
Abstract
To understand host gastrointestinal response after exposure to probiotic Lactobacillus acidophilus L-92, microarray analysis of cultured epithelial Caco-2 cells was performed. Of the 187 genes down-regulated after 4 h treatment with L-92, 25 were involved in RNA splicing; 12, in cell cycle; 8 were transcriptional regulators; 2 were involved in ubiquitin proteolysis; 2, in adhesion; 2, in meiosis; 2, in splicing; and 2 encoding cytokines. In the RNA splicing group, genes encoding small nuclear RNAs, nuclear pore complex interacting proteins, RNA binding motif proteins, and SMG1 homologs (phosphatidylinositol 3-kinase-related kinase) were identified. Among the only 13 genes up-regulated by the treatment, 5 were involved in histone structure, and 2 were involved in metabolism. Genes belonging to cell adhesion, transmembrane proteins, mitogen-activated protein kinase, immune response, DNA binding, inflammation, and protein synthesis groups were mainly up-regulated after 20 h of treatment, whereas no significantly down-regulated genes were observed. In the present transcriptome analysis, during the early stage of treatment (four hours of treatment) with L-92, genes involved in cell growth and cell meiosis were mainly repressed. During the late phase of treatment (20 h of treatment), the expression of the genes linked to cell adhesion activity and metabolism for cell growth was enhanced. From the present transcriptome analysis, we suggest that Caco-2 cells slow down cell death and turnover of RNA synthesis as an early response to L-92 treatment; at the late stage of treatment, the genes involved in cell proliferation, transcriptional activity, and apoptosis are activated.
Introduction
M
Recently, transcriptome analyses of human intestinal samples, collected from biopsies during clinical trials, were conducted to study host GI events. 9 –12 Studies have sought to understand the host response after the intake of probiotic cells. However, the collection of human samples during clinical trials is not an easy task; GI cells are strongly affected by diet, but it is difficult to control the subjects' diet.
Caco-2 cells are a well-established human epithelial cell line and have been used in many studies pertaining to transepithelial transport, metabolism, and disease. 13 –15 A recent study analyzed changes in the host response through the analysis of Caco-2 cells using microarray analysis. 16 –18 Caco-2 cells could be used to compare differences in probiotic characteristics of different bacterial strains. Of particular interest, Caco-2 cells overlayed on macrophage-like RAJI cells showed cross-communication in the transwell system. 19 –22 A transwell system using co-cultivating Caco-2 cells with macrophage-like cells, such as THP-1 cells and RAJI cells, serves as a useful tool for studying cross-talk between epithelial cells and immune cells. 23,24 In the use of the transwell system, inflammatory damage caused by pathogenic bacterial treatment and its rescue by some probiotics has been reported. 24
Based on a comparative proteome analysis using L. acidophilus L-92 and the less adhesive CP23 strain, 25 it was suggested that surface layer protein A (SlpA) of L. acidophilus L-92 could serve as an adhesion molecule with Caco-2 cells to aid the communication between L-92 cells and host cells. Results show L-92 to be more effective in releasing an inflammatory cytokine, IL-12, from splenocytes compared with CP23, which has less SlpA. Changes in some cytokines and chemokines after uptake of L-92 were reported in animal studies. 6,26 However, information processes, in addition to host metabolism, is required.
In the present study, using microarray analysis, we investigated changes in gene expressions link to metabolism occurring in GI epithelial cells after exposure to L-92 cells.
Materials And Methods
Bacterial strain
L. acidophilus L-92 strain was used from our stock culture collection and was maintained anaerobically at 37°C for 20 h. in Man Rogosa Sharpe (MRS) broth (Difco Laboratories, Detroit, MI).
Caco-2 cell culture
The human colonic cell line Caco-2 was obtained from the RIKEN Cell Bank (Tsukuba, Japan) and maintained in Dulbecco's modified Eagle's medium (Sigma, St. Louis, MO) containing 10% (V/V) of heat-inactivated fetal bovine serum (Hana-Nesco Bio, Tokyo, Japan), 1% (W/V) amino acids (Sigma), streptomycin (100 μg/mL), and penicillin (100 U/mL) (GIBCO, Grand Island, NY) in a 100 cm2 dish (Corning Glass Works, Corning, NY) at 37°C in 5% CO2/95% air. The culture medium was replaced every other day to maintain the cells. Monolayered cells were collected by adding 0.25% trypsin (Sigma) for 5 min, and were seeded at 4.5×105 cells/well in 3.0 mL of the cell suspension.
L-92 treatment of Caco-2 cells
For the transcriptome analysis assay, Caco-2 cells were grown in six-well tissue plates (Asahi Glass Co., Ltd., Tokyo, Japan) at 37°C. The medium was changed every other day. The assay was performed on Caco-2 cells on day 21. On day 21, the L-92 cells cultured in MRS medium were collected by centrifugation at 13,000 g for 10 min, and resuspended in phosphate-buffered saline (PBS) (8.15 mM Na2HPO412H2O, 1.47 mM KH2PO4 2.68 mM KCl, and 0.137 M NaCl, pH 7.4) containing 0.137 M NaCl. The L-92 cells were collected again by centrifugation at 13,000 g for 10 min. The washed cells were resuspended in PBS and heat treated at 100°C for 10 min. Then, 1×1010 cells of heat-killed L-92, suspended in 3 mL of culture buffer were added to each well of the culture plate. After centrifugation of the plate at 3000 g for 1 min to accelerate the association of L-92 and Caco-2 cells, the mixed cells were incubated at 37°C in 5% CO2/95% air. After 4 and 20 h of incubation, the supernatant from the mixed cultured cells was discarded, and the Caco-2 cells were washed twice with PBS to remove nonbound cells. Finally, the Caco-2 cells were harvested using a cell scraper after the addition of cold PBS. After centrifugation, 1.5 mL of RNAlater solution (QIAGEN K. K., Tokyo, Japan) was quickly added to the collected cell pellet to extract RNA. The suspension was maintained at 4°C for 12 h, and then stored at −30°C. Total RNA was purified and used for cDNA synthesis after analysis of its quality. Untreated cells were used as controls. Three replicate samples were prepared for each group.
Microarray analysis
Total RNA was extracted using an RNeasy Mini Kit (QIAGEN, Valencia, CA). Double-stranded cDNA was synthesized from 5 μg of total RNA, and the cDNA was subjected to in vitro transcription in the presence of biotinylated nucleotide triphosphates. Human genome-wide gene expression was examined using the GeneChip® Human Gene 1.0 ST Array (Affymetrix, Santa Clara, CA) containing oligonucoleotide probe sets for approximately 28,869 full-length genes and expressed sequence tags. The biotinylated cRNA was hybridized with the probe array for 16 h at 45°C. The hybridized products were stained with streptavidin-phycoerythrin, and then scanned with a Hewlett–Packard Gene Array Scanner (Palo Alto, CA). The fluorescence intensity of each probe was quantified using GeneChip Analysis Suite 5.0 software (Affymetrix). The level of gene expression was determined as the average difference using the GeneChip software.
Statistical and functional analysis of microarray data
Data analysis was performed using Genespring GX software, version 11.5.1 (Silicon Genetics, San Carlos, CA). Expression data were considered significant when they differed by at least 1.25 fold between the L-92 treatment and the PBS treatment, as P values of <.05 were observed when statistically analyzed by T test unpaired with Benjamini–Hochberg as post test.
Results
Changed gene expressions after four hours of treatment of L-92 cells
Most of the changes in gene expression in Caco-2 cells were relatively mild after treatment with L. acidophilus L-92. However, significant changes were observed. After the 4 h of treatment with L. acidophilus L-92, 187 genes were significantly down-regulated by <0.77-fold, and 12 genes were up-regulated by >1.25-fold in Caco-2 cells compared with nontreated cells, as illustrated in Figure 1. After 20 h of treatment, 45 genes were significantly up-regulated in Caco-2 cells by >1.25-fold (Fig. 1). For a detailed understanding of the response of the cultured Caco-2 cells, up-regulated and down-regulated genes were categorized based on the common pathways in KEGG analysis (
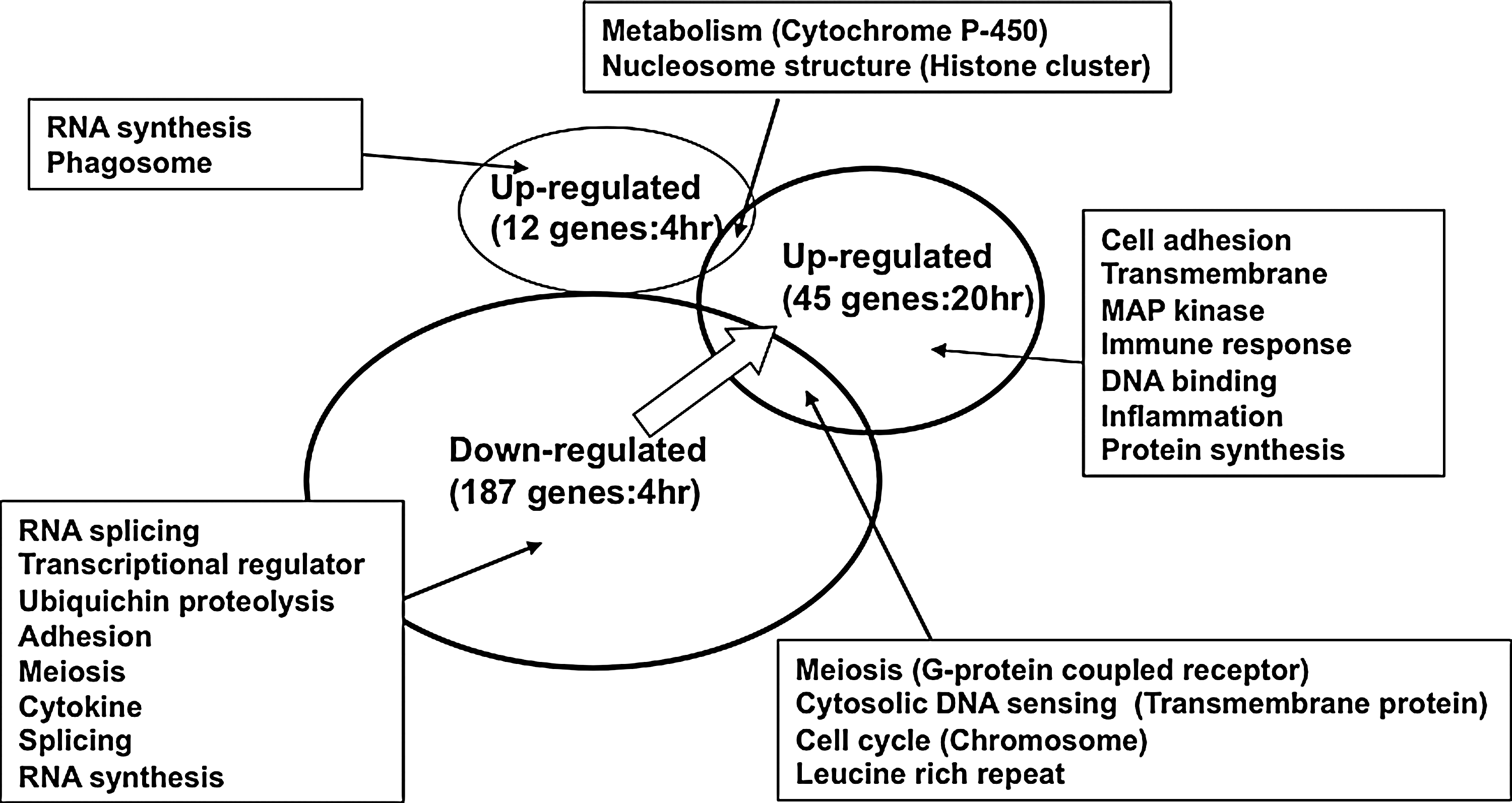
Profile of the changed genes in the transcriptome analysis of Caco-2 cells after 4- and 20-h treatment with Lactobacillus acidophilus L-92. Genes categorized by KEGG analysis are shown in the square.
Changed gene expressions after 20 h of treatment of L-92 cells
After the 20 h of treatment, genes belonging to the categories of cell adhesion, mitogen-activated protein (MAP) kinase, transmembrane function, immune response, metabolism, and others were mainly up-regulated (Fig. 1). Among the up-regulated genes, two dual-specificity phosphatase genes and two small Cajal body-specific RNA genes in the MAP kinase pathway, four carcinoembryonic antigen-related cell adhesion molecule genes, two amphiregulin genes, and two integrin genes for cell adhesion were up-regulated (Table 2). No significantly down-regulated genes were observed in the 20 h treatment group.
Changed gene expressions after 4 and 20 h of treatment of L-92 cells
Only two genes in the metabolism and nucleosome structure group were significantly up-regulated after both the 4 and 20 h of treatments. Genes for meiosis, cytosolic DNA sensing, cell cycle, and leucine rich repeat were down-regulated after 4 h but up-regulated after the 20 h of treatment. Throughout the present transcriptome analysis, during the early stage of treatment with L-92 (four hours of treatment), many genes linked to cell growth and cell meiosis were mainly repressed, as shown in Figure 1. During the late phase of the treatment (20 h), many genes linked to cell adhesive activity and metabolism of cell growth, such as DNA synthesis, protein synthesis, and kinase activity, were increased (Fig. 1).
Summary of the changed gene expressions in L-92 cells
To help understand the events occurring in the Caco-2 cells after L-92 treatment, many of the genes listed in Tables 1 and 2 are illustrated in Figures 2 and 3. During the early stage of the treatment (4 h), genes such as FBX, RNF, ZNF, and ATF (Table 1) were down-regulated, and two metabolism-related genes, both involved in apoptosis, were up-regulated (Fig. 2). Moreover, genes involved in RNA splicing and RNA degradation were down-regulated, and genes encoding RNA synthesis were up-regulated (Fig. 2). Genes encoding the adhesive molecule mucin and some genes linked to transcriptional regulation and MAP kinase were down-regulated (Fig. 2). In addition, genes linked to phagocytosis and the G-protein-coupled receptor were also down-regulated. These results suggest that Caco-2 cells might respond to L-92 treatment by slowing down cell death and RNA degradation, and accelerating RNA synthesis as an early response. After the 20 h of treatment, most of the genes linked to cell proliferation, gene regulation, MAP kinase, apoptosis, and some adhesive molecules were up-regulated (Fig. 3). These results suggest that long-term treatment of L-92 (20 h) might activate cell proliferation, transcriptional activity, and apoptosis in Caco-2 cells.
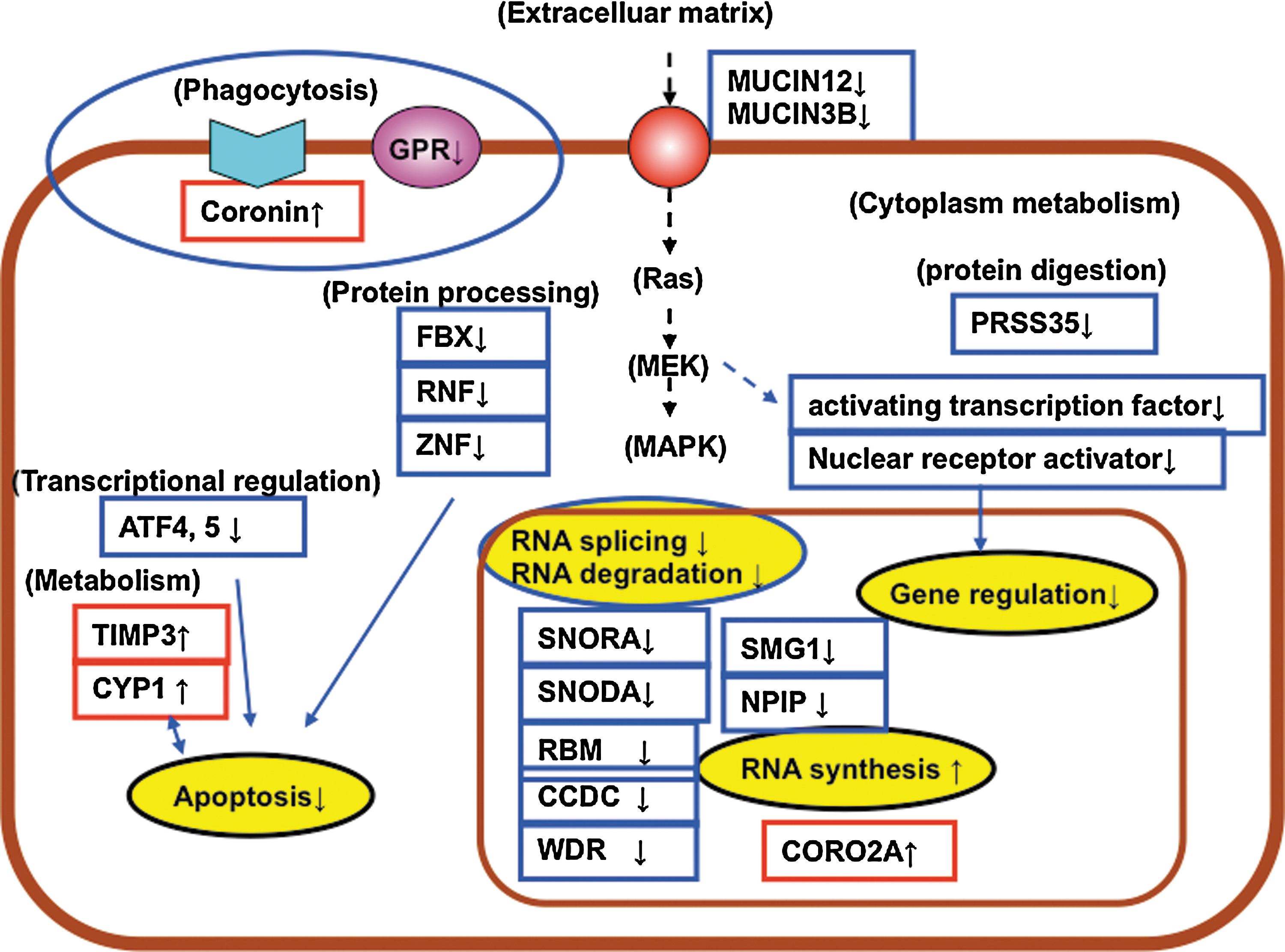
Genes mainly changed in Caco-2 after 4-h exposure to L. acidophilus L-92. Down-regulated genes are indicated with blue, while up-regulated genes are red. Categorized networks are circled.
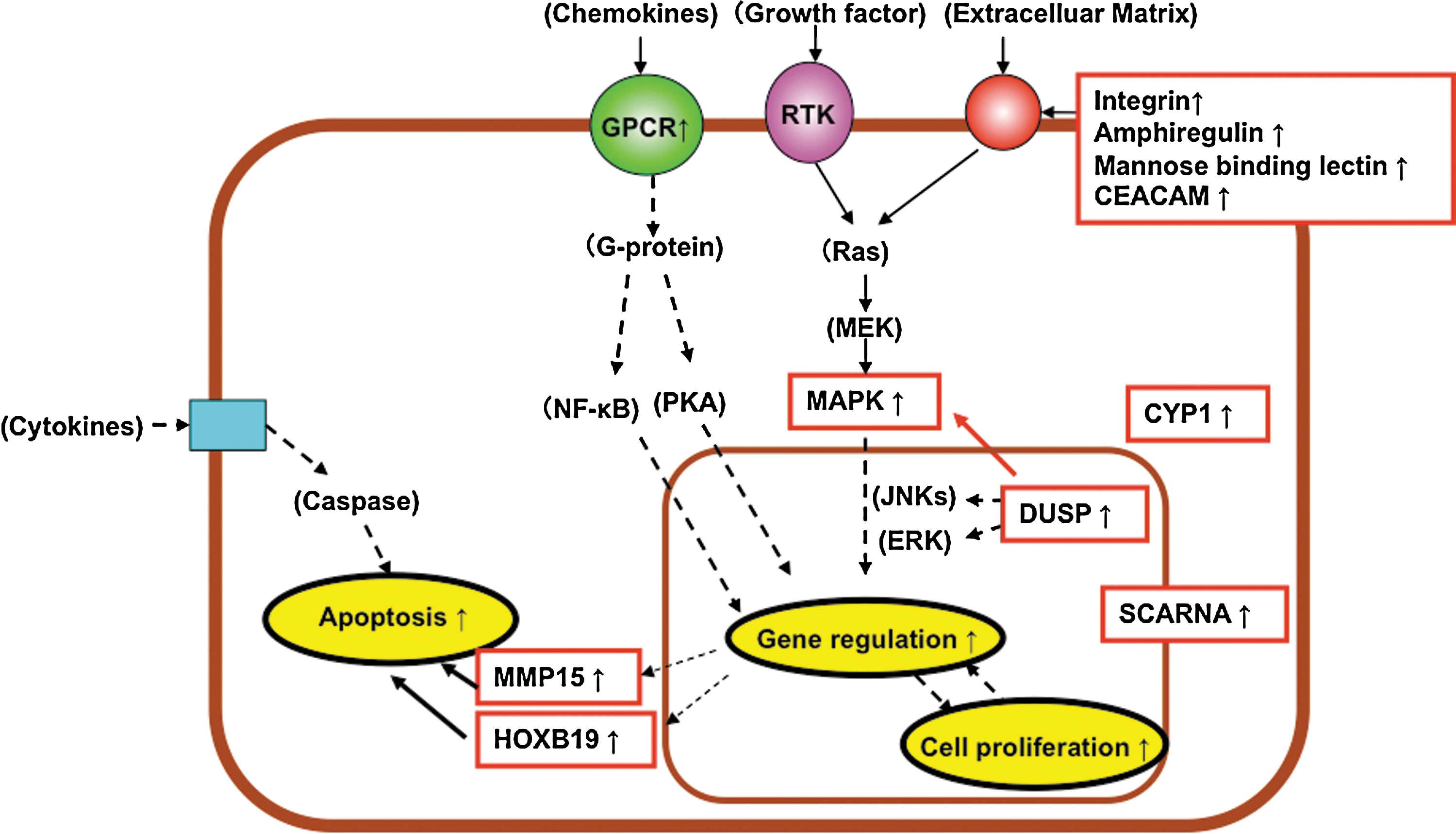
Genes mainly changed in Caco-2 cells after 20-h exposure to L. acidophilus L-92. Up-regulated genes are indicated with red. Categorized networks are circled.
Discussion
Host-bacterial cross-talk between probiotic bacteria and GI epithelial cells is thought to be an important process in eliciting the host response, including defensive responses against pathogenic bacteria and immune responses. To understand the probiotic roles of the host GI cells, transcriptome analyses for human GI cell samples collected by biopsy are indicated. However, this is not an easy task. Separations of the epithelial cells and lamina propria immune cells, which may play different roles in host defense and immunomodulatory effects, are also difficult to achieve. To mimic the in vivo gut environment, human intestinal epithelial Caco-2 cells have been utilized to study GI events, epithelial disease status, transport of nutrients, and bioactive components. 27 –29 In the present study, we performed a transcriptome analysis of epithelial Caco-2 cells to analyze the host GI response after the uptake of probiotic L-92 cells. Specific binding of the L-92 cells to Caco-2 cells was expected via a cell-wall-associated adhesive molecule, SlpA, present in L-92 cells. 25 Changes in gene expression profile in Caco-2 cells were relatively mild, but many significant changes were observed by microarray analysis after 4 and 20 h of treatment, as summarized in Figures 2 and 3. The results suggest that Caco-2 cells are prevented from cell death, categorized as apoptosis, and cell proliferation (growth) during the early phase of the treatment (Fig. 2). Based on these results, Caco-2 cells seem to respond first to the stress from L-92 binding, then adjust their activities to cell proliferation during the late phase of the treatment. By repeated oral uptakes of probiotic L-92 cells, responses observed in Caco-2 cells at 20 h would be most likely expected in the epithelial GI tract in humans.
Probiotic strains are thought to communicate with intestinal epithelial cells in a strain-specific manner. Some of the responses observed in this study were previously reported using different lactic acid bacterial strains in animal, human, and cultured cell studies. Among these previous studies, Lactobacillus GG was shown to suppress cytokine-induced apoptosis in in vitro cultured epithelial cells. 30 By contrast, up-regulation of anti-apoptotic and cytoprotective genes was confirmed in the animal study that used orally administered Lactobacillus GG. 31 The repressive effect on apoptosis was observed in previous studies 32 and in the present study during the early stage. The induction of MAP kinases, observed in this study after the 20 h of treatment, was also detected in intestinal epithelial murine colonic YAMC cells after Lactobacillus GG treatment (32). Changes in gene expression for G-protein coupled receptor, cytochrome P450, and zinc finger protein were observed in the present study and was also previously reported. 17
Differential responses to probiotic bacteria between the present study and previous in vivo studies have been observed. In a previous study on the human intestinal tract, up-regulation of a G-protein coupled receptor was observed after 1 h of treatment in a clinical trial, but it was only up-regulated after 20 h of treatment in the present study. 10 In a human challenge trial, various genes linked to the immunoregulatory response were also changed. 10 In contrast, changes in the expression of genes involved in the immunomodulatory response were small in the present study. These results obtained in the previous in vivo studies may involve responses not only from epithelial cells but also from immune cells, such as dendritic cells and T cells. Moreover, treatment with Lactobacillus GG over a long period in the human challenge trial increased the expression of many immunomodulatory genes, such as cytokines and chemokines. 9 However, this was not observed in the present study. In contrast, increases in the gene expression of mucin, integrin, cytochrome P450, and ubiquitin observed in this in vitro study were also detected throughout the continuous uptake of GG in the previous study. 9 Moreover, human colon epithelial cells, such as T84 cells, HT29 cells, and Caco-2 cells, secreted pro-inflammatory and chemo-attractant cytokines in response to bacterial invasion treatment in a previous study. 33 However, the expression levels of cytokine genes were largely unchanged in the present study.
Many studies have focused on host GI responses to probiotic lactic acid bacteria, yet details of host-probiotic cross-talk are not fully understood. Probiotic strains are thought to specifically communicate with GI epithelial cells in a strain-dependent manner. Throughout the present transcriptome analysis of cultured Caco-2 cells, the host epithelial response to specific exposure with L-92 was mainly discussed to understand its impact on the GI tract. Our previous study, involving comparative proteome analysis with the L. acidophilus CP23 strain, which lacks SlpA, revealed that the binding of L-92 cells to Caco-2 cells might be strongly associated with the cell-surface SlpA protein (25). By attachment of the L-92 strain to the epithelial cells, most likely via the SlpA protein, Caco-2 cells seemed to respond to L-92 by accelerating cell proliferation after the first communication, during the early stage. It is also suggested that L-92 might have an in vivo effect on epithelial cells of the GI tract by specific binding. Information obtained in the present study could be used in considering the impact of L-92 on the GI tract and host health. In addition, comparative transcriptome analyses using different probiotic strains on Caco-2 cells would be of interest to help understand the potential of probiotics. Moreover, transcriptome analysis of immune cells, such as dendritic cells, should be addressed next in order to further understand gastric immunity.
Footnotes
Author Disclosure Statement
The authors confirm that no competing financial interests exist.