
Abstract
The antioxidant and antimicrobial properties of two edible mushrooms (Pleurotus ostreatus PBS281009 and Coprinus comatus M8102) were evaluated in this study. The mycelium ethanolic extract obtained in the presence of four different nitrogen sources was investigated with regard to their production of phenolic compounds, β-carotene, lycopene, and their antimicrobial and antioxidant properties. Significant differences in phenol and β-carotene concentrations in mycelia grown in different nitrogen sources were observed. Corn extract proved to be the most appropriate source for increasing bioactive properties, leading to the lowest EC50 and minimum inhibitory concentration values in the P. ostreatus PBS281009 species, while peptone was the most suitable for the C. comatus M8102 species. The antimicrobial capacity was screened against Gram-positive and Gram-negative bacteria and against two species of Candida. These results indicate that the mycelia from these two edible mushrooms can be used as nutraceuticals and functional products.
Introduction
M
Usually, the production of Pleurotus ostreatus and Coprinus comatus edible fungi takes an extended period of time. Reducing the time for obtaining mycelium by submerged cultivation represents the most advantageous alternative for undertaking in vitro studies to determine the therapeutic effects due to the content of polyphenolic compounds, β-carotene, and even lycopene. 4 –6 P. ostreatus is a significant source of dietary fiber and valuable nutrients. 6,7 This mushroom contains biologically active compounds with different therapeutic properties, including the ability to stimulate the immune system, to decrease cholesterol, and to prevent high blood pressure, atherosclerosis, hypoglycemic effects, and antithrombotic effects. 6,8 The mycelium of C. comatus was found to contain ergothioneine, a thiol compound with antioxidant properties. 9 The extracts of mycelium and the isolated polysaccharides confirmed these and also additional, significant anticancer properties. This mushroom is also known for its hypoglycemic effect. 4,10
Due to the high demand for bioactive compounds, the development of efficient, submerged cultivation processes have become an important research direction. In this regard, finding new, efficient sources of carbon and nitrogen represents an important role. The effects of using plant extracts as carbon sources (from corn flour, bran, or other plant materials) for cultivating edible mushroom mycelia is well known. 11 In contrast, the nitrogen source has a greater influence on the production of mycelium in a submerged medium; its influence is related to the amount of compounds with antioxidant (less lycopene) and antimicrobial effects. 2,3
This study was performed to evaluate the antioxidant activities of P. ostreatus PBS281009 and C. comatus M8102 freeze-dried mycelia extracts. Influence of various nitrogen sources on the amount of several antioxidant components (total phenols, total flavonoids, ascorbic acid, α-tocopherol [TOC], β-carotene, and lycopene) of total ethanolic extracts, as well as on its reducing power, superoxide, nitric oxide, and hydroxyl radicals scavenging activity were examined using by typical biochemical methods. Variation with the same nitrogen sources, of antioxidant activity against β-carotene-linoleic acid, of ethanolic extracts, was also investigated. Differences in submerted growth rates of the mycelium, as well as the antimicrobial potential of the extract were also expected to occur.
Materials and Methods
Culture and storage condition
The pure culture of C. comatus M8102 species was originally obtained from the Mycelia bvba, Belgium. The mushroom P. ostreatus PBS281009 was isolated from the Baneasa forest, Romania, and was authenticated by Dr. D. Pelinescu, Faculty of Biology, University of Bucharest, Romania. The mycelia were maintained on potato dextrose agar (PDA) at 4°C. The microorganisms were subcultured at regular intervals (45 days) to maintain viability.
Media preparation and fermentation condition
The fungi were initially grown on a PDA medium for 10 days at 25°C. The inoculum was prepared by growing mushrooms on a LabTech rotary shaker at 150 g for 5 days and at 25°C in 500 mL Erlenmeyer flasks containing 250 mL of the following synthetic medium (per liter): 6.0 g glucose, 100.0 g malt extract, 20.0 g yeast extract, 1.0 g KH2PO4, and 0.5 g MgSO4×7H2O. The medium was adjusted to pH 5.5 with 0.2 M NaOH. 12
Submerged fermentation was carried out in 1000-mL Erlenmeyer flasks, containing 700 mL of liquid medium (KH2PO4 0.2%, CaSO4 0.5%, MgSO4 0.05%, and Na2HPO4 0.01% in 5% extract solution of corn flour) and was performed with four different nitrogen sources: corn extract (dry substance 40%), peptone, yeast extract, and ammonium sulfate. The nitrogen sources were added as 10 g/L. The inoculated flasks were maintained on a LabTech rotary shaker at 150 rpm and 25°C. After 7 days of growth, the mycelium was recovered from the liquid medium by centrifugation at 4000 g for 15 min. Next, the obtained mycelia were washed three times with distilled water and freeze-dried in an Alpha 1-2 LD freeze-dryer in the absence of a cryoprotective agent. 2,13
Preparation of mushroom extract
The extract was obtained by ethanol extraction of freeze-dried mushroom mycelia. Ethanol extraction of the mycelia was accomplished by stirring at 150 rpm for 24 h at 20°C, with a ratio of 1 g of freeze-dried biomass per 10 mL solvent. The broth was centrifuged at 3000 g for 15 min and the supernatant was filtered using Whatman no. 1 filter paper. The ethanol extract was freeze-dried. The freeze-dried extract was then redissolved in 80% ethanol (v/v) to yield solutions containing 1.0, 2.5, 5.0, 10.0, 15.0, and 20.0 mg of extract per mL.
Antimicrobial activity
In vitro antimicrobial susceptibility tests were performed using a panel of microorganisms from the collection of the Faculty of Biotechnology, Bucharest, Romania: Gram-positive bacteria (Listeria innocua CMGB 218, Bacillus cereus CMGB 215, and Staphylococcus aureus ATCC 6588), Gram-negative bacteria (Escherichia coli CBAB 2 and Pseudomonas aeruginosa ATCC 15442), and yeast (Candida albicans ATCC 20231 and Candida sp. ICCF15). The yeast and bacteria were maintained in 20% glycerol and maintained at −80°C.
Determination of minimum inhibitory concentration
The standard agar dilution protocol with doubling dilution was used. The extract was incorporated into nutrient agar at concentrations ranging from 0.39 mg/mL to 25 mg/mL. A control without the extract was also prepared. Ten microliters of each test organism, previously diluted to 106 colony-forming units per mL, was used to inoculate the plates. These were incubated at 37°C for 24 h in the first instance, and for another 24 h before the growth was observed and recorded. The minimum inhibitory concentrations (MICs) of the extract for each test microorganism were considered the lowest concentrations without growth. 14
1,1-Diphenyl-2-picrylhydrazyl radical scavenging activity of freeze-dried mushroom extracts
The reaction mixture contained 50 μL of test samples (or 80% EtOH as a blank) and 5 mL of a 0.04 g/100 mL solution of 1,1-diphenyl-2-picrylhydrazyl (DPPH) in ethanol. Discoloration was measured at 517 nm after incubation for 30 min. DPPH radical concentration was calculated using the following equation: DPPH scavenging effect (%)=(A0 – AP)/A0×100%, where A0 was the absorbance of the control and AP was the absorbance in the sample. The actual decrease in absorption induced by the test compounds was compared with the positive controls. Ascorbic acid was used for comparison and as a positive control. The extract concentration providing 50% of free radical scavenging activity (RSA), known as EC50, was calculated from the graph of the RSA percentage against extract concentration. 15,16
Antioxidant activity by β-carotene-linoleic acid
The antioxidant activity was determined with slight modifications of the procedure previously described by Sokmen et al. 17 Briefly, 0.5 mg β-carotene was dissolved in 1 mL of chloroform and 25 μL linoleic acid and 200 mg Tween 40 were added. Chloroform was removed using a rotary vacuum evaporator and 100 mL of distilled water saturated with oxygen was added with vigorous shaking. A volume of 2.5 mL of this reaction mixture was dispensed into test tubes and 350 μL of various concentrations of the extracts were added. The absorbance was immediately measured at 490 nm. The reaction mixture was incubated at 50°C for 2 h and the absorbance was measured again. The same procedure was repeated with a synthetic antioxidant (α-TOC and butylated hydroxytoluene [BHT]) and a control. Inhibition ratio of linoleic acid oxidation was calculated for the test samples and for the synthetic antioxidants. 18
Reducing power
Reducing power was determined according to the method described by Gulcin et al. 19 Each extract (in 2.5 mL of ethanol) was mixed with 2.5 mL of 200 mM sodium phosphate buffer (pH 6.6) and 2.5 mL of 1% potassium ferricyanide, and the mixture was incubated at 50°C for 20 min. Next, 2.5 mL of 10% trichloroacetic acid was added, and the mixture was centrifuged at 3000 g for 10 min. The upper layer (2.5 mL) was mixed with 2.5 mL of deionized water and 0.5 mL of 0.1% ferric chloride. Finally, the absorbance was measured at 700 nm and compared to a blank. The EC50 value is the effective concentration at which the absorbance was 0.5. BHT and TOC were used as positive controls. 6,16
Determination of antioxidant component
Determination of total phenolic content
The extract sample (0.50 mL) was added to a 1:10 dilution of Folin–Ciocalteu reagent (2.5 mL). After 4 min, a saturated sodium carbonate solution (approximately 75 g/L, 2 mL) was added. The absorbance of the reaction mixture was measured at 760 nm after incubation for 2 h at room temperature. Gallic acid was used as a reference standard, and the results were expressed as milligrams of gallic acid equivalent (mg GAE) per gram of extract. 20
Determination of total flavonoids
Flavonoid content measurements were conducted according to the simple and reliable method described by Jia et al. 21 Fifty microliters of ethanolic extract was mixed with 700 μL of deionized water and 37 μL of 5% NaNO2. After a 5-min incubation at room temperature, 75 μL of 10% AlCl3 was added, followed by 250 μL of 1 M NaOH for the next 6 min. After shaking, the mixture was centrifuged (5000 g, room temperature, 15 min), and the absorbance of the supernatant was read at 515 nm against a blank. Quercetin was used as a standard. 1
Determination of β-carotene and lycopene
For β-carotene and lycopene determination, the dried ethanolic extract (100 mg) was vigorously shaken with 10 mL of an acetone–hexane mixture (4:6) for 1 min and filtered through Whatman No. 4 filter paper. The absorbance of the filtrate was measured at 453, 505, and 663 nm. β-carotene and lycopene content were calculated according to the following equations: lycopene (mg/100 mL)=−0.0458×A663+0.372×A505−0.0806×A453; β-carotene (mg/100 mL)=0.216×A663−0.304×A505+0.452×A453. The results are expressed as mg of carotenoid/g of extract. 22
Determination of α-TOC
The content was determined spectrophotometrically according to the method of Kivcak and Akay. The α-TOC content in the extracts was calculated from the regression equation of the standard curve. 23
Determination of ascorbic acid
A sample of extract (20 mg) was extracted with 10 mL of 1% metaphosphoric acid for 45 min at room temperature and filtered through Whatman No. 4 filter paper. One milliliter of filtrates was mixed with 9 mL 2,6-dichloroindophenol and the absorbance was measured at 515 nm. The ascorbic acid content in the extracts was calculated from the regression equation of the standard curve. 24
Superoxide RSA of freeze-dried extracts
The scavenging activity on superoxide radicals was evaluated according to the methods described by Chou et al. 25 The reaction mixture contained the same volume of 120 μM phenazine methosulfate, 936 μM NADH, freeze-dried extract, and 300 μM NBT, in a total volume of 1 mL of 100 mM phosphate buffer (pH 7.4). After 5 min of incubation at ambient temperature, absorbance of the resulting solution was measured at 560 nm. The superoxide radical activity was calculated as: scavenging effect (%)=[1−(absorbance of sample/absorbance of control)]×100%. Ascorbic acid was used for comparison. The EC50 value (mg extract/mL) is the effective concentration at which hydroxyl radicals were scavenged by 50%. 26
Hydroxyl radical scavenging of freeze-dried extracts
Hydroxyl radical scavenging was assayed as described by Varshneya et al. 27 with a slight modification. The assay is based on the quantification of the degradation product of 2-deoxyribose by condensation with thiobarbituric acid (TBA). Hydroxyl radicals were generated using the Fe3+–ascorbate–EDTA–H2O2 system (the Fenton reaction). The reaction mixture contained, in a final volume of 1 mL: 2-deoxy-2-ribose (2.8 mM), KH2PO4-KOH buffer (20 mM, pH 7.4), FeCl3 (100 μM), EDTA (100 μM), H2O2 (1.0 mM), ascorbic acid (100 μM), and various concentrations (0–200 μg/mL) of the freeze-dried extracts or reference compound. After incubation for 1 h at 37°C, 0.5 mL of the reaction mixture was added to 1 mL 2.8% trichloroacetic acid, then 1 mL of 1% aqueous TBA was added, and the mixture was incubated at 90°C for 15 min to develop the color. After cooling, the absorbance was measured at 532 nm against an appropriate blank solution. Percentage inhibition was evaluated by comparing the test and blank solutions. The EC50 value (mg extract/mL) is the effective concentration at which hydroxyl radicals were scavenged by 50%. 28
Nitric oxide scavenging of freeze-dried extracts
Nitric oxide scavenging activity was measured spectrophotometrically. Sodium nitroprusside (5 mM) in phosphate-buffered saline, pH 7.4, was mixed with different concentrations of the extract prepared in ethanol and incubated at 25°C for 30 min. A control without the test compound, but with an equivalent amount of ethanol, was also used. After 30 min, 1.5 mL of the incubated solution was removed and diluted with 1.5 mL of Griess reagent (1% sulfanilamide, 2% phosphoric acid, and 0.1% N-1-naphthylethylenediamine dihydrochloride). Absorbance formed during diazotization of the nitrite with sulfanilamide and subsequent coupling with N-1-naphthylethylene diamine dihydrochloride was measured at 546 nm and the percentage scavenging activity was measured with reference to the standard (ascorbic acid). The EC50 value (mg extract/mL) is the effective concentration at which hydroxyl radicals were scavenged by 50%. 29
Statistical analysis
All parameters for antimicrobial and antioxidant activity were assessed in triplicate, and the results were expressed as mean±standard deviation values of three observations. The mean values and standard deviation were calculated with the Excel program from Microsoft Office, version 2010.
Results and Discussion
Effect of nitrogen source on mycelia growth rate
P. ostreatus PBS281009 and C. comatus M8102 mycelia growth in a liquid medium with different nitrogen sources is presented in Figure 1. After 7 days of culture, all nitrogen sources were found to be suitable for the growth of both mushroom mycelia in submerged culture. For P. ostreatus PBS281009, corn extract resulted in better growth than any other nitrogen source, followed by yeast extract and peptone. For C. comatus M8102, the best growth was achieved with the following nitrogen sources: peptone>corn extract>ammonium sulfate>yeast extract (P<.05). In comparison to organic nitrogen sources, inorganic nitrogen sources gave rise to relatively lower mycelial biomass, with an average difference of 30%±1.65%. This result reflects the effect of nitrogen on mycelial growth. 2,30,31 It has been suggested that certain essential amino acids for growth cannot be synthesized when in the presence of only inorganic nitrogen sources. The results obtained from other species, such as Leucopaxillus giganteus, Ganoderma lucidum, Cordyceps militaris, and P. citrinopileatus, show similar behavior. 2 The results are supported by data presented by Khandakar et al. 32 regarding the effects of peptone and yeast extract. It was reported that continued growth after this period causes a decrease in the quantity of obtained mycelium. The average decrease was 10%±1.43% to 20%±0.9% for both mushrooms. Aging of the hypha, visible under the microscope, was also noticeable. 33
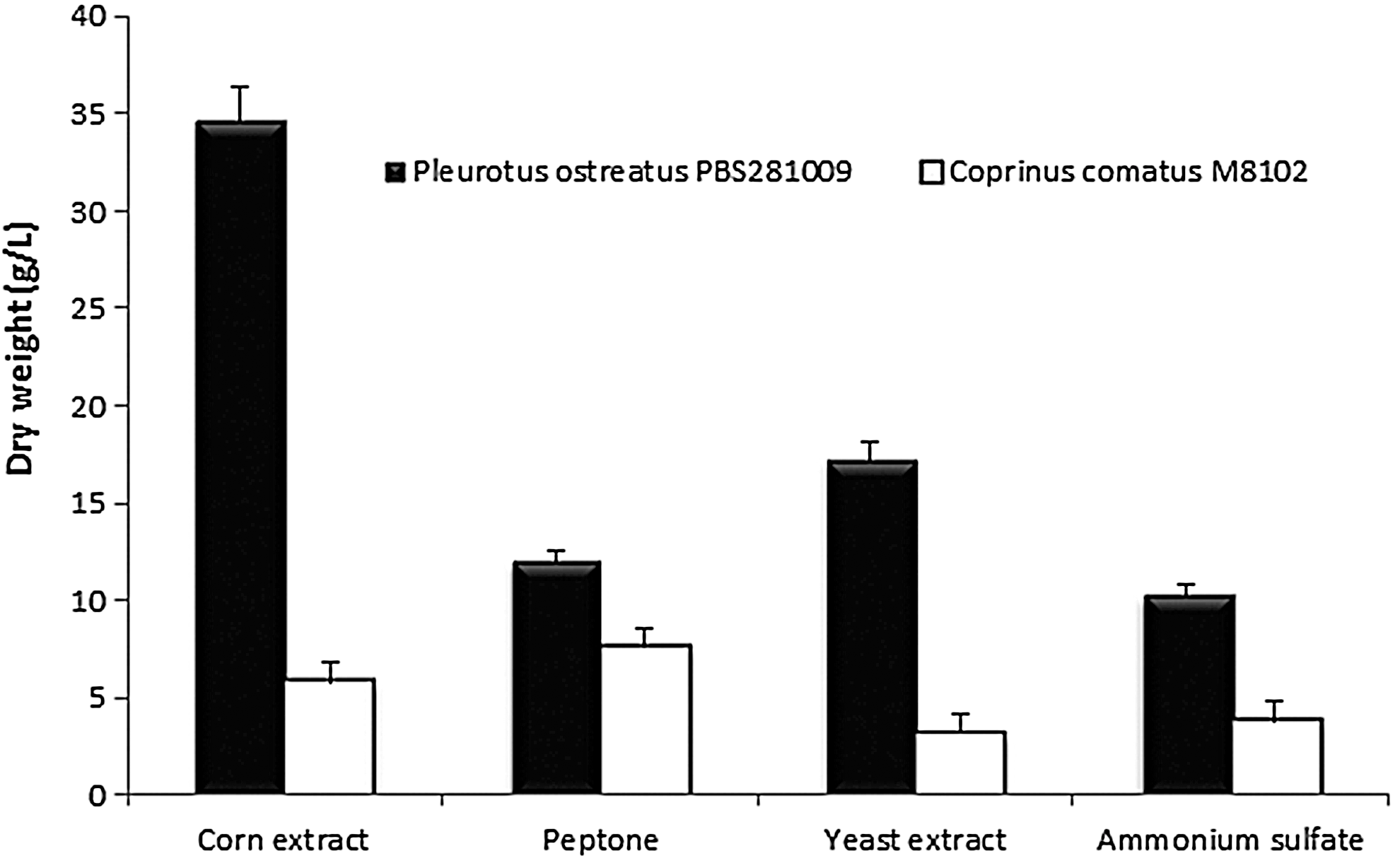
Mycelia growth of Pleurotus ostreatus PBS281009 and Coprinus comatus M8102 in the presence of different nitrogen sources. The results were significantly different P<.05.
Effect of nitrogen source on antimicrobial activity
The results of the antimicrobial activity of ethanolic freeze-dried extract of P. ostreatus PBS281009 and C. comatus M8102 are presented in Table 1. The mycelia extracts were screened for their antimicrobial properties against B. cereus, L. innocua, S. aureus, P. aeruginosa, E. coli, and two species of Candida. Results were recorded as the presence or absence of zones of inhibition. 34 The results revealed the antimicrobial potential of these extracts; in particular, the ethanol extracts of P. ostreatus PBS281009. Among the microbial species used as controls, Candida were the most sensitive to the two extracts, presenting MICs of 0.78±0.1 and 3.12±0.00 mg/mL for the mycelia obtained in the presence of organic nitrogen sources and inorganic nitrogen sources, respectively. Among the bacteria, Gram-positive specimens were found to be more sensitive to extracts, especially to that from P. ostreatus PBS281009. MIC of the extracts was found to be higher in Gram-negative bacteria, namely, P. aeruginosa (12.5±0.39 mg/mL) and E. coli (25±0.78 mg/mL), compared to Gram-positive bacteria (1.56±0.00 mg/mL). The extracts from C. comatus M8102 mycelium caused a significant increase in MIC for the 2 Candida species, up to 25±0.00 mg/mL. In this case, the maximum MIC values were recorded for the mycelia obtained in the presence of yeast extract and ammonium sulfate. For the other two organic sources of nitrogen, the MIC values varied between 3.12±0.00 and 12.5±0.36. These results are consistent with a series of previous studies performed with different edible mushrooms: Dictyophora indusiata, 35 Lactarius deliciosus, Sarcodon imbricatus, and Tricholoma portentosum. 2 The presented data show that the extracts of the two fungi species may be used for therapeutic purposes; these extracts are active at low concentrations against species of medical interest. 36
MIC, minimum inhibitory concentration.
Power reduction assay
Figures 2 and 3 show the power reduction of the two mushroom mycelium extracts. The yellow color of the reaction mixture changes to blue by varying degrees, depending on the reduction capacity of each extract. 37 The color intensity depends also on the species: the extracts from P. ostreatus PBS281009 mycelium caused a more intense blue color from the reaction mixtures than those from C. comatus M8102 mycelium. The reducing power of the mushroom ethanolic extracts increased with concentration. For the mycelium of P. ostreatus PBS281009, at 20 mg/mL, the reducing power values according to the source of nitrogen were: corn extract>peptone>yeast extract>ammonium sulfate. Also for C. comatus M8102, the greatest values were obtained in the presence of organic nitrogen sources compared to inorganic ones, in the following order: peptone>corn extract>yeast extract. Compared with BHA and TOC, the values are relatively equal, the differences being less than 10%±0.5%. The reducing power exceeds 1.5±0.07 at concentrations of 15 mg/mL for P. ostreatus PBS281009 and 0.5±0.02 for C. comatus M8102 per same amount of sample. The values correspond to those obtained in species, such as P. squarrosulus, 38 C. comatus, 4 and L. deliciosus. 39
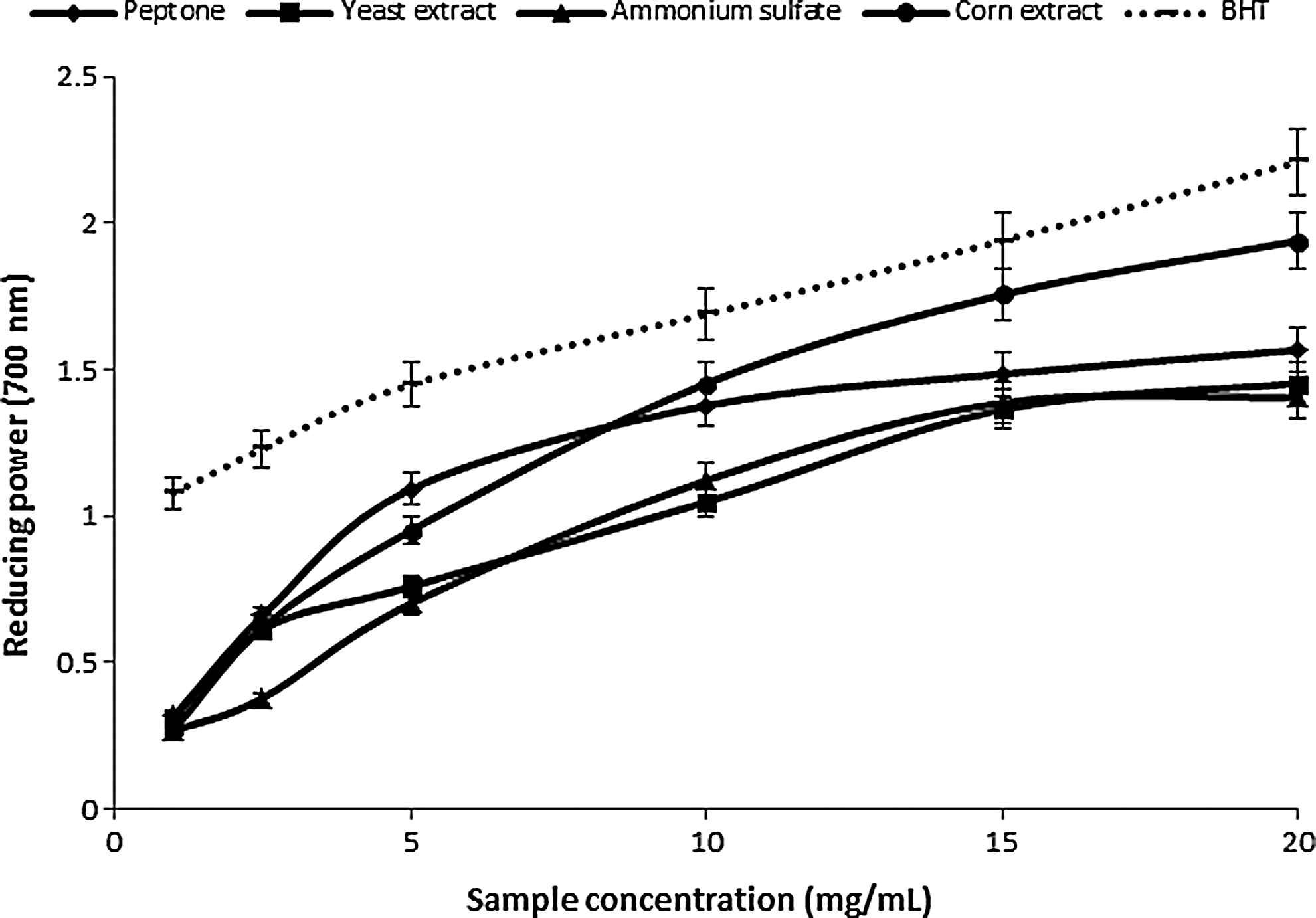
Reducing power of mushroom ethanolic extracts from P. ostreatus PBS281009.
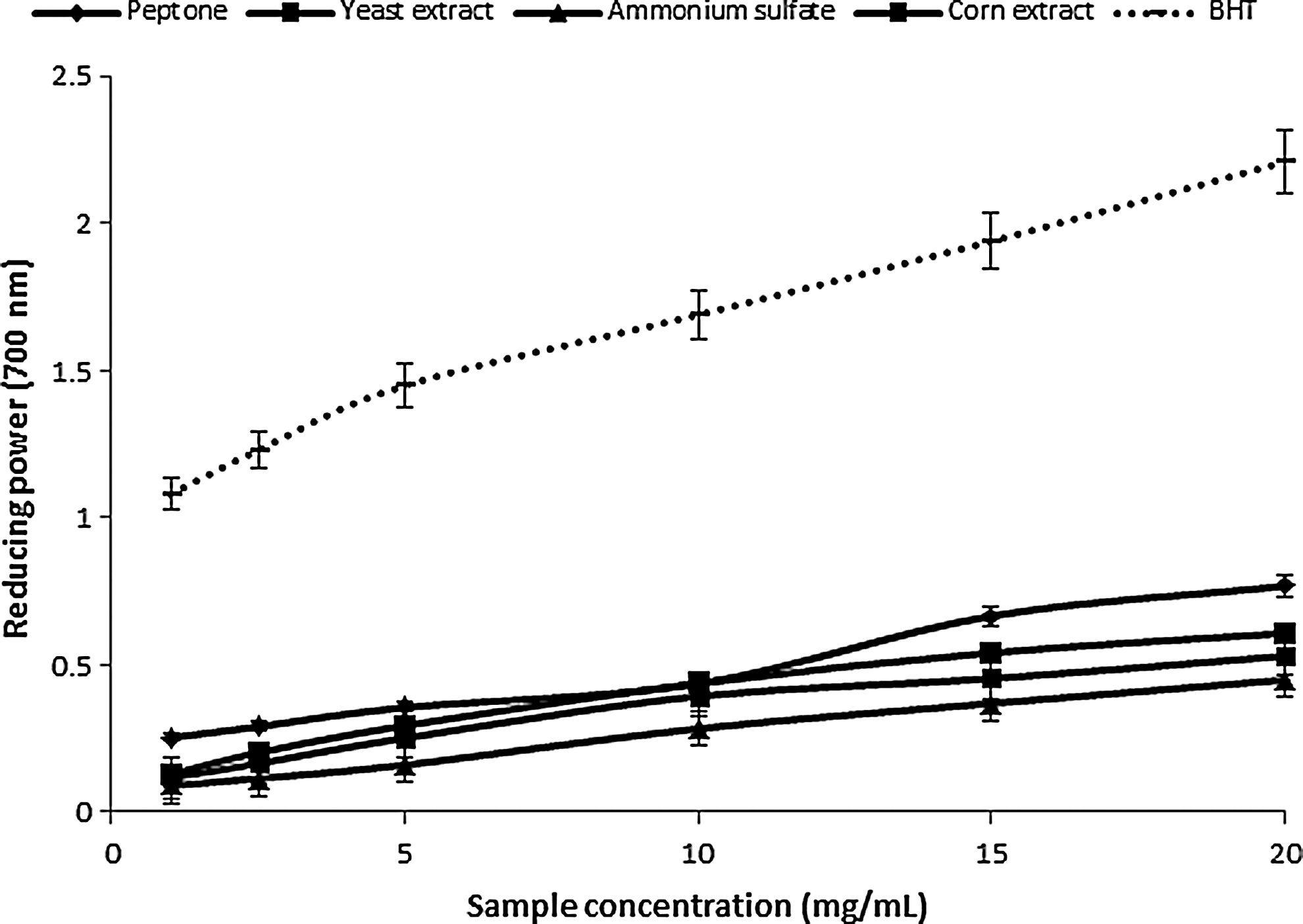
Reducing power of mushroom ethanolic extracts from C. comatus M8102.
Effect of nitrogen source on antioxidant compounds
Table 2 shows the phenol and flavonoid concentrations in the mycelium extracts obtained in the presence of 4 different nitrogen sources. For P. ostreatus PBS281009, the best nitrogen source was the corn extract (23.14±0.08 mg/g phenolic compounds and 0.32±0.17 mg/g flavonoids), while for C. comatus M8102, it was peptone, with 19.77±0.21 mg/g phenolic compounds and 0.33±0.03 mg/g flavonoids. Generally, the organic sources of nitrogen generated the synthesis of a quantity of compounds with higher antioxidant effects than the inorganic ones. This is also confirmed by the amount of lycopene and β-carotene, which conforms to the previously determined order of nitrogen source, although they are found in small amounts in mushroom extracts. It is of note that in the extracts of C. comatus M8102, mycelium lycopene or β-carotene were not determined. The quantities are confirmed by those reported for other, similar species in studies performed by Alam et al. 40 and Tsai et al. 4
The interest in extracts containing bioactive compounds has steadily increased in recent years due to the demand for products with a direct effect on human health. It is preferable to control the negative effects of free radicals, generated by the current pace, way of life, and diet standards, through the use of products containing bioactive compounds, such as phenols, flavonoids, ascorbic acid, and carotenoids. Thus, among the existing natural antioxidants in mushroom extracts, the phenolic compounds are the most important for the development of supplements or products with a preservative effect. 41 The phenolic compounds, including flavonoids, are known to exert a direct inhibitory effect on free radicals. 42 The effect is manifested through their chemical structure, in particular by the number of hydroxyl groups, which is directly proportional with antioxidant activity, and by their location in the molecule. These compounds reduce the risk of major chronic diseases and have antitumor effects. 43 In particular, the flavonoid mechanism of action includes antioxidant, anti-inflammatory, and antiproliferative effects, as well as detoxification of the body. For this class of biologically active substances, the primary role is cardiovascular protection, manifested through oxidation processes, reducing DNA degenerations, and the occurrence of tumors. 44,45
β-carotene and lycopene may play a major role in cancer and cardiovascular disease prevention and treatment; β-carotene acts as a prooxidant as its antioxidant action involves only small quantities of oxygen 46 and lycopene is an intermediary in carotenoid biosynthesis, including that of β-carotene. 47 Lycopene is considered an anticancer agent, particularly for the prevention of prostate cancer; it acts as an anticancer agent, reducing the incidence of tumors of the stomach and lungs, and also improves several cardiovascular problems and diabetes. 48
In addition to these compounds that have antioxidant effects, Pleurotus extracts also contain ascorbic acid and α-TOC. In addition to the inhibition of the O2 • and OH• radicals, ascorbic acid is involved, along with α-TOC, in the inhibition of lipid peroxidation. In addition, α-TOC is involved in DNA repair, thus inhibiting carcinogenesis. It also plays an important role in the inactivation of radicals, such as (ROO•), CCl3 •, OH•, and O2 •. Ascorbic acid is involved in various oxidative stress-related diseases, such as heart disease, stroke, and several neurodegenerative diseases. 49,50,51
In this study, the content of ascorbic acid in extracts from P. ostreatus ranged between 0.09±0.02 and 0.17±0.08 mg/g. The maximum amount was registered for the mycelium obtained in the presence of corn extract, while in the extracts of C. comatus mycelium, ascorbic acid was not identified. α-TOC was detected in all extracts, with a maximum of 9.2±0.23 mg/g extract for the C. comatus mycelium obtained in the presence of peptone. Yeast extract was again the less productive nitrogen sources from the point of view of antioxidant compounds.
Scavenging activity of DPPH radicals
The determination of DPPH radical binding offers a practical way to assess the antioxidant potential of the two extracts. DPPH radical is very stable in the presence of an antioxidant, obtaining an additional electron, and thus decreasing absorbance. 52 The degrees of discoloration resulting from the loss of the original purple color are indicative of the binding potential of free radicals. The DPPH scavenging activity reflected the concentration of the samples. Depending on the source of nitrogen for the extracts from P. ostreatus PBS281009 mycelium, the order of growth media according to the greatest scavenging activity was: corn extract>peptone>yeast extract (Fig. 4). For the same study of C. comatus M8102 mycelium, the following hierarchy resulted: peptone>corn extract>ammonium sulfate>yeast extract. Compared with ascorbic acid (45.97%±2.29% for 20 mg/mL), for the mycelium extract of P. ostreatus PBS281009, a value by 44.6%±2.2% higher was obtained at the same concentration and 25%±1.3% higher for the C. comatus M8102 mycelium extract (Fig. 5). Thus, the values of the binding capacity for the former mushroom were good, as it was close to those of known antioxidants. However, for the second species, the values obtained were moderate. Our findings indicate that the species used is very important, and our results for C. comatus M8102 were comparable to the results presented by Tsai et al. 4
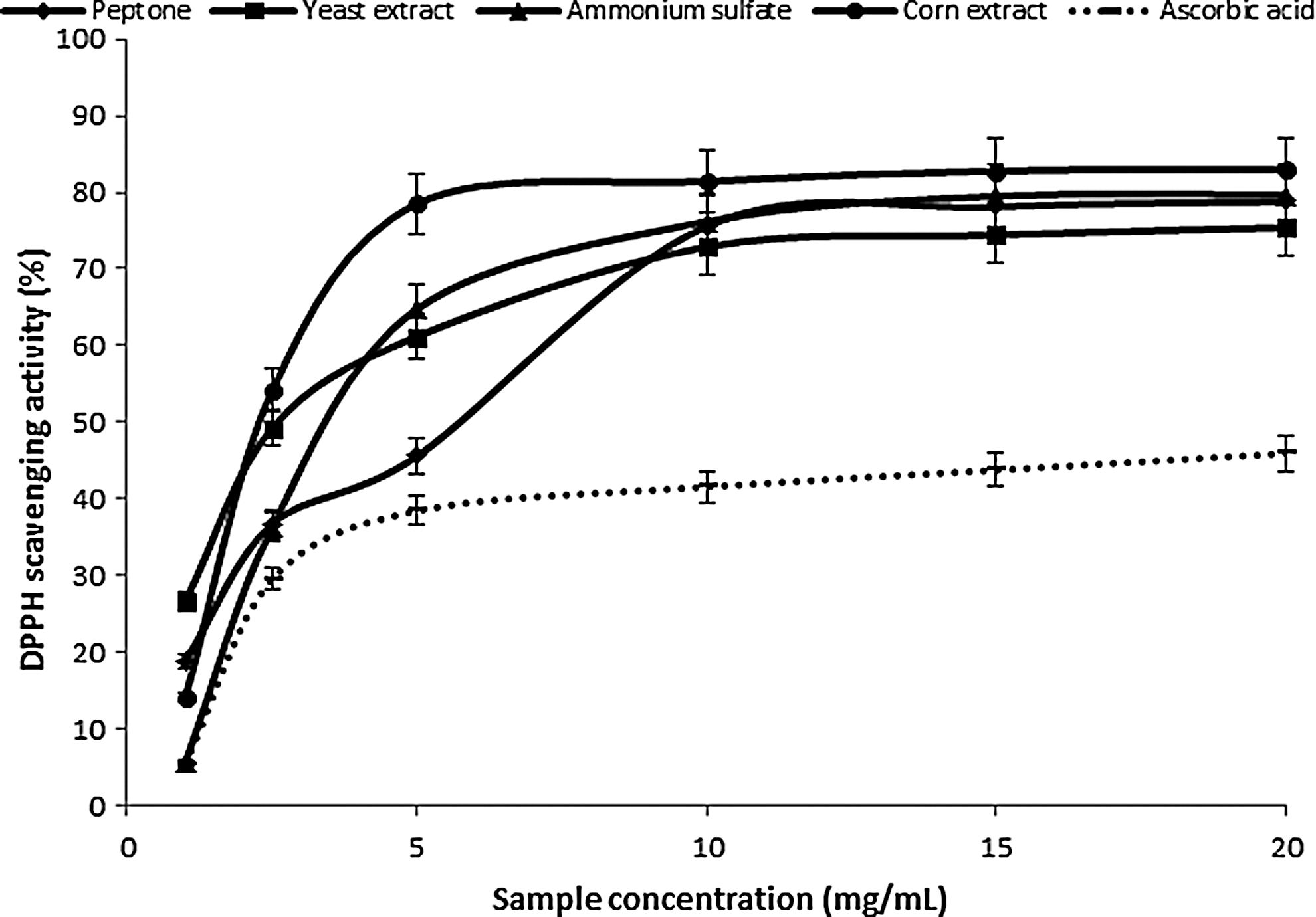
Scavenging ability of ethanolic extracts from Pleurotus ostreatus PBS281009 on 1,1-diphenyl-2-picrylhydrazyl (DPPH) radicals.
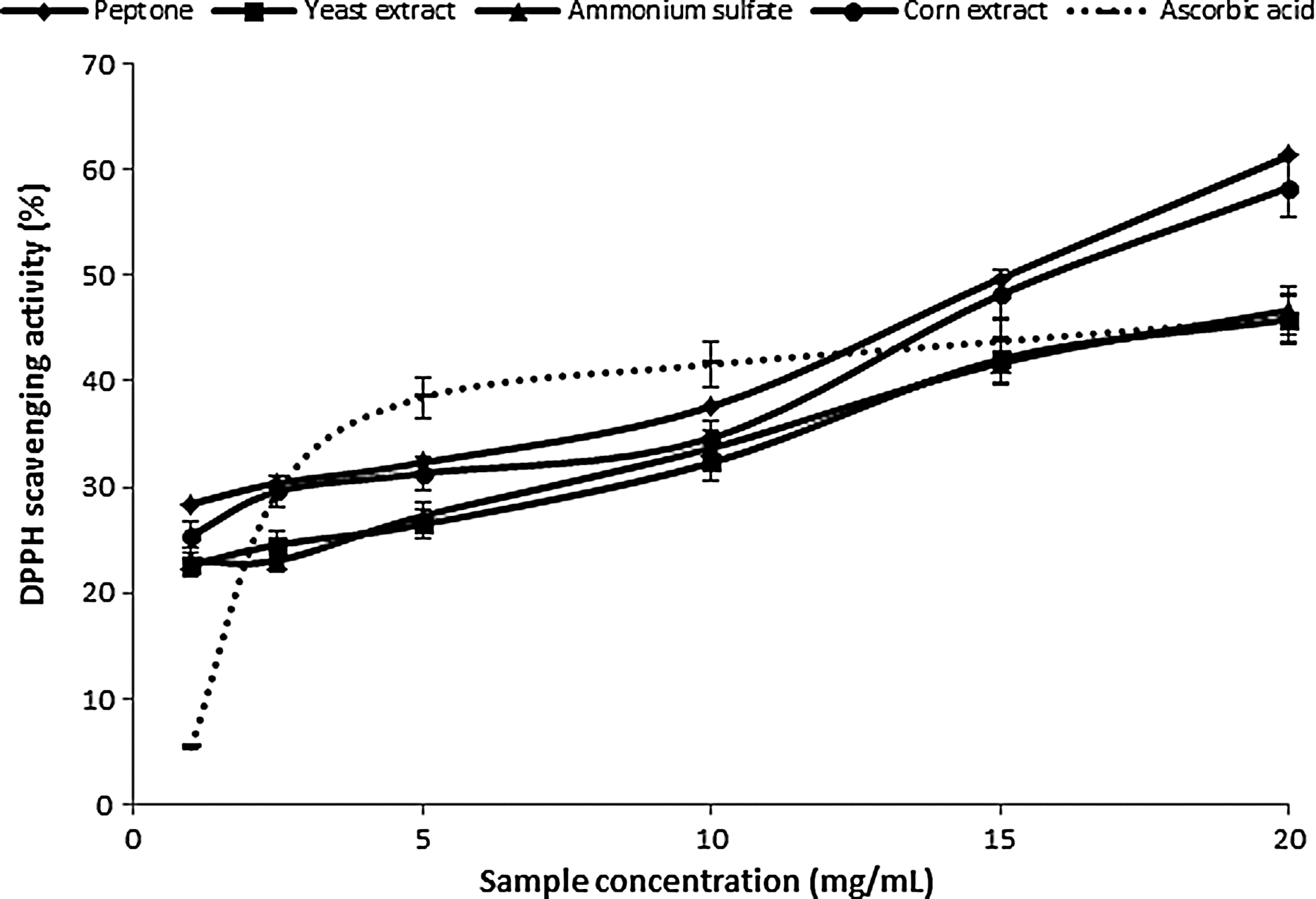
Scavenging ability of ethanolic extracts from C. comatus M8102 on DPPH radicals.
Compared to species of the genera Ganoderma, Boletus, and Agaricus, at concentrations similar to those from P. ostreatus PBS281009 mycelium extracts, the differences did not exceed those determined for ascorbic acid. 53
Antioxidant assay against β-carotene-linoleic acid
Depending on the nitrogen source, the extracts of the two species presented different patterns of antioxidant activity against β-carotene and linoleic acid. The antioxidant activity of carotenoids is based on the radical adducts of carotenoids with free radicals from linoleic acid. The linoleic acid free radical attacks the highly unsaturated β-carotene models. 2,54,55 For P. ostreatus PBS281009, the extract of the mycelium grown in the presence of corn extract presented the highest antioxidant activity, compared with the other three extracts (Fig. 6). Compared with TOC and BHT, this extract presented an activity of over 50%±3.0%, provided that its concentration was increased to 20 mg/mL. The mycelium of C. comatus M8102 showed a maximum activity when grown in the presence of peptones. In this case, the lowest values, 27.41%±1.4% for the 20 mg/mL sample concentration, were obtained from the extract of the mycelium grown in the presence of yeast extract (Fig. 7). For the tested extracts, the antioxidative components in the mushroom extracts reduced β-carotene destruction by neutralizing the linoleate free radical and other formed free radicals. 6,8 The inhibition capacity of linoleic acid oxidation of ethanolic extract was found to be consistent with the studies performed by Barros et al. 2 and Zengin et al. 18 They correspond to an inhibition of around 50%±2.8% from that of known antioxidants, such as BHT and TOC.
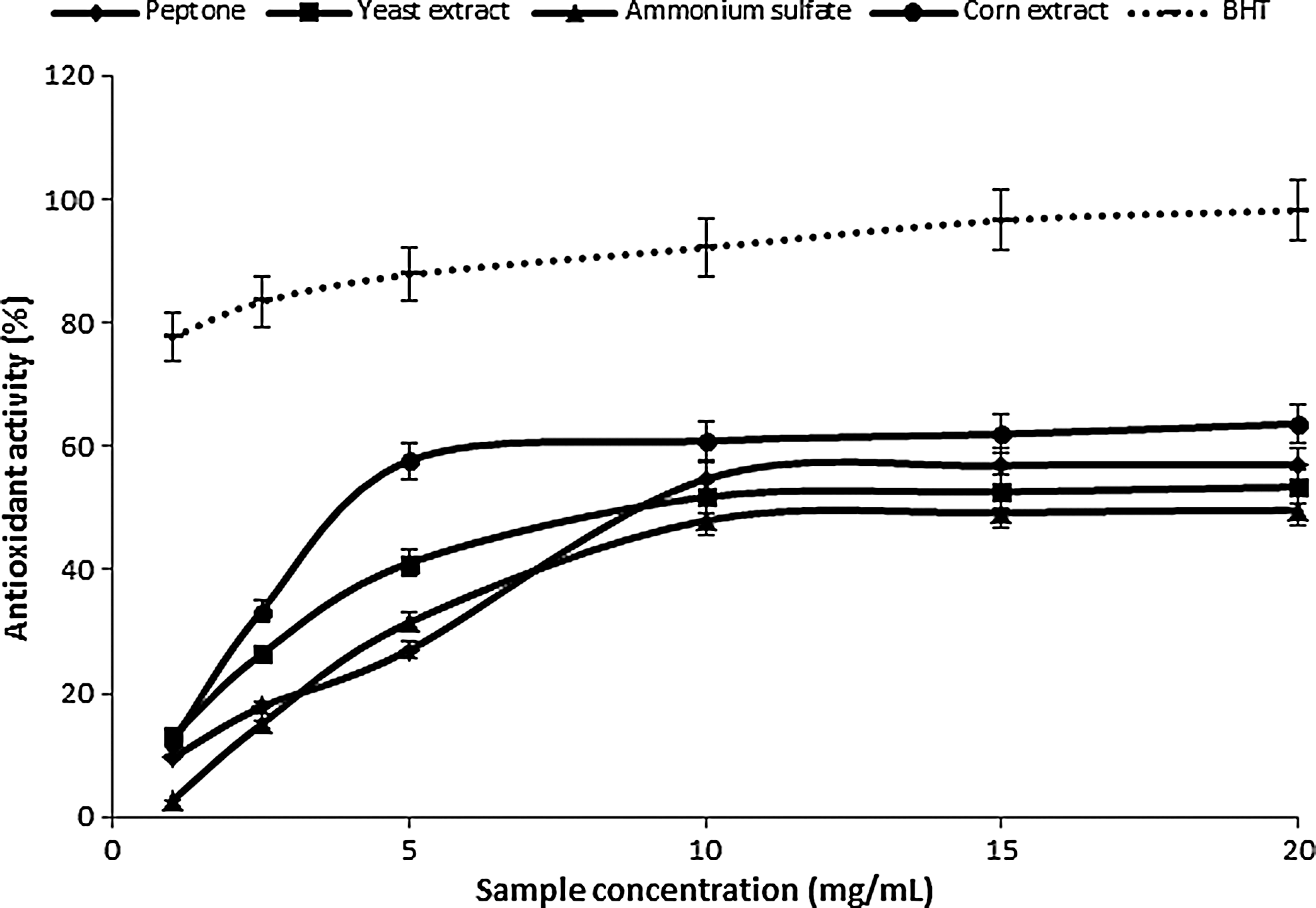
Antioxidant activity against β-carotene-linoleic acid of ethanolic extracts from P. ostreatus PBS281009.

Antioxidant activity against β-carotene-linoleic acid of ethanolic extracts from C. comatus M8102.
Scavenging effect on superoxide radicals
The superoxide radical scavenging activity of ethanolic extracts from lyophilized mycelium of P. ostreatus PBS281009 and C. comatus M8102 is important because it shows the ability to inhibit the precursor of several reactive oxygen species. 56 The extract of mycelium of P. ostreatus PBS281009, in the presence of corn extract, resulted in an EC50 value of 7.1±0.001 mg/mL (Table 3). The value was comparable with the value obtained with ascorbic acid (4±0.1 mg/mL). For C. comatus M8102, peptone generated the lowest EC50 value, but it was approximately 85% higher than the corresponding concentration of the previous species (47.03±0.02 mg/mL). The lowest values obtained were also generated in the presence of the inorganic nitrogen source. The values are comparable with results obtained for other species, including P. ostreatus, 57 Agaricus arvensis, Pleurotus salmoneostramineus, 40 and C. comatus, 9 and also with those obtained from plants. 18,58
EC50
Each value is expressed in mg/mL as mean±standard deviation (n=3).
Scavenging effect on hydroxyl radicals
The EC50 values in Table 3 demonstrate the potential of extracts from lyophilized mycelium of both fungi to scavenge hydroxyl radicals according to their concentration (1–20 mg/mL). The lyophilized mycelium of P. ostreatus PBS281009 had higher hydroxyl RSA compared with C. comatus M8102. This difference was manifested regardless of the nitrogen source used for submerged cultivation of the mycelia. Thus, for P. ostreatus PBS281009, the corn extract caused a minimum of 3.76±0.05 mg/mL, which is a recognized value for alcoholic extracts from this species. 54 The value is also supported by other measurements, performed in algae-like Ulva fasciata 59 or in medicinal plants, such as Rhododendron arboretum. 60 For C. comatus M8102, the values were significantly higher and this was previously reported in a series of previous studies. 9 As presented by Tsai et al. 4 in connection with the species C. comatus GK-131, this feature is specific only to the aqueous extracts and their EC50 values ranged from 53.98±0.96 to 94.00±0.47 mg/mL (Table 3). The fact that P. ostreatus species are regarded as good scavengers of hydroxyl radicals is also supported by an EC50 value of approximately 8±0.5 mg/mL for these fungi, which is comparable to the EC50 of ascorbic acid, which is 6±0.2 mg/mL.
Scavenging activity of nitric oxide
Determination of nitric oxide scavenging activity is important because of its involvement in a number of inflammatory diseases. It was noted that its inhibitors significantly reduce inflammations at the tissue level. 28 The effect is dependent on the sample concentration. In this case, the minimum EC50 value (14.76±0.002 mg/mL) in P. ostreatus PBS281009 was obtained in the presence of corn extract and the value obtained for ascorbic acid was ∼0.1±0.01 mg/mL. For C. comatus M8102, the EC50 minimum value was 41.23±0.01 mg/mL, obtained in the presence of peptone, as in previous cases (Table 3). The results indicate that the ethanolic extract of the lyophilized mycelium from both fungi contained compounds capable of inhibiting nitric oxide and corresponds with previous data from both medicinal plants and for Cyperus rotundus rhizomes. 61 The values obtained for the mycelium of both fungi are adequate and comparable to the data from aqueous extracts of P. rimosus. The results are important because the aqueous or alcoholic extracts of known medicinal mushrooms, such as G. lucidum, P. pulmonarius, and P. florida do not have nitric oxide scavenging activity. 18
A large number of edible mushrooms are also used for their antioxidant properties. Additional beneficial effects include dietary fiber, minerals, and vitamins. The antioxidant activity, determined by various methods, demonstrates that the mycelium obtained by submerged cultivation and freeze-dried are adequate for the preparation of extracts with high biological activity. In addition, it was observed that the source of organic nitrogen determines the development of mycelium-rich substances with antioxidant properties. 62 The current studies have shown that, for the two used species used in these experiments, ammonium sulfate does not generate a mycelium that contains large amounts of antioxidant substances. This was demonstrated in both species, irrespective of the growth medium used, with regard to the type of radicals inhibited and the reduction power.
Comparing the antioxidant activity of extracts from the mycelium of both species has revealed that corn extract is the best source of nitrogen for the PBS281009 species and peptone is the best source for M8102 and the yeast extract generated the lowest EC50 values. Compared to other studies conducted exclusively in the presence of inorganic nitrogen sources, 2 the presented data show that the species has a major role in determining the antioxidant activity of mycelium extracts. Also, lyophilizing the mycelium represents a relatively simple way of preserving higher quantities of biologically active substances in the mycelium and in the extract. The obtained data demonstrate a direct link between the amount of phenols, flavonoids, and carotenoid compounds in the presence of the corn extract and, respectively, of peptone. The increased antioxidant capacity of PBS281009 is also confirmed by the existence in the extracts of the two carotenoid compounds, since the differences between the phenolic compounds and the flavonoids are low in both species.
For phenols, the difference is, on average, 11.5%±0.1% in favor of PBS281009 for organic nitrogen sources and, in M8102, approximately 3%±0.15% for ammonium sulfate. For flavonoids, the difference was 4%±0.5% for ammonium sulfate and for organic nitrogen sources the amount was higher, on average by 7.9%±0.21% for M8102, except for corn extract where the determined values were similar.
The absence of carotenoids in the mycelium demonstrates that the EC50 values may have differences of even more than 90%±0.1% between the extracts from the two mycelium species. For PBS281009, the corn extract resulted in a minimum EC50 value compared to other sources of nitrogen. For M8102, the EC50 differences for peptone and corn extract are generally low, about 6.5%±0.06%, with the exception of hydroxyl RSA. In the case of yeast extract, differences exceed 20%±0.1% in favor of peptone. When using ammonium sulfate, the differences reach up to 50%±0.02% with respect to superoxide RSA.
Free radicals interfere with human organisms at the cellular and molecular level, causing disruptions of normal functions and disease. Reactive oxygen species (ROS), such as super oxide, hydroxyl radicals, and reactive nitrogen species, such as nitric oxide, are molecules with higher reactivity. In the human body, ROS play an important role in cell death. 63 –65 Free radicals contain a number of electrons that make them unstable and cause damage at the molecular level. Through such mechanisms, they are the cause of many diseases, resulting in the need to find reliable, natural sources capable of inhibiting these chemical species. 66 Natural extracts, particularly those from mushrooms, represent a modern way to prevent or alleviate the effects of these molecules. The content of phenolic substances, carotenoids, and other molecules with antioxidant effects represents a method of quantifying the mitigating effectiveness of an extract. In the case of the two types of mycelium tested in this article, the EC50 values were dependent on the content and on the nitrogen source from which it was obtained.
The obtained data show that mycelium with high biological activity can be relatively easily obtained by submerged cultivation in the presence of corn extract or peptone, as sources of nitrogen. The capacity of the mycelium to inhibit the effects of hydroxyl radicals, superoxide anions, and nitric oxide, which all cause damage to biological systems and significant inflammatory effects, depends on a suitable source of nitrogen in the growth media. These observations are also supported by their reduction capacity, which reveals the antioxidant level of an extract.
There is little data on compounds that determine the antimicrobial effect of the extracts from the Pleurotus species; the antimicrobial effect is a response by the fungus to the environment. In the case of liquid medium cultivation, P. ostreatus ceases the production and secretion of a number of substances with antimicrobial effects into the culture medium. 67 The effectiveness of said inhibition depends on the composition of the culture medium and on the species used. One of these substances is pleuromutilin, which inhibits Gram-positive bacteria. 68 Following extraction with hexane and dichloromethane, p-anisaldehyde was isolated, and it reportedly inhibits the growth of B. subtilis, P. aeruginosa, and Aspergillus niger. 69 There are some studies that show that even the polysaccharides synthesized by P. ostreatus species exhibit an antibacterial effect. 63 These studies were conducted in submerged cultivation of the mycelium and showed that the spectrum of inhibition depends on the carbon and nitrogen sources of the culture medium. 2,70 Also, submerged cultivation involves the synthesis of oxalic acid, which is responsible for a significant portion of the antimicrobial effect of biologically active products from these fungi. 71
From the results, obtained by statistical analysis, has emerged the importance of bioactive compounds both at the antioxidant level and concerning the antimicrobial properties of each mycelium (lyophilized extract). These observations do not exclude the differences between the two species as a biological value. Following analysis, it was found that the biological value is influenced by the nitrogen source used for obtaining the mycelium, and also by the method of processing the extract. 72 The quantitative differences among the components with a biological effect were revealed by the lack of β-carotene, lycopene, and ascorbic acid in the mycelium extract from the species C. comatus M8102. The content of phenolics determined in extracts from P. ostreatus PBS281009 has a good correlation with the free radicals scavenging activity (R 2 =0.724±0.096 to 0.918±0.025), with reducing power (R 2 =0.786±0.043 to 0.889±0.033), and with antioxidant assay against β-carotene–linoleic acid (R 2 =0.768±0.079 to 0.913±0.007). Also, a correlation was found between total phenols and flavonoids content and antimicrobial activity, depending on the species. 73 Coefficient R 2 had good values (0.871±0.12 to 0.974±0.01) for inhibition of two strains of Candida, with maximum specificity for corn extract and peptones as nitrogen sources. In the case of bacteria, a moderate correlation resulted for the three strains of Gram-positive, excluding yeast extract and ammonium sulfate. By analyzing all these data, results showed that the phenolic content represented the main component which expressed not only antioxidant, but also antimicrobial activity. The mode of action upon each group of microorganisms depends also on the chemical structure of these compounds. 74
The ethanolic extracts of P. ostreatus PBS281009 and C. comatus M8102 mycelia have significant antioxidant and antimicrobial activity in the presence of organic nitrogen sources at the expense of the inorganic source, the ammonium sulfate. These properties achieve a maximum value in the presence of corn extract for P. ostreatus PBS281009 and peptone for C. comatus M8102. Lyophilized mycelium extracts possess noticeable antimicrobial activity against Gram-positive, Gram-negative bacteria, and two species of the genus Candida. The mycelium produced by submerged cultivation can exert an effective antimicrobial effect against microorganisms that can cause disturbances in the human gastrointestinal tract. The ethanolic extracts from the oyster mushroom P. ostreatus and C. comatus showed concentration-dependent antioxidant activity. In addition, these extracts, mainly those from the mycelium of P. ostreatus PBS281009, contain significant amounts of phenolic compounds, β-carotene, and lycopene, which contribute to their antioxidant activity, but is dependent on the nitrogen source. It follows that the mycelium obtained, in particular, the ethanol extract of its lyophilizate, constitute a significant source of compounds that can act as a barrier against free radicals. Therefore, the ethanolic extracts can be used as food supplements of a nutraceutical nature, with demonstrated antioxidant and antimicrobial effects. Future studies will consider the direct effect of lyophilized mycelium and its extracts on human intestinal microflora, as well as determining their effect on the immune system.
Footnotes
Acknowledgment
This research was financed through the project PNCDI II CNCSIS–Human Resources, Theme 9/2010.
Author Disclosure Statement
No competing financial interests exist.