
Abstract
In the present study, we injected pregnant mice at Day 7 of gestation with cadmium chloride (CC) (1.5 mg/kg) intraperitoneally and determined its effect on the frequency of fetal malformations at Day 17 of pregnancy. On the same day, we also determined the level of micronucleated polychromatic erythrocytes (MNPEs) and of micronucleated normochromatic erythrocytes (MNNEs) in blood cells of both the mothers and their fetuses. A significant increase in the number of malformations was found, mainly exencephaly, micrognathia, ablephary, microphthalmia, and clubfoot, as well as a significant increase in the amount of MNPEs and MNNEs. In addition, pregnant mice were administered grapefruit juice (GJ) orally from Days 0 to 17 of the experiment (from 200 to 800 μL/g) to evaluate the potential of the juice in preventing the damage induced by CC. We found a dose-dependent decrease in the number of visceral and skeletal malformations, as well as in the number of MNPEs and MNNEs, in both the mothers and their fetuses. Furthermore, we determined the level of DNA oxidation by measuring levels of the adduct 8-hydroxy-2′-deoxyguanosine, and we found a significant increase in such level induced by CC; in contrast, there was a significant decrease when we added GJ. Therefore, the observed teratogenic and genotoxic protection can probably be related with the antioxidant potential of GJ.
Introduction
T
With respect to cadmium's teratogenic potential, studies have concluded that the results obtained depend on the species/strain tested, on the dose used, and on the time of administration. 4,5 In the mouse in particular, administration of cadmium before gestation can suppress the implantation of the zygote, and in gestating mice it is known to produce a large range of fetal malformations. 6,7 With regard to its genotoxic potential, cadmium was initially considered a weak genotoxic agent, yet subsequent reports have demonstrated that it may alter DNA replication and repair, besides inducing mutations as well as chromosomal aberrations, micronuclei (MNs), and sister chromatid exchanges. 8,9 However, its effect on occupational exposure is controversial, as positive and inconclusive studies have reported. 10 These data support the relevance of finding measures to avoid or reduce its toxicity. With this purpose in mind, various authors have tested antioxidants, such as vitamins (C and E), quercetin, naringenin, mangiferin, and gingseng, 11 –14 to improve the functioning of organisms exposed to cadmium.
In this context, the antioxidant potential of grapefruit has been demonstrated through methods such as the 1,1-diphenyl-2-picrylhydrazyl radical scavenging test, β-carotene bleaching, and oxygen radical scavenging capacity assay. 15 Furthermore, reports have shown that administering grapefruit juice (GJ) to mice treated with various mutagens reduced the genotoxic effect. This has been observed in regard to aflatoxin B1, daunorubicin, ifosfamide, and benzo[a]pyrene by quantifying MNs, sister chromatid exchanges, or comet tail increase. 16 –18 In some of these studies, the authors have suggested that the trapping of reactive oxygen species by GJ may explain its protective effect.
Based on the aforementioned information and in the absence of antiteratogenic studies of GJ, as well as of data regarding its antigenotoxic potential on cadmium-induced damage in the mouse fetus, we established the aims of the present report: to determine whether the juice reduced the frequency of malformations induced by the metal in mice, as well as the number of MNs induced by it in pregnant mice and in their fetuses. We also determined the effect of GJ on the level of 8-hydroxy-2′-deoxyguanosine (8-oxo-dG) induced by cadmium, so as to analyze whether the prevention of DNA-oxidative damage could have a role in our results. Besides, the present approach may contribute to the understanding of the correlation between genotoxic and teratogenic effects produced by cadmium.
Materials and Methods
Chemicals and animals
Alcian blue, alizarin red, and cadmium chloride (CC) were purchased from Sigma-Aldrich Chemicals Co. (St. Louis, MO, USA). Urea, ethanol, acetone, and potassium hydroxide were obtained from J.T. Baker Chemicals (Mexico City, Mexico). Acetic acid, picric acid, Giemsa stain, and formaldehyde were purchased from Merck S.A. (Mexico City). The HT 8-oxo-dG enzyme-linked immunosorbent assay kit for the detection and quantification of 8-oxo-dG in serum samples was obtained from Trevigen, Inc. (Gaithersburg, MD, USA). Male and female mice (ICR strain) weighing 28–32 g were used for the experiment. Animals were kept in stainless steel cages at a temperature of 24±1°C, 50±10% relative humidity, and a 12-h light/dark cycle with light switched on at 9:00 a.m. Mice had free access to food (rodent laboratory chow 5001; Purina, St. Louis). The experimental study was approved by the Committee of Ethics and Biosecurity of the National School of Biological Sciences (Mexico City).
Experimental design
Initially, one male mouse was caged with four nulliparous females from 6:00 to 9:00 a.m. The presence of a vaginal plug was considered evidence of coitus and was also taken to designate gestational Day 0. Pregnant mice were randomly assigned to eight groups with eight animals each and administered treatments as follows: a negative control group was orally given purified water daily, from Days 0 to 17 of the study; another group was orally administered GJ (32.8 μL/g) daily, during the same period; a positive control group was intraperitoneally injected with CC at 1.5 mg/kg (dissolved in water) at Day 7 of pregnancy; and five other groups were administered GJ daily (4.1, 8.2, 16.4, 24.6, and 32.8 μL/g, respectively) plus CC at 1.5 mg/kg at Day 7 of pregnancy. The doses of the tested chemicals were selected according to preliminary assays and previous reports. 17,19 Maternal body weight was monitored on Days 0, 7, 11, 15, and 17 of the experiment.
Determination of fetal malformations
Animals were cervically dislocated on Day 17 of gestation. All fetuses were removed, weighed, and inspected for external malformations. One-third of the fetuses were fixed in 96% ethanol to apply the method of Peters 20 for determining skeletal malformations. The remaining fetuses were fixed in Bouin's solution for 4 weeks; subsequently, the method of Wilson 21 was applied to determine the types and the number of visceral malformations. For the statistical analysis, we considered the litter as the experimental unit. The results concerning visceral and skeletal malformations were statistically analyzed with the χ2 test followed by Fisher's exact test. The remaining variables were assessed by one-way analysis of variance, followed by the Student–Newman–Keuls test.
Determination of MNs
A blood sample by the tail of each of the eight pregnant mice was taken and smeared on clean slides. With regard to the fetuses, a neck incision was made to four fetuses per mother, blood was collected in a capillary tube, and a drop was smeared on each of two clean slides. The remaining blood was centrifuged at 2000 g for 15 min to separate the serum, which was kept at −70°C so as to later determine the DNA oxidative damage.
The slides of both the mothers and their fetuses were fixed in methanol for 3 min, washed, dried, and stained with 10% Giemsa solution made in phosphate-buffered saline for 12 min. To determine the genotoxic effect induced by CC, as well as the protection produced by GJ, we recorded in each organism the number of micronucleated polychromatic erythrocytes (MNPEs) in 1000 polychromatic erythrocytes and the frequency of micronucleated normochromatic erythrocytes (MNNEs) in 1000 normochromatic erythrocytes. Statistical analysis of the obtained results was made with an analysis of variance followed by the Tukey–Kramer test.
Quantification of 8-oxo-dG
The assay was carried out in the serum of the fetuses from mice treated as indicated in Experimental design. The determinations were made in five groups with six individuals each: a group administered purified water, a group administered CC (1.5 mg/kg), another group treated with GJ at 32.8 μL/g, and two other groups treated with CC at 1.5 mg/kg plus GJ at 16.4 and 32.8 μL/g, respectively. The calibration curve and the effect of the tested compounds were determined in duplicate. For the quantification, we followed the instructions of the HT 8-oxo-dG enzyme-linked immunosorbent assay kit. In the immunoassay, the 8-oxo-dG monoclonal antibody binds competitively to 8-oxo-dG immobilized on precoated wells and in solution. Antibody bound to 8-oxo-dG in the sample is washed away, whereas antibody bound to 8-oxo-dG attached to the well is retained. Detection is performed with horseradish peroxidase conjugate and tetramethylbenzidine substrate. The absorbance was measured in a microplate reader at 450 nm, and the intensity of the yellow color was found to be inversely proportional to the concentration of 8-oxo-dG. The basis to calculate the average concentration of 8-oxo-dG for each sample was the standard curve. For the calculation, we used the worksheet provided by the manufacturer. 22
Results
Teratogenic/antiteratogenic study
Results concerning maternal toxicity are shown in Table 1. A comparison between the CC-treated mice and the control group showed no significant differences with respect to maternal weight gain, placental diameter, placental weight, and fetus weight, as well as regarding the number of implantations, resorptions, and live fetuses. However, as to the fetal crown–rump length, we found a significant decrease in mice administered CC and in those treated with CC plus the two low doses of GJ (4.1 and 8.2 μL/g).
The determinations were made in the fetuses of eight pregnant mice per group. Data are mean±SEM values.
Statistically significant difference by one-way analysis of variance and Student–Newman–Keuls tests, P<.05 with respect to *control (water-treated) mice or †cadmium chloride (CC)-treated mice.
GJ, grapefruit juice.
Results concerning the type and the frequency of malformations observed at term are indicated in Table 2. We detected no malformations in the control mice or in animals treated only with GJ; in contrast, mice administered CC showed a large number of abnormalities, with exencephaly being the most prevalent (65.9%). It is interesting that the administration of GJ to CC-treated animals induced a dose-dependent inhibition in the rate of all types of malformations. The highest dose of the juice tested presented the best preventive effect, with a mean reduction of 32.7% for exencephaly, 32.6% for micrognathia, 54.5% for ablephary, 100.0% for microphthalmia, and 56.0% for clubfoot, compared with the corresponding values in the CC-treated group.
The determinations were made in fetuses of eight pregnant mice per group. Values in parentheses represent percentages.
Statistically significant difference by χ2 test followed by Fisher's exact test, P<.05 with respect to *control (water-treated) mice or †CC-treated mice.
Table 3 shows the results obtained with respect to visceral malformations. The control and the GJ-treated groups exhibited no induced abnormalities, and the CC treatment was able to increase the number of the four types of observed anomalies. However, it is also clear that all doses of GJ tested had a significant protective effect.
The determinations were made in the fetuses of eight pregnant mice per group. Values in parentheses represent percentages.
Statistically significant difference by χ2 test followed by Fisher's exact test, P<.05 with respect to *control (water-treated) mice or †CC-treated mice.
Table 4 presents the results obtained with regard to skeletal malformations. In this case, as in the previous determination, the control and the GJ-treated groups exhibited no abnormalities, contrary to the significant elevation induced by CC. These refer to delayed ossification in the head, sternebrae, and vertebrae, as well as in fused ribs and vertebrae. The administration of GJ to CC-treated mice generally gave rise to a dose-dependent reduction in the number of malformations. The strongest preventive effect was detected with the high dose of GJ (32.8 μL/g).
The determinations were made in the fetuses of eight pregnant mice per group. Values in parentheses represent percentages.
Statistically significant difference by χ2 test followed by Fisher's exact test, P<.05 with respect to *control (water-treated) mice or †CC-treated mice.
Genotoxic/antigenotoxic study
Results of the MN assay in pregnant mice and their fetuses are presented in Table 5. In adult mice administered GJ we found a similar range of MNPEs with respect to the control group. The mean frequency for these two groups was 4.35 MNs/1000 polychromatic erythrocytes. In contrast, animals administered only CC showed an increase of more than five times such level, whereas mice treated with both CC and the juice exhibited a dose-dependent reduction in the number of MNPEs that was statistically significant compared with the level found in CC-treated animals. The best protection (72.28%) was determined with the highest dose tested. With respect to the level of MNNEs, we observed a behavior similar to that described earlier, although with a higher amount of MNs, which is a logical finding considering that MNs recorded in normochromatic erythrocytes include those that have been accumulated with time. The administration of CC increased the MNNE control level more than six times, and the combined administration of GJ plus CC produced a dose-dependent decrease in the number of MNs induced by CC. The best result (an inhibition of 53.15%) was obtained with the highest dose tested.
The determinations were made in eight pregnant mice per group and in four fetuses per group. Micronucleated polychromatic erythrocytes (MNPEs) were determined in 1000 polychromatic erythrocytes per mice, and micronucleated normochromatic erythrocytes (MNNEs) were determined in 1000 normochromic erythrocytes per mice. Data are mean±SEM values.
Statistically significant difference by one-way analysis of variance and Student–Newman–Keuls tests, P<.05 with respect to *control (water-treated) mice or †CC-treated mice.
With regard to the fetuses, we found a mean amount of 4.44% and 26.58% for MNPEs and MNNEs, respectively, when we considered the values of the control and the group administered with GJ. The administration of CC to mice increased more than four and five times the level of MNPEs and MNNEs, respectively, and when GJ was administered together with CC, a dose-dependent decrease was determined for both types of erythrocytes.
Quantification of oxidative DNA damage in serum samples
Figure 1 shows the mean concentration of 8-oxo-dG detected in the serum of fetuses. Control mice revealed a low value of DNA oxidation (0.07 ng/mL); a similar value was found in animals administered GJ at 32.8 μL/g, a result that indicated no DNA oxidative potential by GJ. However, animals treated with CC had an increase of 152% with respect to the control level; in contrast, mice administered CC plus GJ showed a mean decrease of 45% with respect to the value of CC-treated mice. The stronger inhibition corresponded to the group treated with the highest dose of GJ.
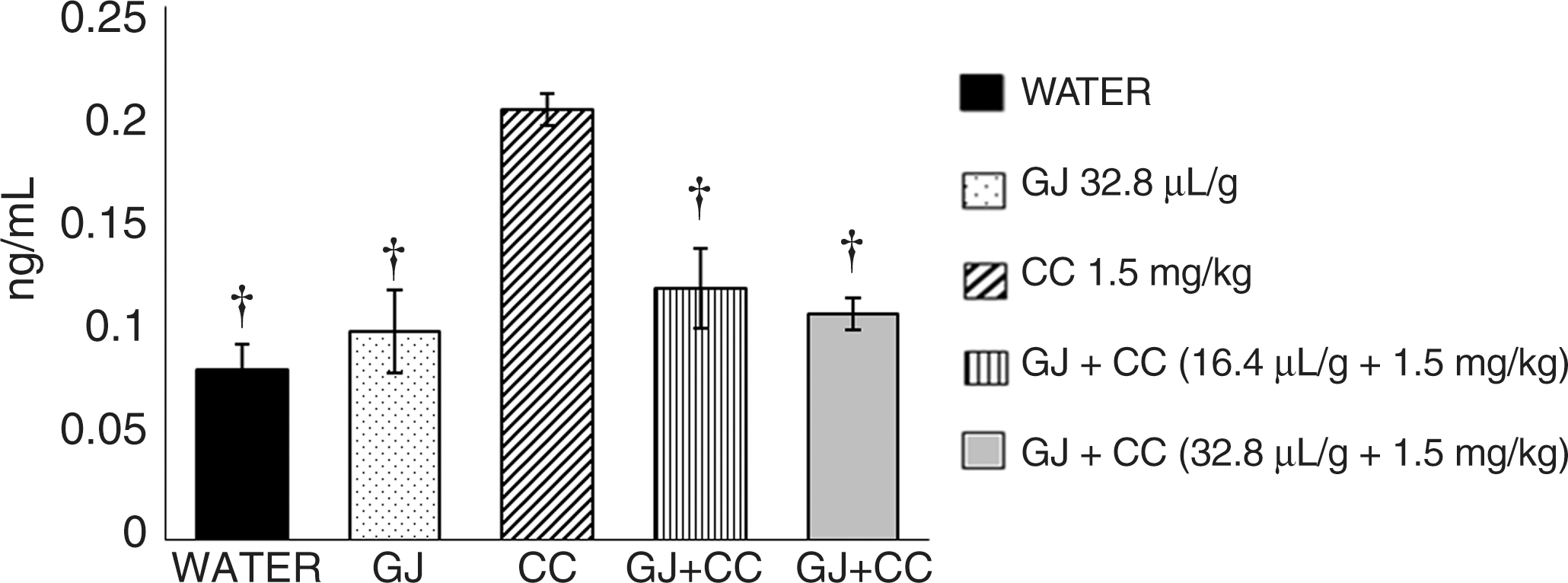
Effect of GJ on level of 8-hydroxy-2-deoxyguanosine (8-oxo-dG) induced by CC in the serum of mouse fetuses. 8-Oxo-dG was measured in fetal serum samples by means of the HT 8-oxo-dG enzyme-linked immunosorbent assay kit. Bars represent mean±SEM values of five groups with six mice each. The calibration curve and the effect of the tested compounds were determined in duplicate. †Statistically significant difference with respect to the value of CC-treated mice by one-way analysis of variance and Student–Newman–Keuls tests, P<.05.
Discussion
In the present report, we determined the deleterious effect of CC in pregnant mice and their embryos. In light of the fact that no significant decrease in maternal weight was observed, our data suggest a direct toxic effect by CC on the fetus, although toxicity through other mechanisms cannot be discounted, such as by altering the supply of nutrients to embryos because of metabolic changes. 19,23 Our results on the type of malformations detected in the CC-treated group agree with previous studies, although the reported frequencies differ among authors 6,7 because of variations in experimental conditions or because certain cells in the embryo can be exceptionally sensitive to the agent. 5,6 In agreement with its teratogenic potential, cadmium is also known to produce other effects in the reproductive tissue, such as necrosis and apoptosis, failure in the progression of oocyte development from primary to secondary stage, failure of ovulation, ineffective pick up of the oocyte by the tubal cilia, and retardation of trophoblastic outgrowth and development, in addition to the inhibition of progression to the blastocyst stage. 24
With regard to MNs, our present results on pregnant mice agree with the strong clastogenic potential of the agent shown in various in vitro and in vivo models. 9,24 –27 However, no information on the matter had been previously reported in the mouse fetus. In such cells, our results also showed a strong induction of MNs, although lower than that observed in the mothers, a result that differs from those reported for other compounds, 28 suggesting that experimental conditions and the tested agent were relevant factors for the observed results.
A transplacental genotoxic effect had been previously reported in umbilical cord and liver fetal cells, but with respect to mutagens such as benzene, vinblastine, benzo[a]pyrene, or verapamil. 28,29 A few reports have shown some relationship with metals, although without a clear conclusion. In Central Serbia, the authors found a significant increase in the number of MNs in the lymphocytes of newborns, which was attributed to the environmental presence of polychlorinated biphenyls and heavy metals, including chromium and nickel. 30 However, another study done in mice reported no effect in mothers or in their fetuses after administration of chromium(VI). 31 On the other hand, maternal smoking has been associated with increased rates of spontaneous abortion, perinatal mortality, congenital defects, and low birth weight. 32 This is interesting in light of the fact that cadmium is a metal found in tobacco smoke, as well as in the urine of pregnant women smokers; 33 furthermore, the suggested explanation for the mentioned effects has been that tobacco-specific metabolites can cross the placental barrier and affect the DNA of the fetus. In this respect, although conflicting results have been observed with regard to MN induction by tobacco smoke, a positive result was found in cord blood samples taken from mothers who smoked during pregnancy, in comparison with the level of those who were nonsmokers. 29 Moreover, presence of the metal has been associated with oxidative stress determined in infertile subjects, as well as with apoptotic increase, and with germ cell density decrease found in human fetal ovaries. 34
The intracellular production of reactive oxygen species has been considered one of the main causes of cadmium toxicity, an effect that has been demonstrated through the induction of lipid peroxidation, protein carbonylation, mitochondrial alterations, and a decrease in endogenous and exogenous antioxidants. 35 Therefore, finding antioxidant agents for preventing or reducing cadmium toxicity is mandatory. In regard to this strategy, chelation therapy or the effect of divalent cations has been tested. 36,37 More interesting, however, has been the positive effect observed with vitamin C or quercetin, 11 which are constituents of grapefruit and possess antigenotoxic properties as well. Results obtained with these latter agents suggest that the protective effect that we found with GJ can be related to its widely known antioxidant capacity; this suggestion is strongly supported by the sharp effect exhibited by GJ in reducing the amount of the adduct 8-oxo-dG, which is a biomarker for DNA oxidative damage, known to adversely affect the DNA function via anomalies in gene transcription or because of mutational mechanisms. 38
In summary, our study determined that CC induced a significant genotoxic effect in pregnant mice, as well as genotoxicity and teratogenicity in their products. It also demonstrated a transplacental preventive capacity of GJ, as well as in the pregnant mice against cadmium, an effect that was due, at least partially, to the action of its antioxidant constituents. Thus, our results add new information concerning the beneficial attributes of GJ.
Footnotes
Author Disclosure Statement
No competing financial interests exist.