
Abstract
The Pistacia lentiscus tree gives a resinous exudate called Chios mastic (CM) rich in triterpenoids. CM can be fractionated into acidic and neutral fractions (AF and NF, respectively). Oleanolic acid (OA) is a major triterpenic acid in CM with several antioxidant and anti-inflammatory properties. We have recently shown that CM is beneficial in experimental colitis in the form of powder mixture with inulin, as supplied commercially. However, the bioactive fraction or compound of CM is unidentified. Thus, based on the hypothesis that terpenoids exhibit functional activities via distinguishable pathways, we fractionated CM and applied different fractions or individual OA in experimental colitis. Furthermore, we investigated the mechanism underlying this effect in human colon epithelial cells. CM powder mixture (100 mg/kg of body weight) or the respective CM powder mixture components (i.e., inulin, AF, NF, or OA) were individually administered in trinitrobenzene sulfonic acid-treated rats. Colonic damage was assessed microscopically, and levels of tumor necrosis factor-α (TNF-α), interleukin (IL)-6, IL-8, and intercellular adhesion molecule-1were measured. A model of inflammation in co-cultured human colon epithelial HT29 cells and monocytes/macrophages was established. Lactate dehydrogenase release and levels of TNF-α, IL-8, and nuclear factor-κB (NF-κB) p65 were measured. In vivo, histological amelioration of colitis and significant regulation in inflammation occurred with CM powder mixture, even at the mRNA level. Although no histological improvement was observed, AF and NF reduced levels of inflammatory markers. Inulin was ineffective. In vitro, CM treatment down-regulated IL-8 and NF-κB p65. Neither fractions nor OA was the bioactive component solely. Most probably, the entire CM rather than its individual fractions reduces inflammation via NF-κB regulation.
Introduction
C
With reference to inflammatory bowel diseases (IBD), a pilot trial in Crohn's disease patients showed that CM improves disease activity index, because of regulation of inflammation and oxidative stress. 8,9 We have recently shown that the above-mentioned CM powder mixture has antioxidant and anti-inflammatory effects in trinitrobenzene sulfonic acid (TNBS)-treated colitic rats; when CM powder mixture was administered at a dose of 100 mg/kg of body weight (BW), colon damage was significantly reduced. 10
CM can be fractionated into acidic and neutral fractions (AF and NF, respectively). 6 AF contains triterpenic acids, whereas NF contains triterpenic alcohols and aldehydes. 4,6,11 AF may be responsible for mastic's antimicrobial properties, 6 whereas both AF and NF prevented in vitro oxidation of low-density lipoprotein. 4 However, the bioactive fraction of CM in experimental colitis has not yet been identified. Therefore, in the present study our major objective was to investigate the anti-inflammatory effects of AF and NF, individually, in TNBS colitis. Oleanolic acid (OA), a major triterpenic acid found in CM, has been found to have potent antioxidant, anti-inflammatory, and chemopreventive properties. 12,13 In the present study, the anti-inflammatory properties of OA in experimental colitis were also tested.
In IBD, in the presence of antigen, activated macrophages lead to the production of tumor necrosis factor-α (TNF-α). 14 Additionally, the expression of several cytokines, including TNF-α, interleukin (IL)-6, and IL-8, as well as intercellular adhesion molecule-1 (ICAM-1), is mediated by the activation of nuclear factor-κB (NF-κB). 15 Therefore, an additional objective was to determine the underlying mechanism of CM's anti-inflammatory effects on a co-cultured model of human colon epithelial cells and activated macrophages.
Materials and Methods
Materials
TNBS, phosphate-buffered saline, OA (purity≥97%), and lipopolysaccharides (LPS) were all purchased from Sigma-Aldrich Co. (Munich, Germany). Commercially available CM powder mixture consisting of 60% CM powder and 40% inulin was kindly donated by the Chios Mastic Growers' Association. Human recombinant TNF-α, antibodies, and enzyme-linked immunosorbent assay (ELISA) kits were from R&D Systems (Abingdon, United Kingdom). The ELISA kit for assessment of NF-κB p65 was obtained from Cell Signaling Technology (Danvers, MA, USA). The lactate dehydrogenase (LDH) cytotoxicity assay kit was from ABCAM (Boston, MA, USA). Dulbecco's modified Eagle's medium and RPMI-1640 culture medium were purchased from Biosera (Ringmer, United Kingdom) and Gibco (Carlsbad, CA, USA), respectively. Ficoll-Paque™ Plus was from GE Healthcare. Fetal bovine serum and penicillin–streptomycin were both supplied by PAA Co. (Farnborough, United Kingdom). The total RNA extraction kit, SuperScript™ One-Step reverse transcription–polymerase chain reaction (PCR) with platinum Taq DNA polymerase, and primer pairs were all from Invitrogen Life Technologies (Carlsbad, CA, USA). All solvents were of high-performance liquid chromatography grade and were all from Merck (Darmstadt, Germany).
Fractionation of CM
CM was fractionated into AF and NF according to previously described protocols. 6 In brief, 100 g of CM was dispersed in ethyl acetate (100 mL) and methanol (300 mL). The mixture was stirred and allowed to stand for 48 h. The clear supernatant solution (now soluble in ethanol) was obtained by filtering, and the residual consisting of myrcene (13.3±0.3g) was decanted. The solution was dried under vacuum and diluted in 5% Na2CO3 (200 mL) and diethylether (700 mL), thus being partitioned into an organic phase and an aqueous phase. The organic phase was re-extracted with 5% Na2CO3 (200 mL×3), and the resulting organic phases were dried under vacuum. The powder obtained was the NF (40±4 g). All aqueous phases were acidified with 1 normality HCl (600 mL) and extracted with diethylether (350 mL×3). The resulting organic phases were dried under vacuum, and the powder obtained was the AF (42±3 g).
Gas chromatography/mass spectrometry of OA
Prior to gas chromatography/mass spectrometry analysis the AF was derivatized by addition of 250 μL of N,O-bis(trimethylsilyl) trifluoroacetamide at 70°C for 20 min. An Agilent (Boeblingen, Germany) HP series GC 6890N gas chromatograph coupled with an HP 5973 mass spectrometry detector (electron ionization, 70 eV), a split-splitless injector, and an HP 7683 autosampler was used. The determination of OA was based on a previously described protocol. 16 An aliquot (1 μL) of the silylated extract was injected into the gas chromatograph at a split ratio of 1:20. Separation of sample was achieved using an HP-5 MS capillary column (5% phenyl/95% methyl siloxane; 30 m×0.25 mm×250 μm). Helium was used as a carrier gas at a flow rate of 0.6 mL/min. The injector and transfer line temperatures were set at 280°C and 300°C, respectively. The oven temperature program was as follows: initial temperature, 70°C for 5 min; 70–130°C at 15°C/min; 130–160°C at 4°C/min; held for 15 min; 170–300°C at 10°C/min; held for 15 min. A selective ion monitoring gas chromatography/mass spectrometry method was applied for the detection and quantification of OA. Detection was based on the ±0.05 retention time presence of target and qualifier ions of OA at the predetermined ratios. Target and qualifier ions (T and Q1/Q2, respectively) were set as follows: charge-to-mass ratio, 203, 320, 482. 3-(4-Hydroxyphenyl)-1-propanol (the internal standard) gave values of 206, 191, and 179. Internal standard quantification was performed based on a series of nine standard mixtures of OA, containing the same quantity of internal standard as that of samples. Two control standards (low and high) were analyzed every 10 samples, and if their values were found to be outside of 10% of calibration standards, recalibration was performed. Gas chromatography/mass spectrometry analysis showed that CM powder contained 30±4% OA.
Experimental animals
Adult male Wistar rats (weighing 240–275 g) were supplied by the Hellenic Pasteur Institute (Athens, Greece) and were housed in stainless cages at constant environmental conditions (temperature of 22°C and humidity of 50%) with 12-h day/night cycle. To reduce stress, an acclimatization period to laboratory conditions lasting 2 weeks preceded the experiments. All animals were fed with an isocaloric diet of standard macro- and micronutrient content (78.5% carbohydrates, 2.5% fats, and 19% protein) (Vergerio Mangimi srl, Padova, Italy). Access to food and water was ad libitum. In particular, food consumption was estimated to be approximately 2530 g/day per animal, and water intake was estimated at 25–35 mL/day. The study was approved by the Animal Care Committee and was in agreement with the European Union Act and Greek Law 160, A-64, May 1991.
On Day 1, BW was recorded, and rats were slightly anesthetized by intramuscular administration of a ketamine–xylazine solution (80 mg/kg of BW and 16 mg/kg of BW, respectively). Colitis was then induced applying a routinely established protocol by intracolonic instillation of 25 mg of TNBS dissolved in 0.25 mL of 50% ethanol. 17 All animals were held in the head-down position for about a minute to avoid leakage. On Days 2, 3, and 4, rats were treated with CM powder mixture or its components (i.e., inulin, AF, NF, or OA) once daily, while no change to diet composition was made. In particular, CM powder mixture or individual components were dispersed in 5 mL of water and administered by mouth by oral gavage once daily. On Day 5, experimental animals were anesthetized with ketamine–xylazine (100 mg/kg of BW and 20 mg/kg of BW, respectively) and euthanized with dolethal infused directly in the heart (0.6 mg/kg of BW). Immediately after colon removal (approximately 5 cm starting from rectum), each specimen was longitudinally separated into three parts and stored properly for (1) histological assessment, (2) evaluation of inflammation, and (3) examination of mRNA expression of inflammatory molecules.
Finally, all animals were randomly assigned into 10 groups (eight rats per group) as follows: Group A, control, healthy rats; Group B, TNBS-induced colitic rats; Group C, TNBS-induced colitic rats additionally treated with 100 mg of CM powder mixture/kg of BW/day; Group D, TNBS-induced colitic rats additionally treated with 40 mg of inulin/kg of BW/day (content in 100 mg of CM powder mixture/kg of BW); Group E, TNBS-induced colitic rats additionally treated with 24 mg of AF/kg of BW/day (content in 100 mg of CM powder mixture/kg of BW); Group F, TNBS-induced colitic rats additionally treated with 48 mg of AF/kg of BW/day; Group G, TNBS-induced colitic rats additionally treated with 24 mg of NF/kg of BW/day (content in 100 mg of CM powder mixture/kg of BW); Group H, TNBS-induced colitic rats additionally treated with 48 mg of NF/kg of BW/day; Group I, TNBS-induced colitic rats additionally treated with 7 mg of OA/kg of BW/day (content in 100 mg of CM powder mixture/kg of BW); and Group J, TNBS-induced colitic rats additionally treated with 14 mg of OA/kg of BW/day.
Histological analysis
Colon samples fixed in 10% buffered formalin were embedded in paraffin blocks. Tissue sections 4 μm thick were stained with hematoxylin–eosin and were blindly examined with a light microscope. Criteria of histological scoring were based on a previously described protocol. 10
Tissue sample preparation
Frozen (–80°C) colonic specimens were homogenized in phosphate-buffered saline using an Ultra-Turrax (IKA-Labortechnik, Stauffen, Germany) blender. 10 Samples were centrifuged (3000 g, 15 min, 4°C), and supernatants were collected for protein measurement. 18
Cytotoxicity test of CM or CM components in HT29 cells
CM or CM components (i.e., AF, NF, or OA) were individually tested for cytotoxicity in HT29 human colon epithelial cells. Each individual component was added in culture dissolved in ethanol after removal of myrcene, which eliminates solubility (see Fractionation of CM). For the cytotoxicity test, HT29 cells (1×105 per well) were placed in polystrene 24-well plates (Corning, Corning, NY, USA) with 90% Dulbecco's modified Eagle's medium, 10% fetal bovine serum, and penicillin (100 U/mL)–streptomycin (100 μg/mL) and were incubated for 24 h (37°C, 5% CO2, 95% air) until adherence. Medium was refreshed, and different doses of individual components (final volume, 1 μL) (i.e., CM [0, 80, or 150 ng/mL) or corresponding doses of AF [0, 37, or 69 ng/mL], NF [0, 37, or 69 ng/mL], or OA [0, 11, or 21 ng/mL]) were added for 24 h. Ethanol was tested for cytotoxicity. Supernatants were collected for LDH measurements.
Isolation of monocyte population and differentiation to macrophages
Human peripheral blood mononuclear cells from healthy volunteers (male and female, 23–45 years old) were isolated using Ficoll-Paque and pooled together were resuspended in fresh medium (RPMI-1640 medium supplemented with 10% human serum, 100 U/mL penicillin, and 100 μg/mL streptomycin). The monocyte (Mo) population was obtained by adherence (3×106 peripheral blood mononuclear cells per well) in polystrene 12-well plates. After a 4-h incubation nonadherent T and B cells were removed. Assessment of T cells and Mo population in the adherent cell population (approximately 3×105 per well) was carried out by flow cytometry analysis. In brief, after washing to dispatch nonadherent cells, adherent cells were stained with fluorescein isothiocyanate–conjugated anti-CD3 and phycoerythrin-conjugated anti-CD14. Analysis was performed using a FACScan™ (Becton Dickinson, Franklin Lake, NJ, USA).
Adherent Mo were kept in culture in fresh medium for 6 days until differentiation to macrophages (MΦ) (phenotype checked under the light microscope). MΦ were activated by treatment with LPS (150 ng/mL, 20 h).
Effect of CM or CM components on Mo/MΦ
Supernatants of activated Mo/MΦ were removed, and levels of TNF-α were measured to ensure LPS-induced inflammation. Medium was refreshed (RPMI-1640 medium supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin), and CM or individual components dissolved in ethanol (final volume, 1 μL) (i.e., CM [0, 80, or 150 ng/mL] or corresponding doses of AF [0, 37, or 69 ng/mL], NF [0, 37, or 69 ng/mL], or OA [0, 11, or 21 ng/mL]) were added. After 24 h in culture, supernatants were collected for TNF-α measurement.
Co-culture system
HT29 and Mo/MΦ used in this system were not in contact with each other, enabling crosstalk between them via the soluble factors secreted from each cell population. HT29 cells (1×105 per well) cultured in 90% Dulbecco's modified Eagle's medium, 10% fetal bovine serum, and penicillin (100 U/mL)–streptomycin (100 μg/mL) were incubated in Transwell® permeable supports (pore size, 0.4 μm; Corning) for 24 h until adherence. On the same day, Mo/MΦ were incubated with LPS in 12-well plates. At the end of a 20-h incubation, the medium of activated Mo/MΦ was refreshed. CM or respective components in ethanol were individually added in Transwell permeable supports with cultured HT29 cells. The Transwell permeable supports were immediately submerged into the 12-well plates and incubated for 24 h. All experiments were performed at least five times. Supernatants of HT29 cells were collected and stored at −80°C for measuring LDH and IL-8. NF-κB p65 activation and IL-8 mRNA expression were measured in HT29 cell lysates.
Damage to HT29 monolayers
A standard protocol assay reported for the LDH cytotoxicity kit was performed according to the manufacturer's instructions following 24 h of cell culture. Replicates of untreated cells served as the high control. Three sets of replicates were used for each condition (high control and assays).
NF-κB activation
To assess the activation of NF-κB, we measured levels of the precursor protein NF-κB-p65 in cell lysates in three independent cell cultures, applying a sandwich ELISA kit according to the manufacturer's instructions.
Inflammatory markers
Rat TNF-α, ICAM-1, IL-6, and IL-8 were measured in colonic tissues. Human IL-8 and TNF-α were assessed in three independent cell cultures by quantitative, sandwich ELISA procedures according to the manufacturer's instructions.
Reverse transcription-PCR
Total RNA was extracted from rat colon samples and HT29 cells previously co-cultured with MΦ/MΦ using a total RNA extraction kit according to the manufacturer's instructions. Reverse transcription-PCR was performed using specific oligonucleotide primer pairs: rat IL-8 (forward 5-GGCAGGGATTCACTTCAAGA-3, reverse 5-GCCATCGGTGCAATCTATCT-3), rat ICAM-1 (forward 5-AGGTATCCATCCATCCCACA-3, reverse 5-AGTGTCTCATTGCCACGGAG-3), rat β-actin (forward 5-AATCGTGCGTGACATCAAAG-3, reverse 5-ATGCCACAGGATTCCATACC-3), human IL-8 (forward 5-ATGACTTCCAAGCTGGCCGTG-3, reverse 5-AATTTCTGTGTTGGCGCAGTGTGG-3), and human β-actin (forward 5-TGACGGGGTCATCCA-CACTGTGCCCATCTA-3, reverse 5-CTAGAAGCATTGCGGTGGACGATGGAGGG-3).
The PCR mixtures were denatured at 95°C for 3 min, followed by 30–39 cycles of (denaturation at 94°C for 1 min, annealing at 55–60°C for 45 s, extension at 72°C for 25 s) and final extension at 72°C for 10 min. PCR products were analyzed by electrophoresis in a 2% agarose gel containing ethidium bromide. Cytokine expression was quantified using QuantityOne Analysis software (Bio-Rad, Hercules, CA, USA). Gene expression levels were normalized by using β-actin.
Statistical analysis
Data were processed with SPSS version 17.0 software. Differences between means were tested for statistical significance using one-way analysis of variance and post hoc Tukey's test. Results were expressed as mean±SEM values. Nonparametric data were analyzed with the Kruskal–Wallis test. Results were expressed as median (range). The statistical significance level was set at P<.05.
Results
Histological evaluation
Histological assessment of the colon (Table 1) revealed significant differences between Group A (healthy controls) (Fig. 1A) and Group B (TNBS-induced colitic rats) (Fig. 1B). CM powder mixture-treated animals (Group C) had a lower histological damage score compared with Group B (Fig. 1C), but no differences were observed when corresponding concentrations of inulin (Group D), AF (Group E), NF (Group G), or OA (Group I) were administered (Fig. 1D). Only when doses of CM components were doubled (Groups F, H, and J) did significant changes occur (P<.05) compared with Group B.
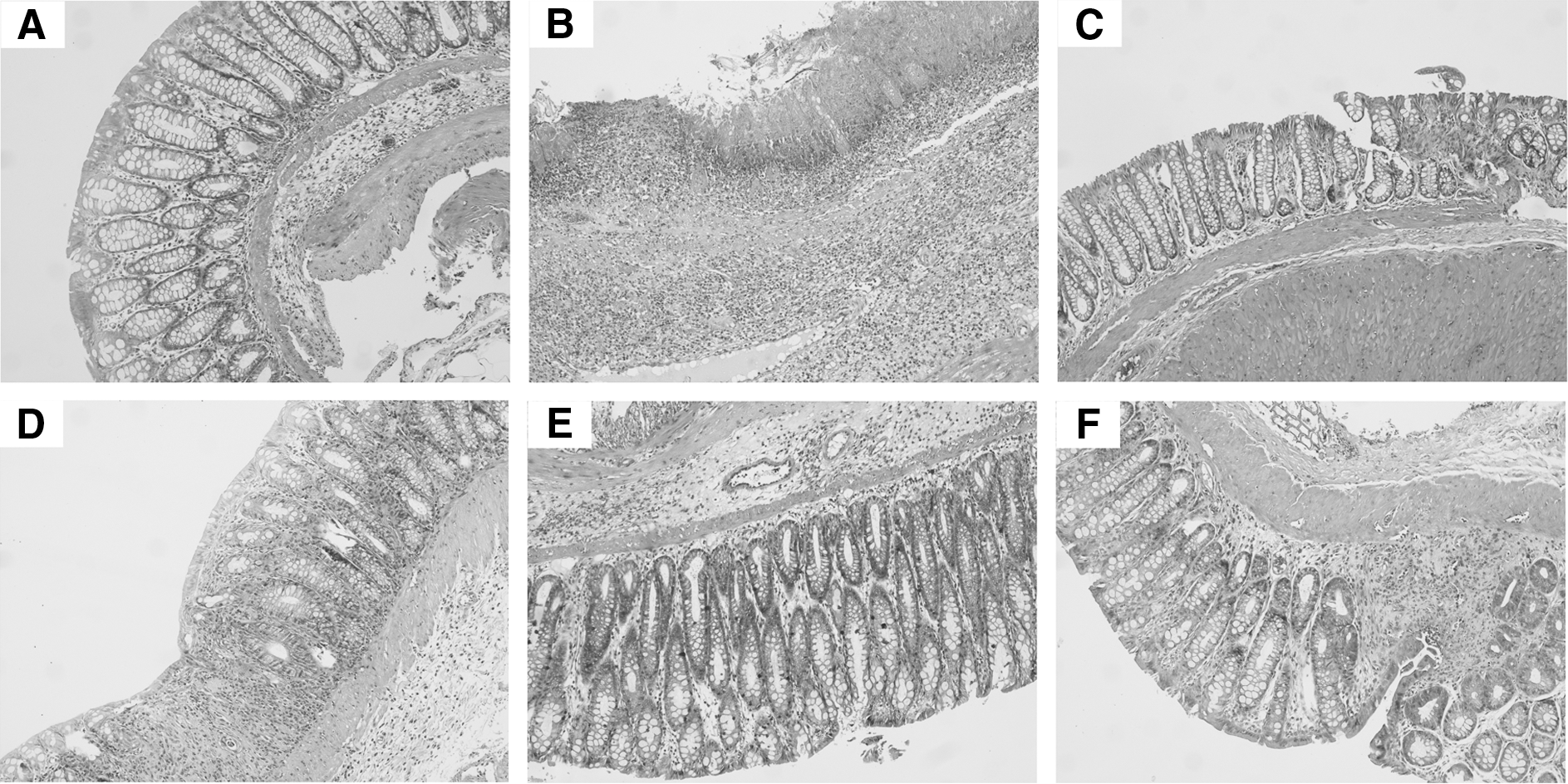
Colonic damage in TNBS-induced colitic rats treated with CM powder mixture or individual components (×100 original magnification):
Grades are expressed as median (range).
The nonparametric Kruskal–Wallis test was applied to ensure differences. Then, the analysis of variance post hoc Tukey's test was applied to check on significant differences between groups. The significance level was set at P<.05. Significant differences occurred between *Groups A and B and **Groups B and C, F, H, and J.
AF, acidic fraction; BW, body weight; CM, Chios mastic; NF, neutral fraction; OA, oleanolic acid; TNBS, trinitrobenzene sulfonic acid.
Inflammatory markers
As shown in Figure 2, the lack of microscopic evidence of colitis in Group A was in agreement with low concentrations of tissue inflammatory markers. Significantly elevated concentrations of TNF-α (445±12 vs. 149±13 pg/mg of protein, P=.001), IL-6 (564±18 vs. 442±9 pg/mg of protein, P=.013), IL-8 (195±10 vs. 102±5 pg/mg of protein, P=.05), and ICAM-1 (12,367±885 vs. 3710±172 pg/mg of protein, P=.001) were observed in Group B compared with Group A. Increases in TNF-α, IL-6, IL-8, and ICAM-1 levels in Group C were significantly suppressed compared with Group B (respectively, 206±7 vs. 445±12 pg/mg of protein, P=.001; 438±6 vs. 564±18 pg/mg of protein, P=.009; 103±8 vs. 195±10 pg/mg of protein, P=.050; and 4222±91 vs. 12,367±885 pg/mg of protein, P=.001). Animals in Group E had significantly lower tissue concentrations of TNF-α (208±21 vs. 445±12 pg/mg of protein, P=.001), IL-6 (450±7 vs. 564±18 pg/mg of protein, P=.026) and ICAM-1 (7515±631 vs. 12,367±885 pg/mg of protein, P=.05), compared with Group B.
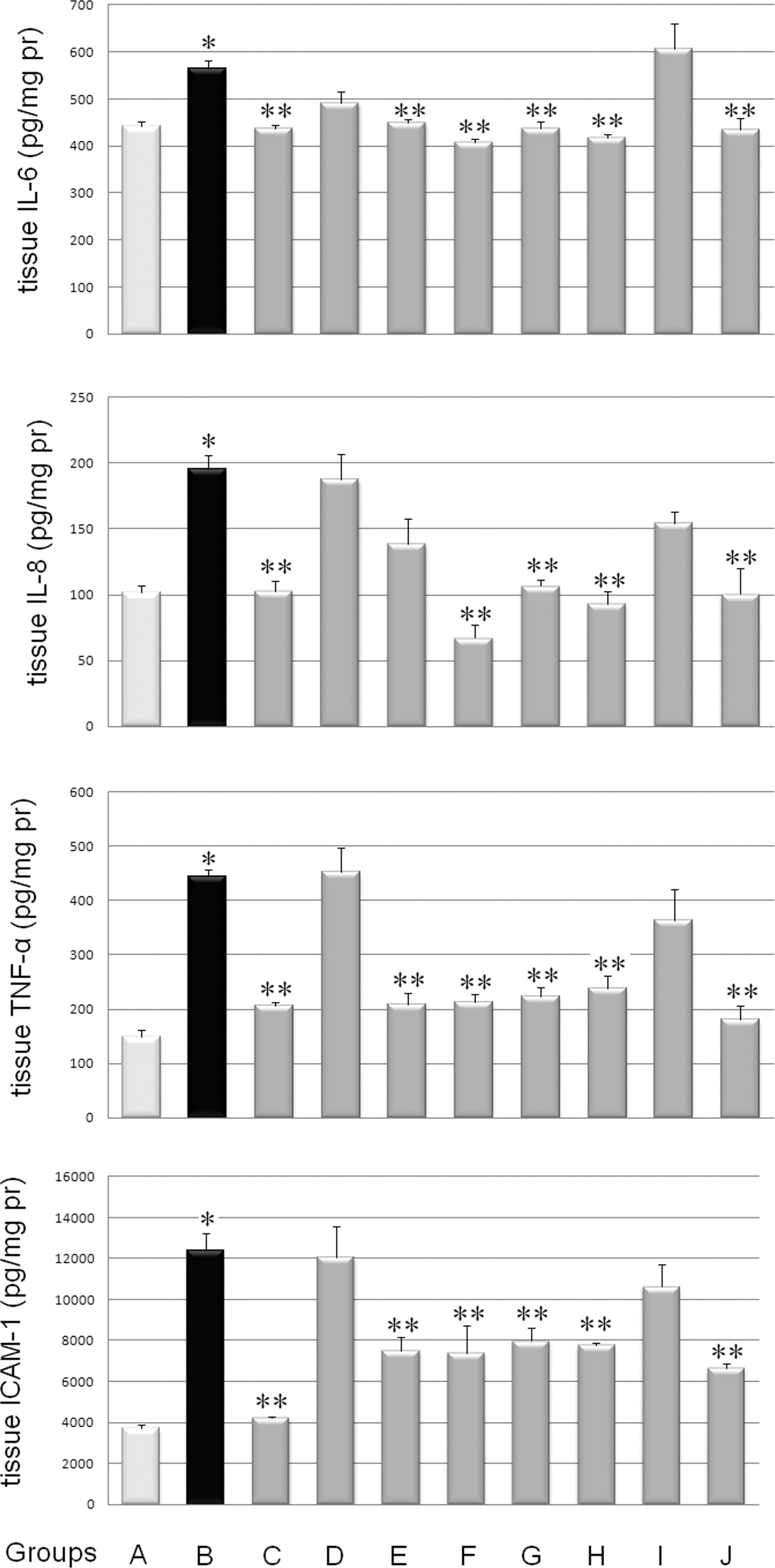
Inflammatory markers in colon of TNBS-induced colitic rats treated with CM powder mixture or individual components: interleukin (IL)-6, IL-8, tumor necrosis factor-α (TNF-α), and intercellular adhesion molecule-1 (ICAM-1). See Table 1 for description of the diferent treatment groups. The significance level of differences was P<.05: *significant differences between Groups A and B; **significant differences between Group B and Groups C–J.
All markers were significantly lower in Group G compared with Group B (TNF-α, 224±8 vs. 445±12 pg/mg of protein, P=.001; IL-8, 438±14 vs. 564±18 pg/mg of protein, P=.009; IL-6, 107±5 vs. 195±10 pg/mg of protein, P=.009; ICAM-1, 7981±618 vs. 12,367±885 pg/mg of protein, P=.017). Levels of TNF-α, IL-6, IL-8, and ICAM-1 measured in Group D or Group I did not differ from the respective value in Group B (Fig. 2). Double doses of CM components administered in Groups F, H, and J, decreased levels of all markers (P<.05), compared with Group B.
TNF-α in Mo/MΦ supernatants
Fluorescence-activated cell sorting analysis showed that >95% of the adherent immune cells expressed CD14 and were the appropriate size for Mo. Less than 5% of cells expressed CD3, indicating that practically no T cells were present. The role of CM in the inflammation cascade was primarily investigated by determining TNF-α levels in Mo/MΦ supernatants (Fig. 3). TNF-α was elevated in LPS-treated Mo/MΦ (38.83±4.13 ng/mL) but did not change when CM was added at 80 and 150 ng/mL (respectively, 34.97±3.30 ng/mL, P=.996; 29.27±3.35 ng/mL, P=.660). Corresponding doses of AF (37 and 69 ng/mL) and NF (37 and 69 ng/mL) also did not affect TNF-α levels: AF, 40.16±4.08 (P=.00) and 35.25±2.69 ng/mL (P=.998), respectively; NF, 40.91±1.71 (P=1.00) and 32.62±3.43 ng/mL (P=.938), respectively. Similarly, corresponding concentrations of OA (11 and 21 ng/mL) had no effect on TNF-α secretion by Mo/MΦ (P=1.00): 40.59±5.52 and 37.58±3.58 ng/mL, respectively.
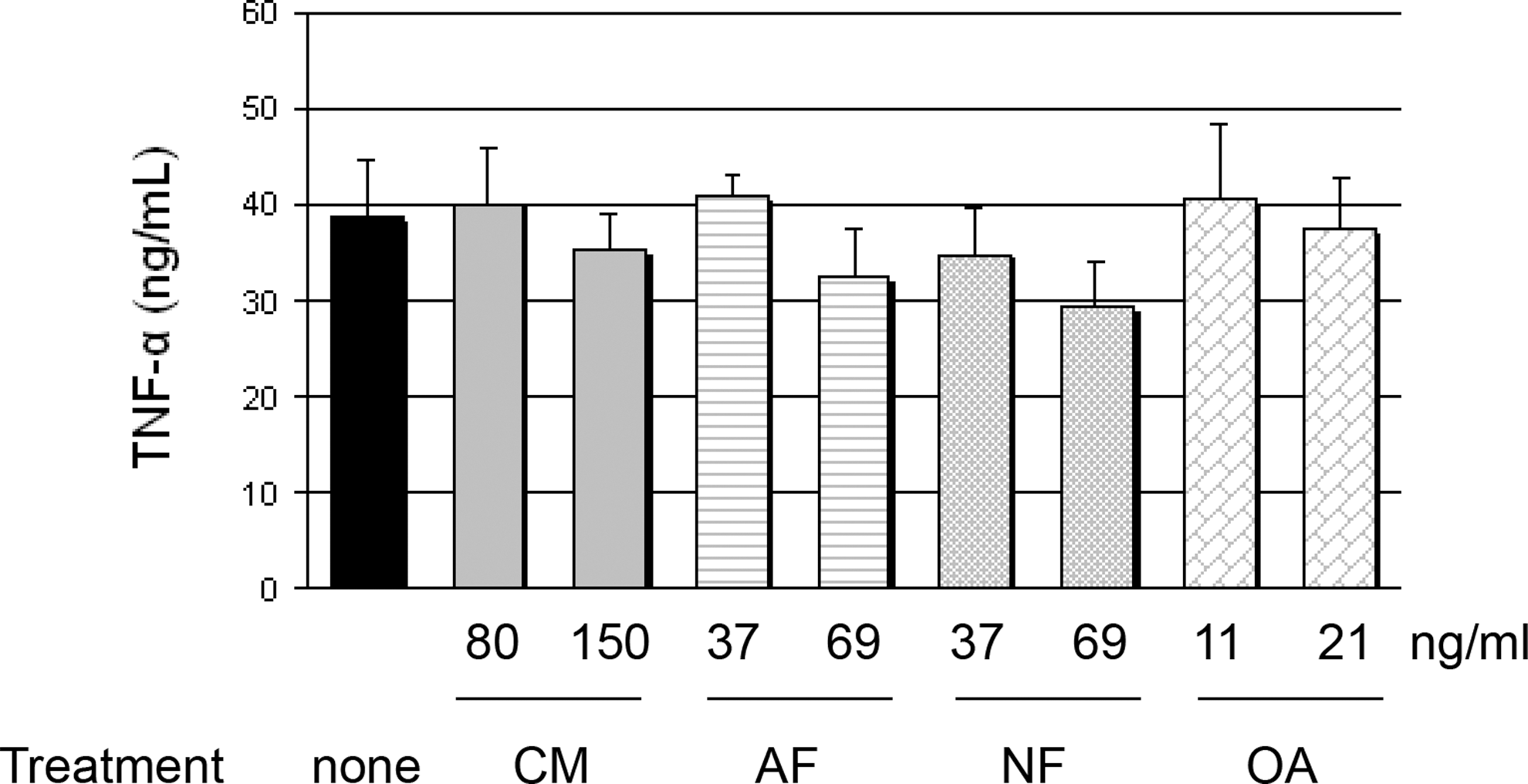
TNF-α in HT29 cells co-cultured with lipopolysaccharides-treated monocytes/macrophages. The significance level of differences was P<.05. There were no significant differences between HT29–monocytes/macrophages and HT29–monocytes/macrophages treated with individual ethanol solutions of CM or corresponding concentrations of AF, NF, or OA.
LDH release
No cytotoxicity was observed when CM or corresponding concentrations of AF, NF, and OA were individually added at 150 ng/mL, 69 mg/mL, 69 ng/mL, and 21 ng/mL, respectively, in HT29 cells. The ethanol applied was not cytotoxic in cell culture. LPS-treated Mo/MΦ induced HT29 cytotoxicity measured by LDH release (53±4%) (Fig. 4). When CM at 80 and 150 ng/mL was tested, LDH release decreased to 15±3% (P=.001) and 6±2% (P=.001), respectively. Corresponding doses of AF (37 and 69 ng/mL) and NF (37 and 69 ng/mL) also down-regulated LDH: AF, 21±3% (P=.001) and 8±3% (P=.001), respectively; NF, 16±4% (P=.001) and 6±3% (P=.001), respectively. Corresponding concentrations of OA (11 and 21 ng/mL) were less effective at reducing LDH: 33±4% (P=.017) and 25±3% (P=.02), respectively.
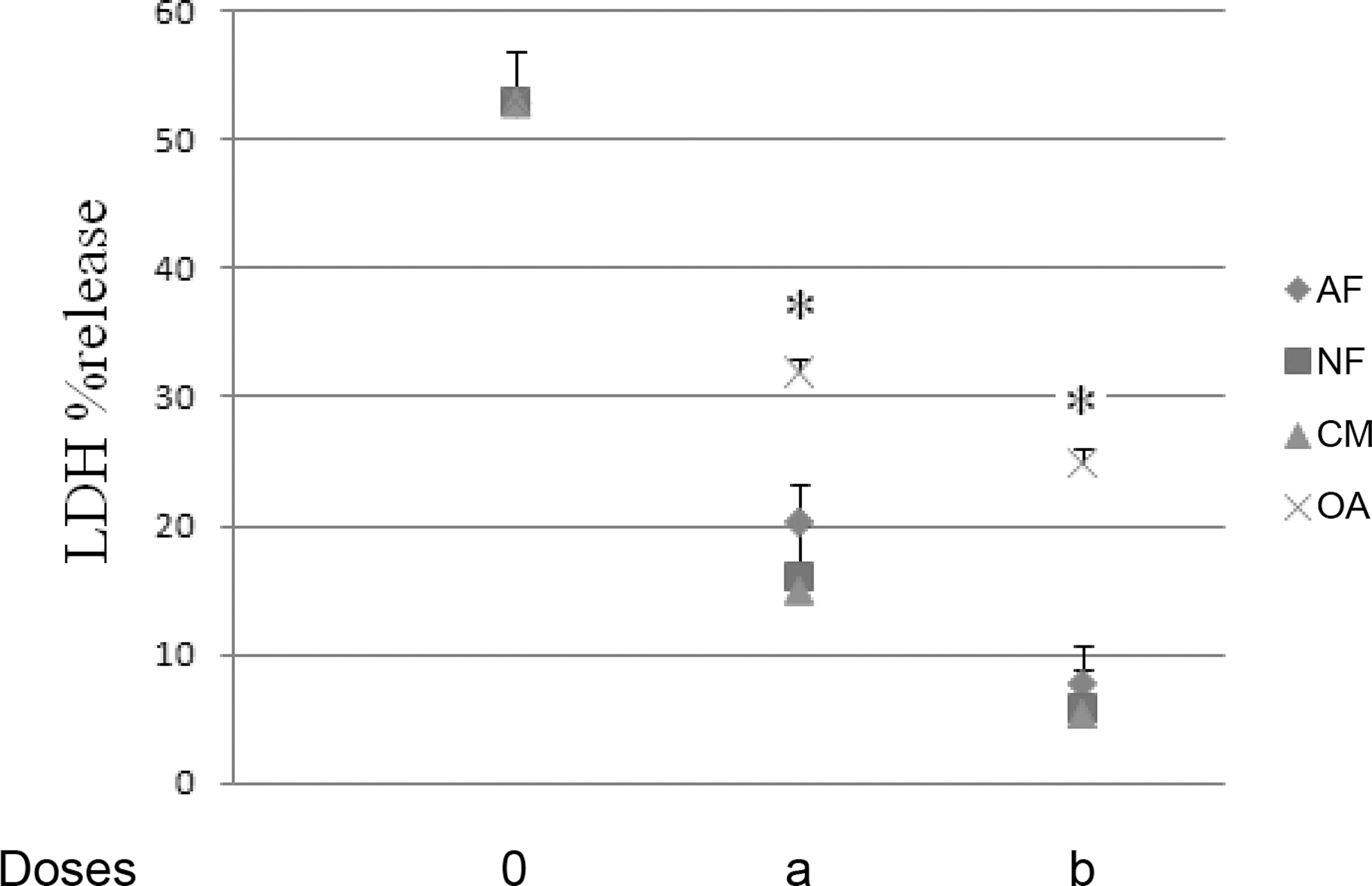
Percentage lactate dehydrogeanse (LDH) release in HT29 cells co-cultured with lipopolysaccharides-treated monocytes/macrophages. The significance level of differences was P<.05: *significant differences between HT29–monocytes/macrophages and HT29–monocytes/macrophages treated with individual ethanol solutions of CM (doses of 0, 80 [a], or 150 [b] ng/mL) or corresponding concentrations of AF (doses of 0, 37 [a], or 69 [b] ng/mL), NF (doses of 0, 37 [a], or 69 [b] ng/mL), or OA (doses of 0, 11 [a], or 21 [b] ng/mL).
IL-8 in HT29 supernatants
In the HT29–Mo/MΦ co-culture system, LPS-treated Mo/MΦ induced IL-8 secretion (Fig. 5A) from HT29 cells (14±1.30 ng/mL) compared with IL-8 levels in HT29 cells prior to addition to the co-culture system (0.0 ng/mL). When CM free of polymers was tested at 80 and 150 ng/mL, the IL-8 concentration was 4±0.40 (P=.001) and 2±0.4 (P=.001) ng/mL, respectively. Corresponding doses of AF (37 and 69 ng/mL) and NF (37 and 69 ng/mL) also reduced IL-8 levels (P=.001): AF, 6±0.65 and 3±0 ng/mL, respectively; NF, 5±0.39 and 3±0.52 ng/mL, respectively. Corresponding concentrations of OA (11 and 21 ng/mL) were less effective at reducing IL-8 levels: 10±0.52 (P=.028) and 8±0.33 ng/mL (P=.02), respectively.
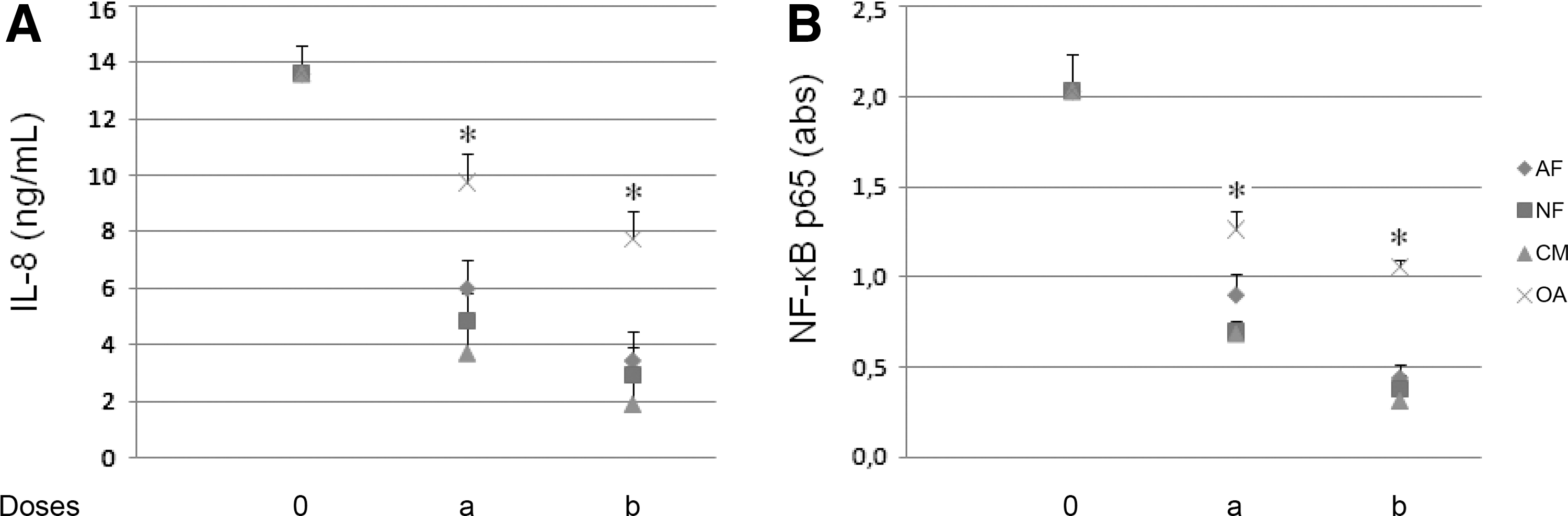
NF-κB p65 in HT29cells
LPS-treated Mo/MΦ induced a significant increase of NF-κB p65 in HT29 cells (2.03±0.21 vs. 0.31±0.06 absorbance units, P=.001) (Fig. 5B). When CM was tested at 80 and 150 ng/mL, NF-κB p65 activation was inhibited, reaching 0.69±0.04 absorbance units (P=.001) and 0.31±0.09 absorbance units (P=.001), respectively. Corresponding doses of AF (37 and 69 ng/mL) and NF (37 and 69 ng/mL) also reduced activation (P=.001): AF, 0.90±0.12 and 0.44±0.08 absorbance units, respectively; NF, 0.69±0.07 and 0.38±0.08 absorbance units, respectively. Corresponding concentrations of OA (11 and 21 ng/mL) were less effective at reducing NF-κB p65: 1.27±0.10 absorbance units (P=.001) and 1.06±0.04 absorbance units (P=.001), respectively.
mRNA expression
Expression of IL-8 and ICAM-1 mRNA in rat colon was up-regulated in Group B compared with Group A (respectively, 92±3% vs. 47±2%, P=.001; 101±3% vs. 42±2%, P=.001) (Fig. 6). Administration of CM powder mixture (100 mg/kg of BW) significantly reduced expression at 44±2% (P=.001) and 58±3% (P=.001) in the respective groups. When corresponding doses of AF (24 mg/kg of BW) and NF (24 mg/kg of BW) were tested, cytokine expression also decreased (P=.001). In particular, IL-8 expression reached 55±1% and 54±3%, respectively, whereas ICAM-1 expression was 72±3% and 76±2%, respectively. OA down-regulated IL-8 (67±3%, P=.001) but not ICAM-1 (105±2%, P=.783) expression.
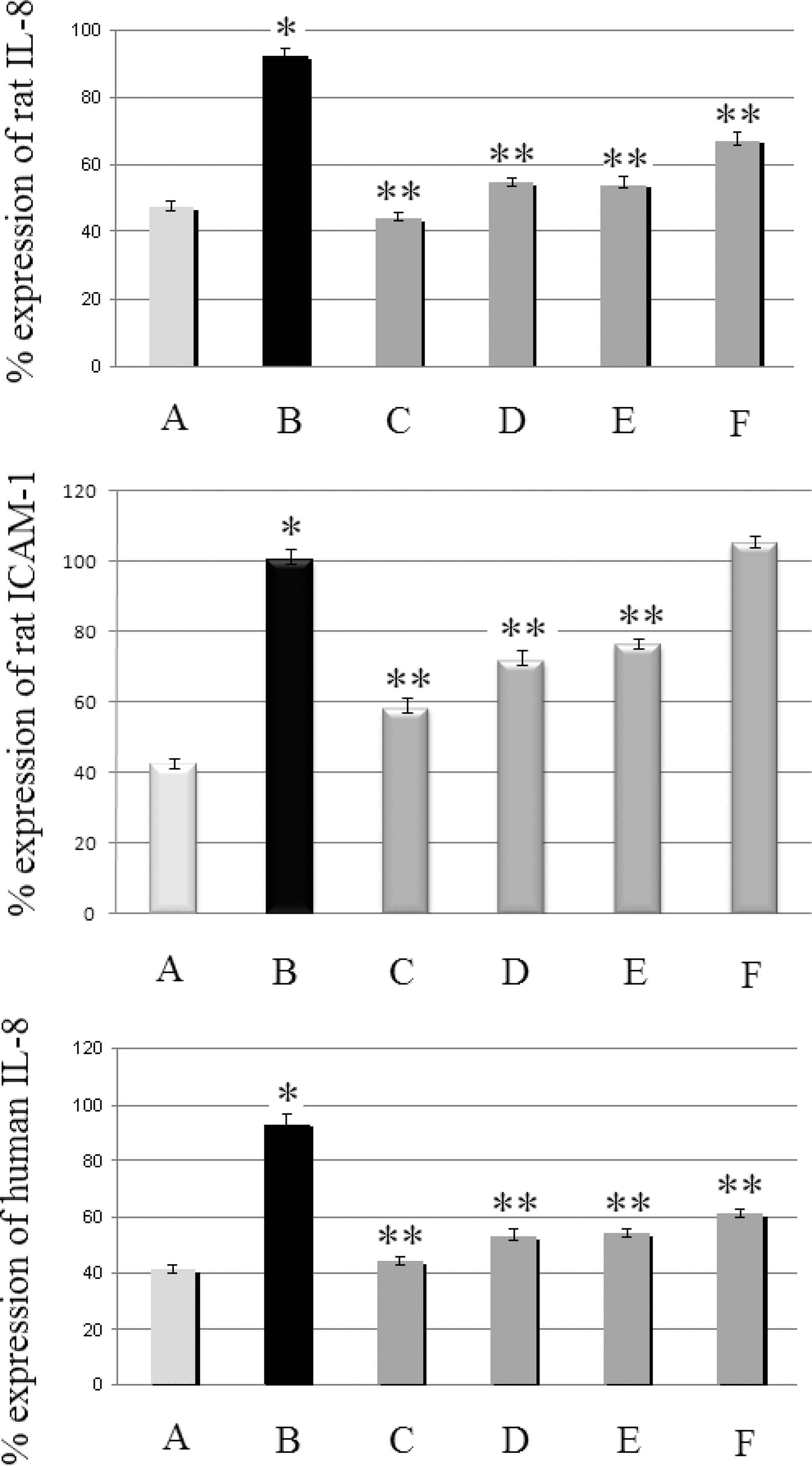
Effect of CM and its respective individual components on IL-8 and ICAM-1 mRNA in rat colon and in human HT29 cells. Levels are expressed as a percentage relative to β-actin. For rat data groups are as follows: A, healthy control group; B, TNBS-induced colitic rats; C, TNBS-induced colitic rats treated with CM powder mixture (100 mg/kg of BW/day); D, TNBS-induced colitic rats treated with AF (24 mg/kg of BW/day); E, TNBS-induced colitic rats treated with NF (24 mg/kg of BW/day); and F, TNBS-induced colitic rats treated with OA (7 mg/kg BW/day). For human HT29 cells groups are as follows: A, untreated HT29 cells; B, HT29 cells co-cultured with monocytes/macrophages; C, monocytes/macrophages-interacted HT29 cells treated with CM (150 ng/mL); D, monocytes/macrophages-interacted HT29 cells treated with AF (69 ng/mL); E, monocytes/macrophages-interacted HT29 cells treated with NF (69 ng/mL); and F, monocytes/macrophages-interacted HT29 cells treated with OA (21 ng/mL). The significance level of differences was P<.05: *significant differences between Groups A and B; **significant differences between Groups B and C–F.
In agreement with rat colon data, IL-8 expression was up-regulated in HT29 cells: 93±4% versus 41±2% (P=.001). CM (150 ng/mL) reduced IL-8 expression at 44±2% (P=.001). When AF (69 ng/mL) and NF (69 ng/mL) were tested, cytokine expression also decreased: 53±3% (P=.001) and 54±2% (P=.001), respectively. OA (21 ng/mL) down-regulated IL-8 expression, reaching 59±4% (P=.001).
Discussion
To our knowledge, this is the first study aimed at investigating the bioactive component of CM powder mixture and the mechanism underlying the anti-inflammatory effect in IBD. Data shown herein indicate that CM powder mixture might be effective against TNBS-induced colitis. Histologically, colonic damage was significantly reduced after a 3-day treatment with CM powder mixture, but no improvement was evident when corresponding doses of AF or NF were tested. Both fractions, however, reduced mRNA and protein of inflammatory markers. OA was ineffective. The lack of any beneficial effect due to inulin alone points to the anti-inflammatory properties of CM contained in the powder mixture. The co-culture of colon epithelial–Mo/MΦ cells indicated that the beneficial effect of CM is probably mediated by NF-κB. The overall beneficial effect of CM powder mixture was instead attributable to both AF and NF. OA, one of the main triterpenoid constituents of CM, exhibited anti-inflammatory activity only when administered at a concentration twofold of that contained in the active CM powder mixture dose (i.e., 100 mg/kg of BW/day). Many studies have shown decreases in inflammatory markers in applying natural products or individual components in TNBS colitis. 19 –21 The decrease was in most cases attributed to a synergistic activity of several phytochemicals rather than to the activity of an individual component. 22,23
CM powder mixture consisting of 60% CM powder (obtained from the P. lentiscus tree) and 40% inulin improved colonic damage and ameliorated inflammation. This is in agreement with our previous work in TNBS-induced colitic rats. 10 The effect of inulin administered in our TNBS model was negligible (Fig. 2). Apart from its prebiotic activity, inulin is also used to avoid coagulation and improve product spreadability. In the case of CM, inulin is applied for spreadability and powder stability. Moreover, in this study, inulin facilitated the suspension of CM powder in water prior to oral gavage. In some studies inulin has been found effective against IBD; 24 –26 however, doses were much higher than that used here (40 mg/kg vs. ∼200 mg/kg).
On the whole, recruitment of activated MΦ, along with neutrophils and dendritic cells into the lamina propria induces the local immune response. TNF-α produced by nonlymphoid cells, mostly MΦ, is essential for IBD development. 27 Depletion of MΦ in an IL-10-deficient mouse colitis model inhibited the development of colitis, which would otherwise occur because of cytokine secretion by MΦ. 28 MΦ generate reactive oxygen species that trigger inflammation and tissue damage and also increase epithelial disruption. In our in vitro study, the effect of CM on Mo/MΦ is negligible (Fig. 3). In our in vivo study, CM mixture powder reduced TNF-α levels, indicating that immune cells other than MΦ regulate TNF-α (e.g., natural killer cells, lymphocytes).
The NF-κB p65 level was elevated in HT29 cells cultured with Mo/MΦ, whereas a decrease occurred when CM or individual components were added. The p65 subunit activation is important in IBD as it is highly activated in mucosal biopsy specimens of IBD patients. 29 Activated NF-κB was found in biopsy specimens from inflamed mucosa, but not in uninflamed mucosa by using the p65 monoclonal antibody. 30 We hypothesize that the underlying mechanism engages a decrease in NF-κB activation and down-regulation of the genes encoding for IL-8 and ICAM-1. Subsequently, a decrease in inflammatory cell recruitment occurs.
Down-regulation of NF-κB results in expression of several anti-apoptotic genes, inhibition of tissue damage, and protection of the intestinal barrier. CM reduced LDH release from the HT29 cell monolayer, consequently affecting cell membrane permeability. Cytoplasmic LDH outflow is a useful marker of membrane permeability and integrity. 31 The integral barrier of epithelia is essential for physiological homeostasis. Furthermore, physiological cell turnover via apoptosis is critical. The integrity of the mucosa is vital for nutrient and for fluid absorption and secretion. Nevertheless, increased permeability of the intestinal epithelium fluctuates with disease severity. Although during spontaneous cell apoptosis and division no disruption occurs in the epithelium, severe cytokine-induced cell damage may create distinct barrier defects and may down-regulate mucosal integrity. 27 Thus, the significance of CM's effect in HT29 cytolysis may be critical.
Supporting nutrition is an important area of treating patients with IBD. Encompassing the use of natural products in nutrition might become an effective means to inhibit intestinal damage.
Footnotes
Acknowledgments
This work was supported by the Chios Mastic Growers' Association and the Greek State Scholarships Foundation.
Author Disclosure Statement
A.P. is an employee of ELPEN-Pharmaceuticals Co. Inc. A.G., A.C.K., A.L., G.A., E.P., and N.K.A. declare no competing financial interests exist.