, cells were treated with various concentrations of BGE for 6 days. Free fatty acid (FFA) and triglyceride (TG) levels were determined enzymatically using commercial kits (Asan Diagnostics, Seoul, Korea). To analyze lipid content, the lipids were extracted by the method of Folch et al. 18 FFAs were quantified by the acyl-CoA synthase–acyl-CoA oxidase method 19 using a NEFA-HR kit (Wako Pure Chemical Industries, Osaka, Japan).
RT-PCR
The total RNA was isolated from 3T3-L1 adipocytes using an easy-spin total RNA extraction kit. RNA concentrations were determined using an Optizen 2120UV spectrophotometer (Mecasys, Daejeon, Korea). RT-PCR assays for target mRNA were performed using a Maxime RT-PCR PreMix kit (iNtRON) according to the manufacturer's protocol. Sequences of PCR primers are summarized in Table 1. PCR was carried out as follows: 5 min at 94°C, 30 cycles of (94°C for 1 min, 56°C for 1 min, and 72°C for 5 min), and a 5-min incubation at 72°C. The glyceraldehyde 3-phosphate dehydrogenase gene was used as an internal control.
ACC, acetyl-coenzyme A carboxylase; aP2, adipocyte protein 2; C/EBPα, CCAAT/enhancer binding protein α; F, forward; FAS, fatty acid synthase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PPARγ, peroxisome proliferator-activated receptor γ; R, reverse; SREBP-1, sterol regulatory element binding protein-1.
Western blot analysis
3T3-L1 preadipocytes (3×104 cells per well) were seeded into six-well plates, cultured in differentiation medium for 7 days, then treated with or without sample, and incubated at 37°C for differentiation. After the medium was discarded, the adipocytes were rinsed twice with ice-cold phosphate-buffered saline and lysed in cell lysis buffer (80 mM Tris-HCl [pH 7.5], 600 mM NaCl, 100 mM EDTA, 100 mM NaF, and 100 mM Na3VO4) containing a protease inhibitor cocktail tablet. Total cell lysates were ultrasonically dispersed for 30 min and centrifuged at 10,000 g at 4°C for 5 min.
For the western immunoblotting, equal amounts of proteins were boiled for 5 min and chilled on ice. Cell lysates (30 μg) were separated by 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electrophoretically transferred to an Immobilon-P transfer membrane. Total protein concentration was determined by the Bradford assay (Bio-Rad). The membranes were blocked by 5% skim milk in Tris-buffered saline with Tween-20 (TBST) (10 mM Tris [pH 7.5], 100 mM NaCl, and 0.1% Tween-20), and then blots were incubated with 1:1000-diluted primary antibody overnight. After being washed with TBST, the membranes were probed with 1:5000-diluted secondary antibody for 1 h and washed with TBST several times. The bound antibody was visualized using an enhanced chemiluminescence detection system (Fuji Film, Tokyo, Japan). The band densities were measured with Science lab version 2.0 software (Fuji Film).
Statistical analysis
Results are expressed as mean±SD values of triplicate experiments. Differences between groups were analyzed by Student's t test, and P values of <.05 were considered statistically significant.
Results
BGE suppresses lipid accumulation in 3T3-L1 cells
We recently reported that BGE reduced the increase of body weight, fat pad weights, and serum and hepatic lipid levels induced by feeding high-fat diets to rats. 17 To understand the mode of action of BGE on 3T3-L1 preadipocyte differentiation, we first performed a cell viability assay using 3T3-L1 cells treated with BGE to determine the optimal concentration of BGE for assay. BGE treatment up to 300 μg/mL showed no significant effect on cell viability (data not shown). To assess the effect of BGE on lipid accumulation during differentiation, we measured TG and FFA levels in lipid droplets of 3T3-L1 preadipocytes treated with different concentrations of BGE (10, 50, and 100 μg/mL) during differentiation. The TG content of intracellular fat droplets was attenuated by BGE treatment in a dose-dependent manner (Fig. 1A). We also found that the total FFA level was remarkably lowered by BGE treatment in a dose-dependent manner (Fig. 1B). BGE treatment with 100 μg/mL resulted in a 31% decrease in FFA in the cells, suggesting that BGE treatment can reduce lipid accumulation in 3T3-L1 cells undergoing differentiation to adipocytes.
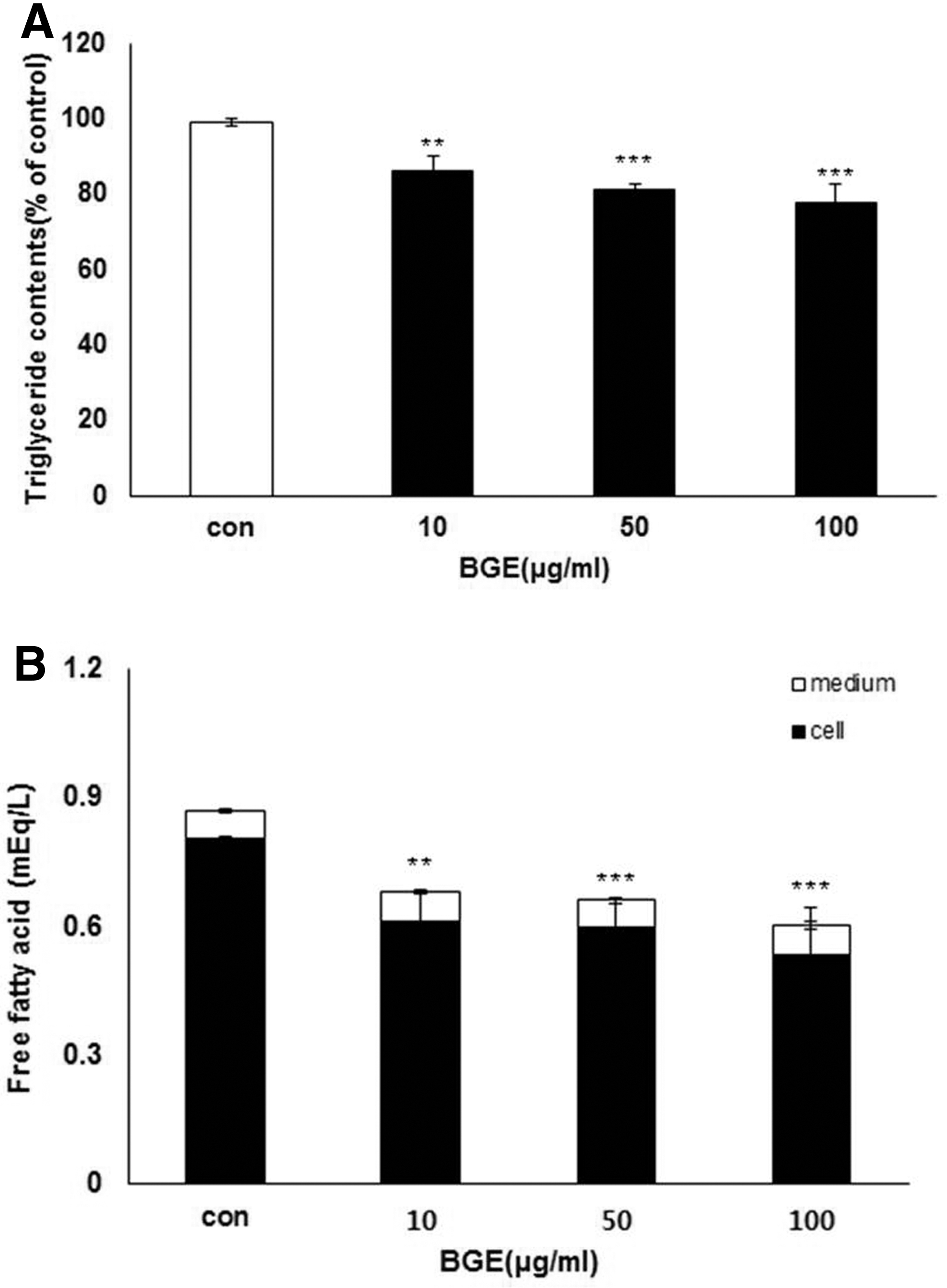
Effects of B. gracilis Miers var. pseudobaselloides Bailey ethanol extract (BGE) on lipid accumulation in 3T3-L1 preadipocytes during differentiation:
Effects of BGE on expression of transcription factors related to 3T3-L1 differentiation
Given that BGE treatment lowers lipid accumulation in 3T3-L1 preadipocytes during differentiation, we next determined whether BGE affects the expression of genes involved in lipid accumulation and differentiation. We assessed differentiation of 3T3-L1 cells in the presence or absence of BGE and measured the expression levels of adipogenic genes such as those for peroxisome proliferator-activated receptor-γ (PPARγ) and CCAAT/enhancer binding protein α (C/EBPα). 20 As expected, BGE treatment reduced both PPARγ and C/EBPα expressions to 37% and 47%, respectively. Moreover, BGE treatment efficiently decreased mRNA levels of C/EBPα even at the lowest concentration of BGE (10 μg/mL) (Fig. 2).
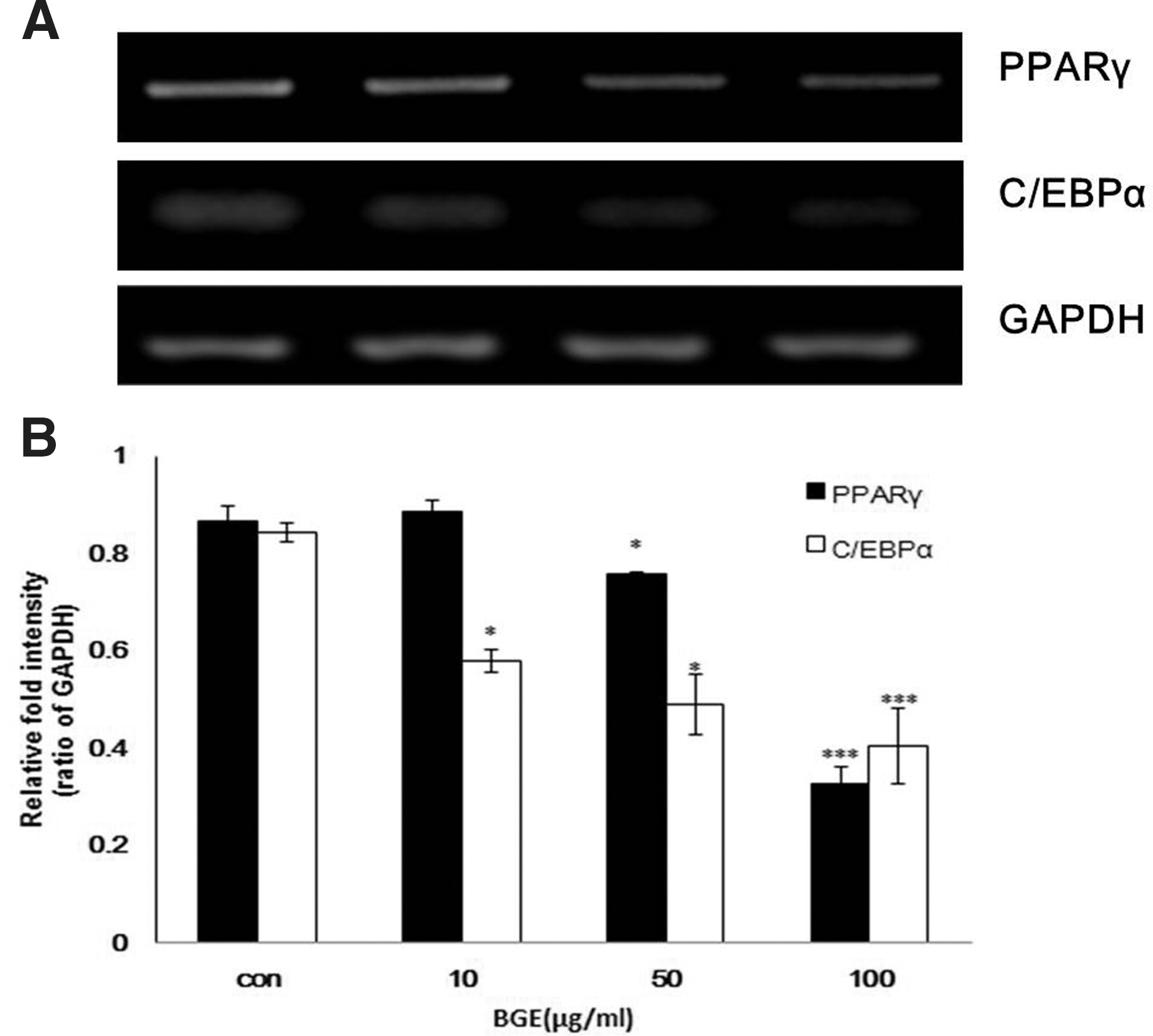
BGE affects the expression of genes related to differentiation. 3T3-L1 cells were treated with BGE during differentiation. Total RNA was extracted at the end of differentiation (Day 6).
BGE inhibits the expression of sterol regulatory element-binding protein-1c and its target genes
Based on our finding that BGE treatment inhibits the levels of cellular TG and FFA, we further investigated whether the hypolipidemic effect of BGE involves regulation of genes related to lipid biosynthesis. Sterol regulatory element-binding proteins (SREBPs) are key transcription factors that bind to the sterol regulatory element DNA sequence and regulate the expression of target genes that are involved in biosynthesis of cholesterol and fatty acids. 20 Fatty acid synthase (FAS) and ACC are transcriptionally regulated by SREBP-1c. 21 The gene for adipocyte protein 2 (aP2) (also known as FABP4), a PPARγ-responsive gene, is also a key regulator of fatty acid synthesis and lipid metabolism. 22 The SREBP-1c gene and its target gene, FAS, were down-regulated in 3T3-L1 cells treated with BGE (Fig. 3). Because SREBP-1c is an essential enzyme for fatty acid synthesis, it is likely that the ability of differentiated 3T3-L1 adipocytes to synthesize fatty acid was impaired by BGE treatment, which is consistent with our finding from the analysis of fatty acid (Fig. 1). Similarly, ACC and aP2 expressions were also affected by BGE treatment. These data suggest that BGE regulates both the SREBP signaling pathway and the fatty acid oxidation pathway at the transcription level.
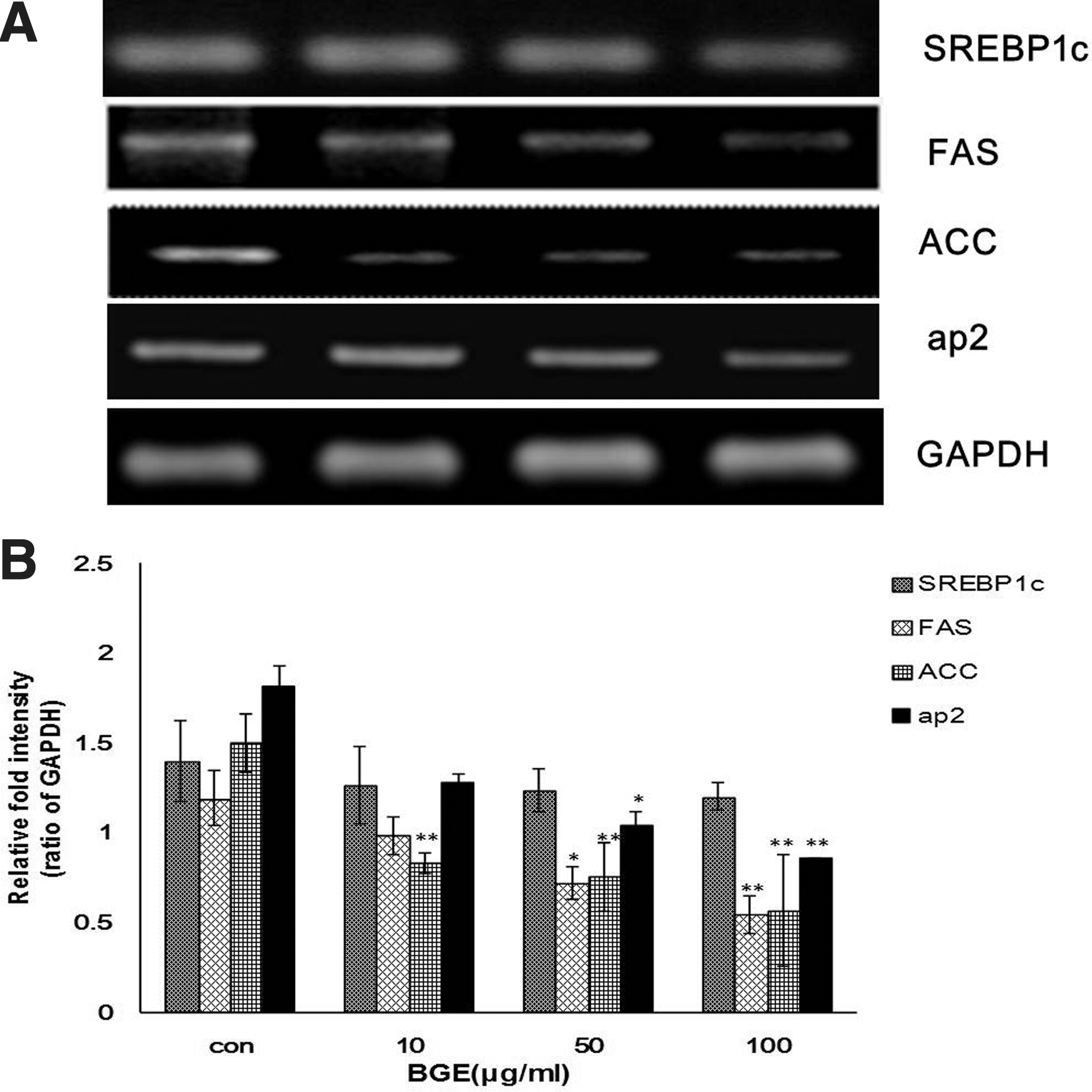
Effect of BGE on expression of genes related to lipid biosynthesis in 3T3-L cells. 3T3-L1 preadipocytes were treated with BGE during differentiation. Total RNA was extracted at the end of differentiation (Day 6).
Activation of the AMPK pathway by BGE treatment
Recently, it has been shown that BGE treatment increases AMPK expression in adipose tissue of high-fat diet-induced obese rats. 17 Here, we sought to determine the effect of BGE on the activity of AMPK, which plays a crucial role in lipid metabolism. 23 As shown in Figure 4A, 3T3-L1 preadipocytes treated with BGE showed increased levels of phosphorylated AMPK and ACC compared with control preparations, suggesting that BGE promotes fatty acid oxidation by activating AMPK.
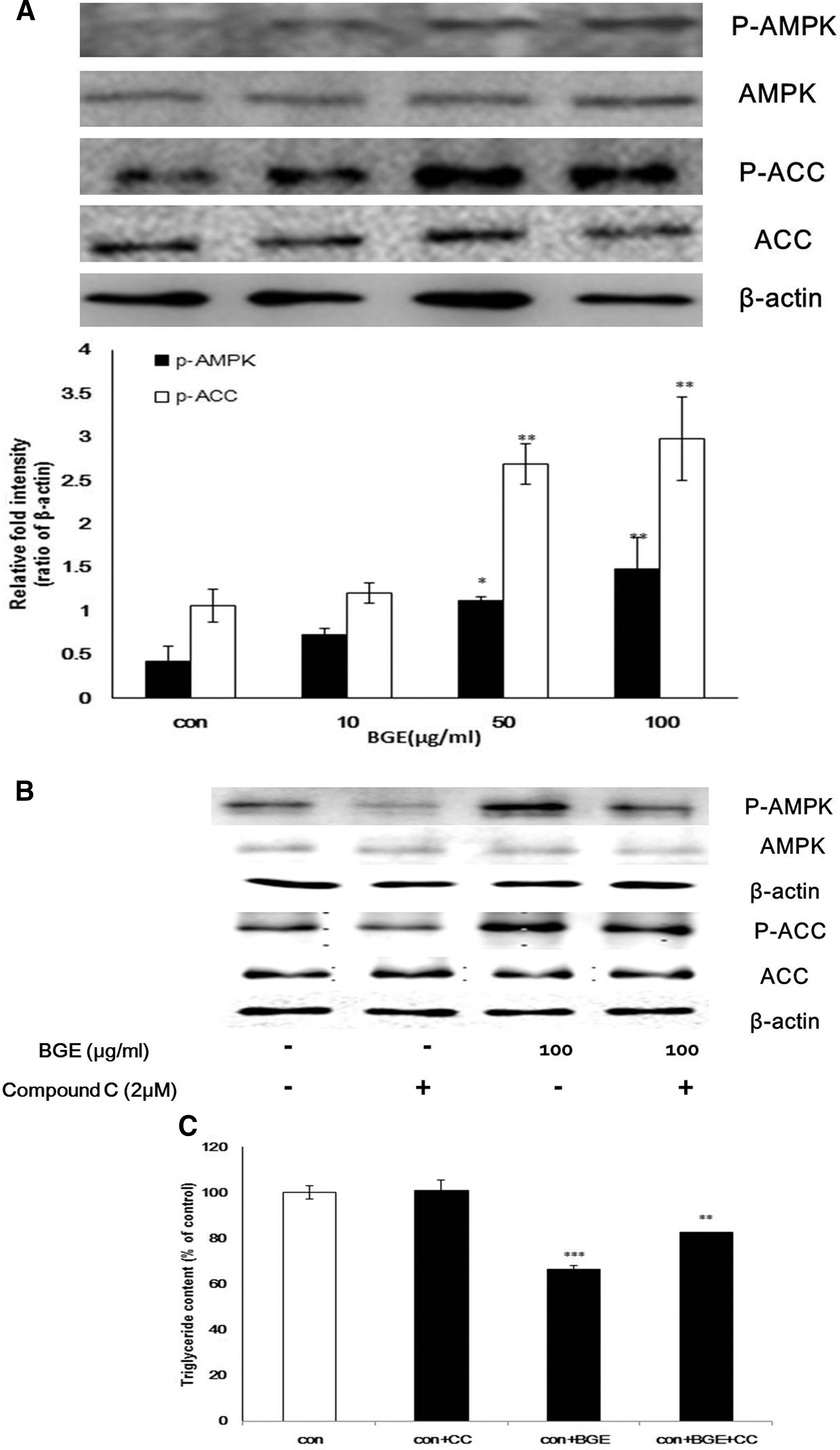
Effect of BGE on phosphorylation of AMP-activated protein kinase (AMPK) and ACC. BGE treatment enhanced phosphorylation in a dose-dependent manner.
To confirm the effect of BGE on phosphorylation of AMPK, we measured phosphorylated AMPK or ACC levels in the presence of compound C, which is an AMPK inhibitor (Fig. 4B). Consistent with the previous report, 17 compound C treatment resulted in decreased AMPK phosphorylation (Fig. 4B, lane 2). It is interesting that BGE treatment resulted in a 159% increase in AMPK phosphorylation (Fig. 4B, lanes 1 and 3), and the effect of BGE on phosphorylation of AMPK was compromised by compound C treatment.
Finally, to test whether BGE inhibition of TG accumulation involves AMPK phosphorylation, lipid contents in fat droplet were measured in BGE-treated cells in the presence or absence of compound C. As shown in Figure 4C, BGE treatment reduced the TG level by 34%, and the effect of BGE on the TG level was attenuated by compound C treatment, suggesting that activation of AMPK might be the molecular mechanism mediating the anti-obesity effects of BGE.
Discussion
BGE has been used for the treatment of diabetes, inflammation, liver diseases, gastric pain, and cough in many countries. 24 Chemical characterization of BGE has been conducted and led to isolation of several sapogenins, including larreagenin A, 3β-hydroxy-30-noroleana-12,19-dien-28-oic acid and its ethyl ester, ursolic acid, 28-ethylhydrogen-3β-hydroxylean-12-ene-28,29-dioate, and 3β-hydroxy-30-noroleana-12,18-dien-29-oicacid. 25 Our recent studies have demonstrated that obese rats treated with BGE for 6 weeks showed decreased body weight gain and serum and hepatic lipid levels compared with rats in the control group. 17 The expression of adipogenesis-related transcription factors, such as PPARγ and C/EBPα, was significantly decreased by BGE administration in obese rats. In addition, BGE treatment decreased blood glucose and lipid levels, indicating that BGE affects lipid metabolism in obese rats.
An initial finding from the current study is that BGE inhibits the lipid accumulation of 3T3-L1 preadipocytes. During adipocyte differentiation, TG and FFA accumulations in BGE-treated 3T3-L1 cells were lower than those in the nontreated group (Fig. 1). Adipocyte differentiation is regulated by transcriptional factors such as C/EBPα and PPARγ, which are able to coordinate the expression of genes related to adipocyte differentiation. 26 These proteins participate in a transcriptional cascade that controls the expression of several genes that are essential for lipid accumulation in adipocytes during the process of differentiation. 26,27 We observed that expression of both PPARγ and C/EBPα was greatly reduced by BGE treatment (Fig. 2), suggesting that the inhibitory effect of BGE on differentiation of preadipocytes might be due to the inhibition of PPARγ and C/EBPα expression.
Fatty acid metabolism is stringently controlled to balance synthesis (e.g., lipogenesis) and degradation (e.g., β-oxidation) in response to various physiological signals. SREBPs, transcription factors that have been associated with lipogenesis, regulate the expression of ACC, FAS, and acyl-CoA synthase. 27 SREBP-1c mRNA expression was decreased by BGE treatment. Consequently, the expression of its downstream target genes such as FAS and ACC was markedly suppressed by BGE (Fig. 3), suggesting that BGE can effectively reduce lipid accumulation by inhibiting lipid synthesis through the SREBP-1c pathway.
AMPK is a “cellular fuel gauge” and acts to simultaneously shut down ATP-consuming biosynthetic processes and facilitate ATP-producing catabolic processes during periods of metabolic stress, 28 leading to rapid changes in the control of fatty acid metabolism. AMPK stimulation of fatty acid metabolism occurs as a result of AMPK phosphorylation and inhibition of ACC. 22,23,26 –28 Our results demonstrate that BGE activates the AMPK pathway to accelerate β-oxidation of FFA. 3T3-L1 cells treated with BGE showed an enhanced phospho-AMPK level compared with the nontreated group (Fig. 4A). Because activity of ACC, which is responsible for producing malonyl-CoA in fatty acid synthesis, is negatively regulated by AMPK, we next examined whether BGE treatment can affect ACC activity. As expected, BGE treatment also increased phosphorylation of ACC (Fig. 4A). Moreover, the increased levels of phosphorylated AMPK and ACC were significantly attenuated by compound C, which is an AMPK inhibitor (Fig. 4B). Thus, it appears that BGE stimulates β-oxidation of FFA by activation of the AMPK pathway.
Previous studies have shown that activation of AMPK in adipocytes leads to decreased fatty acid uptake, decreased TG accumulation, and increased fatty acid oxidation. To confirm the effect of BGE on lipid accumulation via the AMPK pathway in 3T3-L1 preadipocyte differentiation, cells were treated with compound C, and cellular TG levels were monitored. Consistent with western immunoblotting results, the effect of BGE on TG levels was compromised by compound C treatment (Fig. 4C), indicating that activation of AMPK is one of the major molecular mechanisms of BGE in its anti-adipogenic effect.
Recent results showed that the AMPKα subunit interacts with and directly phosphorylates SREBP-1c and SREBP-2, and phosphorylation of SREBP-1c by AMPK represses de novo lipogenesis in hepatocytes. In this previous study, SREBP-1α mRNA was not sensitive to either hyperinsulinemia or S17834, a synthetic polyphenol, and AMPK activation suppressed the accumulation of nuclear SREBP-1c and -2, subsequently repressing expression of their target genes. 29 Thus, it is noteworthy that BGE treatment resulted in a decrease of SREBP1c mRNA level as well as activation of AMPK (Figs. 2B and 3A), suggesting that BGE can efficiently modulate two major regulatory mechanisms of lipid metabolism (the AMPK pathway and the SREBP-1c pathway) not only at the protein level, but also at the transcriptional level.
In conclusion, BGE has anti-adipogenic effects in 3T3-L1 cells by inhibition of lipid synthesis and stimulation of β-oxidation of fatty acid, suggesting that BGE exerts beneficial effects on diabetes.
Footnotes
Acknowledgment
This work was supported by the R&D Program of MKE/KEIT (grant 10033818).
Author Disclosure Statement
No competing financial interests exist.