
Abstract
Conventional and newly emerging treatment procedures such as chemotherapy, catalytic therapy, photodynamic therapy, and radiotherapy have not succeeded in reversing the outcome of cancer diseases to any drastic extent, which has led researchers to investigate alternative treatment options. The extensive repertoire of traditional medicinal knowledge systems from various parts of the world are being re-investigated for their healing properties. It has been reported that several members of the Convolvulaceae, Iridaceae, and Liliaceae families have antitumor activity against some tumor cell lines. Here we first report that Convolvulus galaticus, Crocus antalyensis, and Lilium candidum species have cytotoxic activity on human breast cancer cell line MCF-7 cells. Plant samples were collected and identified, and their cytotoxic effects on the MCF-7 cell line were examined at different concentrations of methanol extracts. We found that all three plants have cytotoxic effects on MCF-7 cells but that C. galaticus has the strongest cytotoxic effect even in the lowest extract concentration tested (0.32 μg/mL). Our results indicate that these plant extracts have cytotoxic effects on human breast carcinoma cell line MCF-7 cells and that this cytotoxic effect comes from p53-mediated stimulation of apoptosis.
Introduction
B
Phytotherapy has been practiced since antiquity in Africa, Asia, Europe, and America. During the past 30 years, the use of herbs and related products has increased from 34% in 1990 to 42% in 1995. Throughout the centuries, several plant extracts have been tested for antitumor potential. Plants have provided many effective anticancer agents in current use such as irinotecan, taxanes, topotecan, vinblastine, vincristine, etc. 3 –5 Plant-derived products are excellent sources for the discovery and development of new anticancer agents. Moreover, plant materials represent promising sources of anticancer agents with lower side effects compared with chemical drugs.
Lilium candidum L. belongs to the Liliaceae family, which grows throughout Mediterranean region and western Asia. It probably originated from Syria and Persia. The essential oil, extracted from flowers (0.3%), is rich in vanillin (up to 2.5%), p-hydroxy-m-methoxytoluene (up to 50%), p-cresol, linalol, terpineol, phenylethyl alcohol, and its esters, with acetic, palmitic, benzoic, propionic, and cinnamic acids. 6 On the basis of usage in traditional medicine and on contemporary experimental experiences, some authors refer to the anti-inflammatory and healing effects of L. candidum L. Selected compounds isolated from the bulbs and flowers of L. candidum L. were investigated for potential anticarcinogenic activity. 6 L. candidum L. (Liliaceae), also called “white Madonna lily,” is well known in folk medicine for the treatment of burns, ulcer, and inflammations and for healing wounds. L. candidum L. extract contains various biologically active compounds. 6,7
Crocus species are members of the family Iridaceae. The plants in this family are herbs with rhizomes, corms, or bulbs. The family Iridaceae embraces about 60 genera and 1500 species. The genus Crocus includes native species from Europe, North Africa, and temperate Asia and is especially well represented in arid countries of southeastern Europe and Western and Central Asia. Crocus antalyensis, generally located in the Mediterranean region, is an endemic plant for Turkey. 8,9 There is no report on medicinal uses of C. antalyensis in the literature, but it is known that Crocus species are extensively used in traditional medicine for various purposes, as an aphrodisiac, antispasmodic, and expectorant, for treatment of stomach ailments, for reducing stomachache, and for relieving tension. 10 This study is the first to report antitumor activity of C. antalyensis.
There have been several chemical studies of both the family Convolvulaceae and the genus Convolvulus. Alkaloids have been reported from Convolvulus, 11 and acylated anthocyanis have been identified in Convolvulus. 12 There is also no report on medicinal uses of Convolvulus galaticus in the literature, but it is known that Convolvulus species are extensively used in traditional medicine for various purposes as in ulcer treatment, diabetes, and tension. 13
The aim of this study was to investigate the potential antitumor effects of L. candidum, C. antalyensis, and C. galaticus on human breast cancer cell line MCF-7 cells.
Materials and Methods
Plant materials
Plants were collected from around Turkey from different locations. C. galaticus was collected from around Kayseri, at an altitude of 1300 m above sea level. L. candidum was collected from around Mugla, at an altitude of 30 m above sea level. C. antalyensis was collected from around Antalya-Elmali, at an altitude of 750 m above sea level.
Plant extract preparation
After collection of plant samples, dried bulbs and leaves were chopped with a blender, and methanol extracts were prepared for the experiment. In this study, 10 g of the dried plant and 100 mL of solvent methanol (Merck, Darmstadt, Germany) were used for every sample. 14 The mixture was extracted after being heated in a vibrating water bath at 55°C. Having been acquired as a result of extraction, the mixture was filtered through filter paper (Whatman No. 1), and the solvents were evaporated in a rotary evaporator at 48–49°C. The water in each extract was frozen in a freeze-drying machine and drawn out.
Cell lines and culture conditions
MCF-7 cells were cultured in RPMI 1640 medium (Sigma Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (Invitrogen, Carlsbad, CA, USA) at 37°C in a humidified incubator with 5% CO2.
Cell proliferation assay
The medium was aspirated when MCF-7 cells grown to about 90% confluence. Cells were washed with phosphate-buffered saline, trypsinized, counted with a hemocytometer, and seeded into 96-well plates (3×104 cells/mL). After a 24-h incubation at 37°C in a 5% CO2 incubator, the medium was removed, and cells were treated with plant extracts added to the medium in different concentrations (0.3, 0.6, 1.25, 2.5, 5.0, 10.0, and 20.0 μg/mL) for 72 h. For the untreated control group, cells were not treated with any extracts. Plant extract concentrations were prepared in RPMI 1640 medium that included 10% fetal calf serum and then sterilized by passage through a filter (pore size, 0.2 μm). At the end of incubation periods, medium was removed, and cytotoxicity in plant extract-treated and untreated control groups was measured by the luminometric method using a CytotoxGlo® kit (Promega, Madison, WI, USA). Values for the concentration at which 50% inhibition occurred (IC50) were calculated for all three plant extracts.
Terminal transferase dUTP nick end-labeling apoptosis analysis
MCF-7 cells were trypsinized, counted with a hemocytometer, and then seeded into flasks (3×104/mL). For the detection of the induction of apoptosis of each plant extract, MCF-7 cells were treated with IC50 values of each plant extract (0.32 μg/mL for C. galaticus, 1.06 μg/mL for L. candidum, and 0.72 μg/mL for C. antalyensis) for 24 h at 37°C in a humidified incubator with 5% CO2. At the end of the incubation period late apoptotic events were analyzed by terminal transferase dUTP nick end-labeling (TUNEL) analysis using the In Situ Cell Death Detection Kit (Millipore, Billerica, MA, USA). Apoptotic cells were counted under the microscope.
Western blotting
At the end of the incubation period, MCF-7 cell lysates treated with IC50 values of each plant extract were prepared in ice-cold RIPA buffer (10 mM Tris-HCl [pH 7.5], 150 mM NaCl, 2 mM EDTA, 1% Nonidet P-40, 1% sodium deoxycholate, and 0.1% sodium dodecyl sulfate). Cellular debris was removed by centrifugation at 12,000 g for 5 min at 4°C. Proteins (100 μg) were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis using 2–15% gradient or 7.5% polyacrylamide gels (Pierce, Rockford, IL, USA). Proteins were immunoblotted onto Hybond™ polyvinylidene difluoride membranes (Amersham-PharmaciaBiotech, Piscataway, NJ, USA) and labeled with related antibodies. The p53 antibodies were obtained from Cell Signaling Technology (Danvers, MA, USA). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and horseradish peroxidase–labeled anti-mouse secondary antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Primary and secondary antibody labeling blots were treated with Super Signal West Pico chemiluminescent substrate (Pierce), exposed to Hyperfilm ECL™ (Amersham-PharmaciaBiotech), and developed.
Real-time reverse transcriptase–polymerase chain reaction analysis
Quantitative real-time reverse transcriptase (RT)–polymerase chain reaction (PCR) analysis was applied to determine relative mRNA levels in plant extract-treated and untreated control groups of MCF-7 cells. Cells were treated with IC50 values of each plant extract (0.32 μg/mL for C. galaticus, 1.06 μg/mL for L. candidum, and 0.72 μg/mL for C. antalyensis) for 24 h at 37°C in a humidified incubator with 5% CO2. At the end of the incubation time plant extract-treated and untreated control group cells were washed with ice-cold phosphate-buffered saline, and then total RNA from the MCF-7 cell line was prepared as described previously. 15 Expression of p53 and of its target genes for the proteins p21 and bax was assessed by quantitative real-time RT-PCR analysis using a QuantiTect® Probe PCR kit (Qiagen, Hilden, Germany) and an ABI Prism® 7900HT sequence detection system (Applied Biosystems, Foster City, CA, USA) according to the supplier's protocols. Sets of TaqMan® probe and primers were purchased from Applied Biosystems. A predeveloped TaqMan assay endogenous control of a GAPDH kit (Applied Biosystems) was used for amplification of a GAPDH cDNA fragment. The relative mRNA levels were calculated using the comparative C t method. Levels of p21, bax, and p53 transcript were evaluated. The transcript level of the GAPDH gene was used as the endogenous reference.
Statistical analysis
All experiments were performed in replicates of three and repeated independently to confirm the results. Significance of the differences in the means was determined using Student's t test and considering P<.05 to be statistically significant.
Results
For the detection of specific cytotoxic activity of each plant extracts, luminometric analysis was used as described in Materials and Methods. MCF-7 cells were incubated with seven different concentrations (0.3, 0.6, 1.25, 2.5, 5.0, 10.0, and 20.0 μg/mL) of total methanol extracts of C. galaticus, C. antalyensis, and L. candidum for 72 h. Figure 1 clearly indicates that total methanol extracts of C. galaticus, C. antalyensis, and L. candidum have cytotoxic effects on MCF-7 cellular proliferation. C. antalyensis and C. galaticus have a stronger cytotoxic effect than L. candidum (Fig. 1). We also calculated IC50 values for each plant extract from Figure 1 and represented in Table 1. According to Table 1, C. galaticus extract has the lowest IC50 values (0.32 μg/mL) compared with C. antalyensis (0.72 μg/mL) and L. candidum (1.06 μg/mL); the IC50 values are in good agreement with our cytotoxicity results.
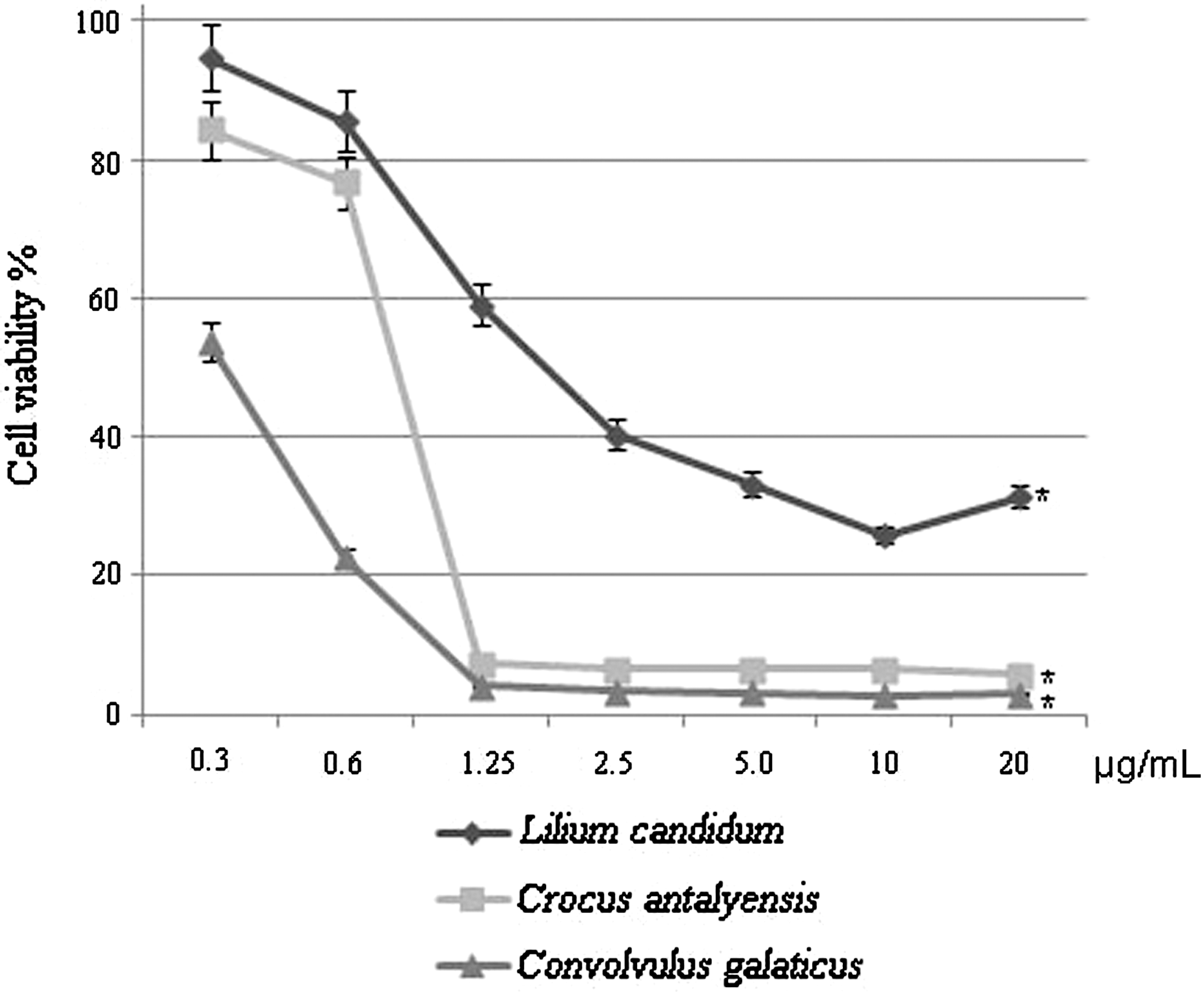
Convolvulus galaticus, Crocus antalyensis, and Lilium candidum extracts have cytotoxic effects on MCF-7 cells. Human breast carcinoma cell line MCF-7 cells were seeded at a density of 3×104/mL in 96-well plates. After 24 h, the cells were washed with phosphate-buffered saline, fresh growth medium was added, and then cells were treated with C. galaticus, C. antalyensis, and L. candidum extracts in different concentration for 72 h. At the end of the incubation, cell viability was determined in plant extract-treated and untreated control groups by the luminometric method. Data are mean±SD values (n=3). *P<.001 by Student's t test for cytotoxic effects of plant extract compared with the untreated control group.
50%
IC50, concentration at which 50% inhibition occurs.
Because extensive cell death was observed in proliferating MCF-7 cells after treatments with the plant extracts, we have investigated if the plant extract treatment-induced cell death occurred through apoptosis. MCF-7 cells were incubated with IC50 values of total methanol extracts of C. galaticus, C. antalyensis, and L. candidum for 24 h. At the end of the incubation time, cells were harvested and assayed for induction of apoptosis with use of the TUNEL method. The TUNEL assay was developed as a method to identify individual cells that were undergoing apoptosis by labeling the ends of degraded DNA with the polymerase terminal deoxynucleotidyl transferase. Figure 2 shows that C. galaticus, C. antalyensis, and L. candidum extracts have the ability to induce apoptosis on MCF-7 cell lines. Among the three plant extracts C. galaticus has the biggest effect on induction of apoptosis on MCF-7 cells, which is positively related to the cytotoxic effects of the plants.
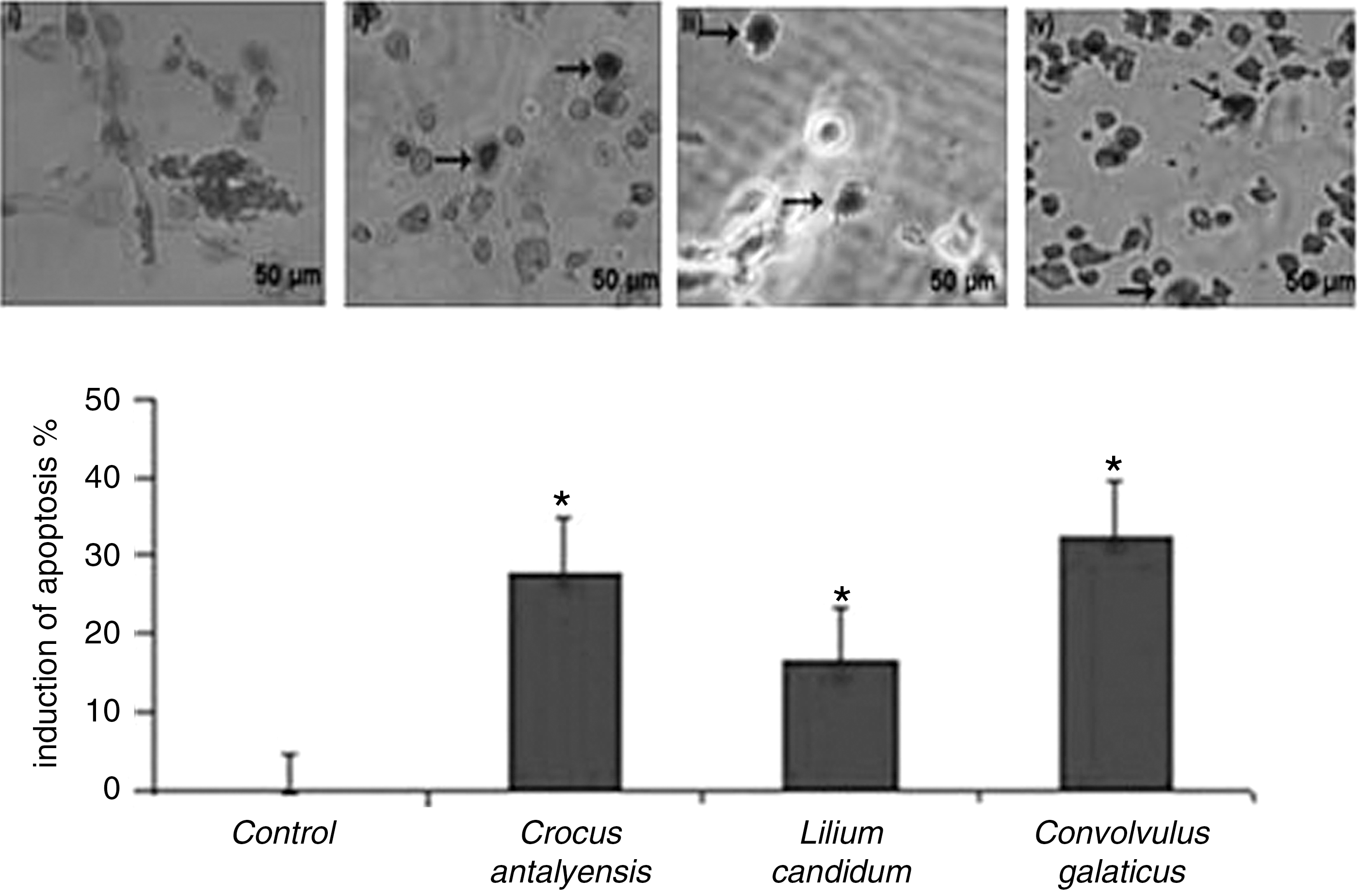
C. galaticus, C. antalyensis, and L. candidum extracts induce apoptosis in MCF-7 cells. Human breast carcinoma cell lines MCF-7 cells (3×104/mL) were incubated with IC50 values of C. galaticus, C. antalyensis, and L. candidum extracts for 24 h. After the end of the incubation time cells were washed with phosphate-buffered saline and then assayed by terminal transferase dUTP nick end-labeling analysis using an In Situ Cell Death Detection Kit (Millipore) system to indicate cellular apoptosis:
The p53 tumor suppressor gene regulates cell cycle progression and cell survival in response to cellular stress. DNA damage or oncogenic stress induces p53 protein levels, allowing elimination of incipient tumor cells by apoptosis. To determine if the plant extracts can induce apoptosis through p53, we detected p53 protein expression by western blotting in the plant extracts of treated samples. Western blot analyses showed that levels of tumor suppressor p53 protein were significantly increased after the 24-h incubation with C. galaticus, C. antalyensis, and L. candidum extracts in MCF-7 cells (Fig. 3). The results indicate that these plant extracts induce apoptosis via induction of cellular accumulation of p53 in the MCF-7 cell line. Because tumor suppressor p53 is a transcriptional activator, we want to examine the effects of p53 expression on its target genes, p21 and bax. Therefore, we analyzed p53, p21, and bax mRNA expressions in plant extract-treated and untreated MCF-7 cells with quantitative real-time PCR. Figure 4 clearly shows that levels of p53 and its target genes p21 and bax mRNA increased after all three plant extract treatments.
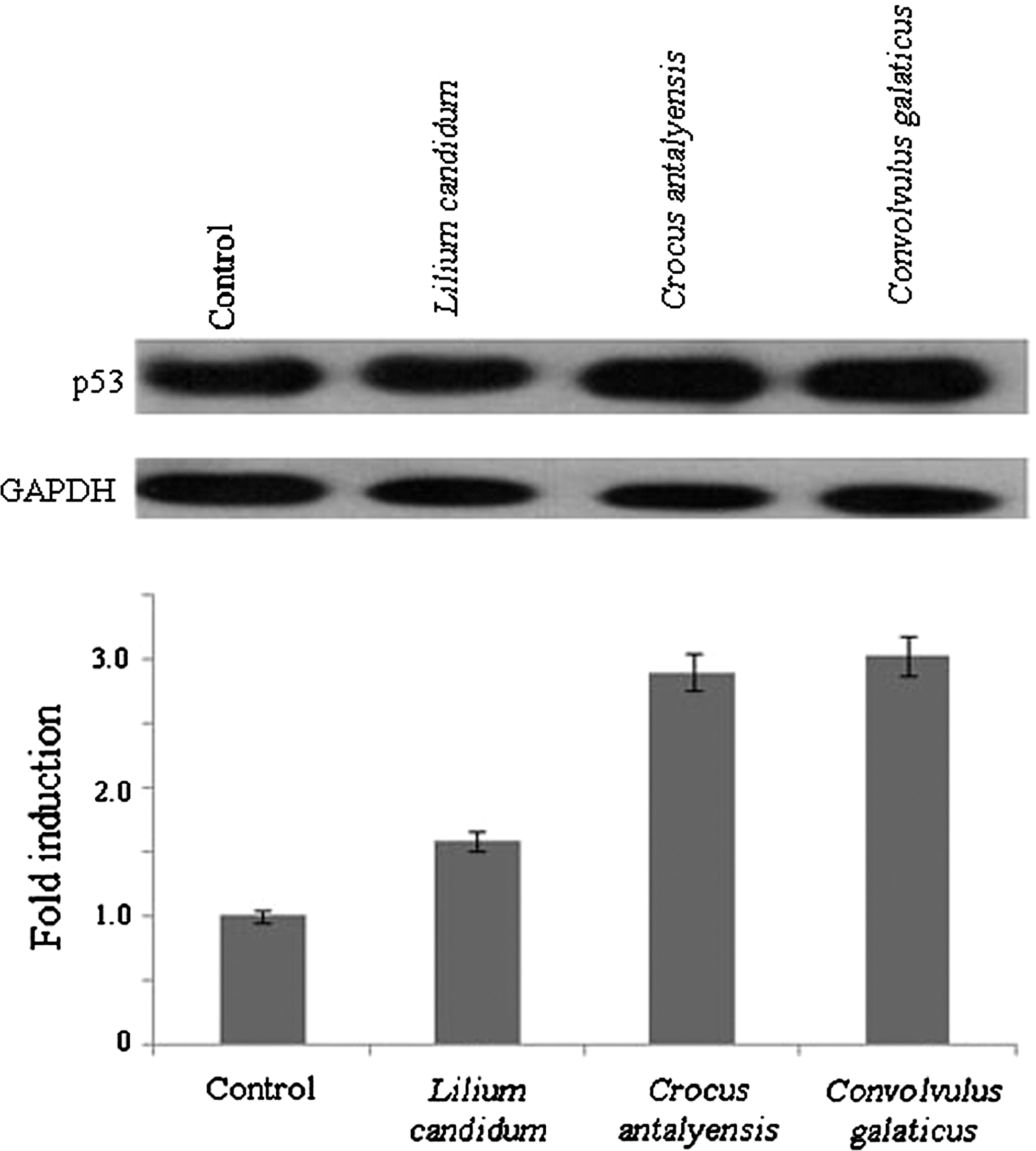
C. galaticus, C. antalyensis, and L. candidum extracts induce accumulation of p53. MCF-7 cells were cultured in RPMI 1640 medium with 10% fetal calf serum and incubated with C. galaticus, C. antalyensis, and L. candidum extracts with IC50 values for 24 h. Cell lysates were fractioned on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, and western blots were probed with antibody directed against human p53, striped off, and reprobed with anti-human glyceraldehyde 3-phosphate dehydrogenase (GAPDH) antibody for even loading. Results are representative of three independent experiments. Densitometric analysis was also performed. Band intensities of p53 were normalized against that of GAPDH to determine fold increase in the amount of p53.
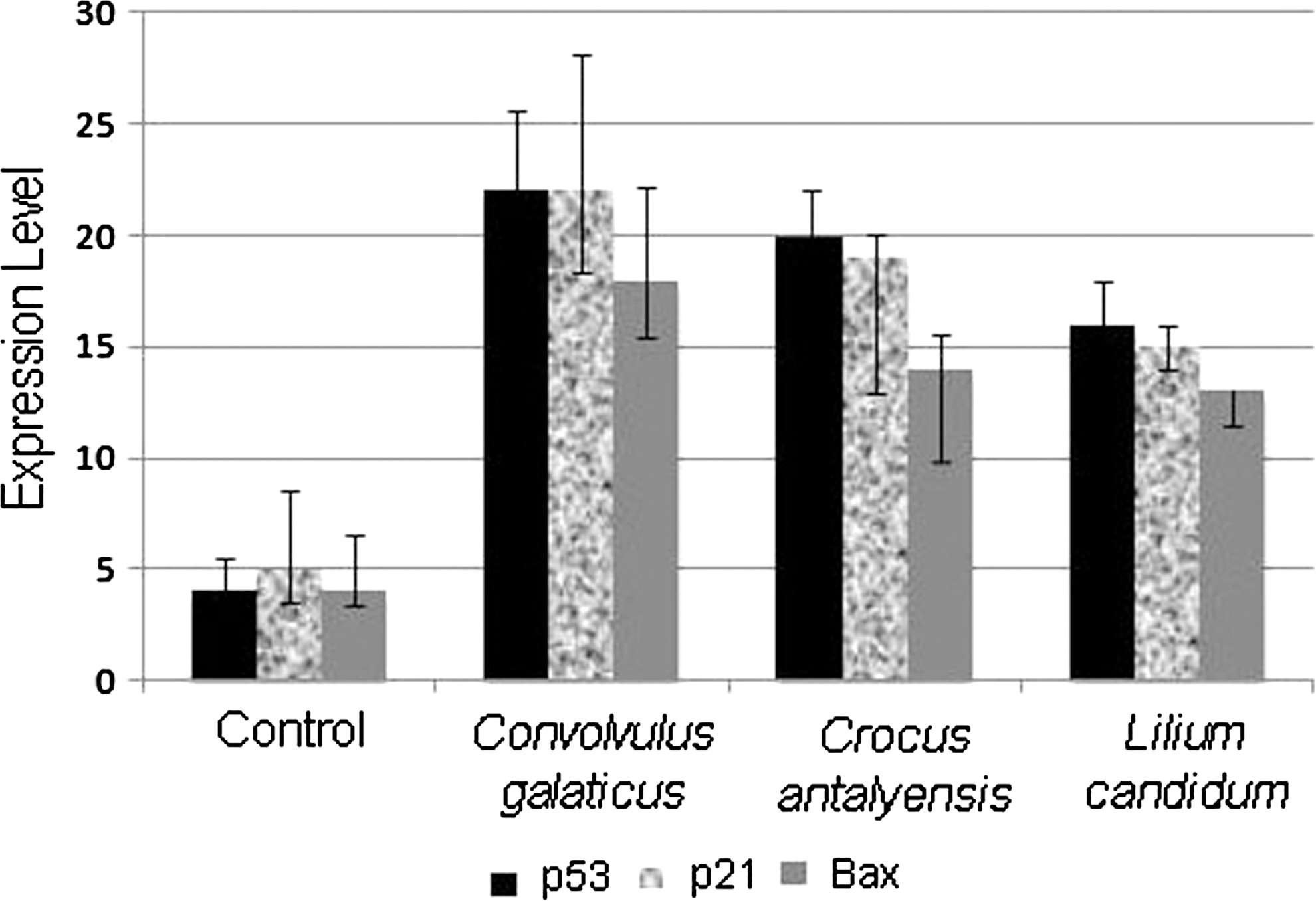
C. galaticus, C. antalyensis, and L. candidum extracts induce p53, p21, and bax mRNA expressions. Data are mean±SD values of three analyses by quantitative reverse transcriptase–polymerase chain reaction.
Discussion
Cancer remains one of the leading causes of death around the world. Breast cancer is the second highest incidence in the world after lung cancer. Chemotherapy is the most common and effective method for the treatment of breast cancer. Various cancer therapies have currently been examined, including the use of natural products from plants. The need to develop more effective and less toxic anticancer drugs has prompted researchers to explore new sources of pharmacologically active compounds. Natural products have long been used to prevent and treat diseases, including cancers, and might be good candidates to develop anticancer drugs. 16 Plants have substantial potential for the discovery of active anticancer compounds as most chemotherapeutic drugs, like taxol and vincristine, were already isolated from plants. Therefore, attempts to search for plant-derived active compounds for new anticancer treatments might be promising.
The present study was designed to investigate the potential therapeutic abilities of C. galaticus, L. candidum, and C. antalyensis extracts in human breast cancer cell line MCF-7 cells. Before this study, there is no report on these plant species on anticancer abilities in the literature. This is the first to report that C. galaticus, L. candidum, and C. antalyensis have strong cytotoxic effects on human breast cancer cell line MCF-7 cells in a dose-dependent manner. Among the extracts of the three plants, C. galaticus extract has the strongest cytotoxic effects on MCF-7 cells in a dose-dependent manner. We also detected strong cytotoxic effects of the extracts of C. antalyensis, which is an endemic plant in Antalya, Turkey, on MCF-7 cells. It was reported that saffron, which also belongs to the Crocus genus, can induce apoptosis in MCF-7 cells 17 and in HeLa and HepG2 18 cell lines. Our results are in good agreement with previously published results. Anticancer effects of C. antalyensis are also observed in other species of the Crocus genus as reported for saffron. Also, some reports were published on the anticancer effects of Lilium species on human cancer cell lines, 7,19 but until our study, there has been no report on the antitumor activity of L. candidum on any human cancer cell lines in the literature.
Apoptosis is an active physiological process resulting in cellular self-destruction that involves specific morphological and biochemical changes in the nucleus and cytoplasm. 20 –22 Agents that suppress the proliferation of malignant cells by inducing apoptosis may represent a useful mechanistic approach to both cancer chemoprevention and chemotherapy. 23 As many anticancer agents have been developed, unfavorable side effects and resistance become serious problems. 24 Thus, there is growing interest in the use of plant materials for the treatment of various cancers and the development of safer and more effective therapeutic agents. 25 p53 is an extremely efficient inhibitor of cell growth, inducing cell cycle arrest and/or apoptotic cell death, depending on cell type and environment. 26 Therefore, regulation of p53 activity is critical to allow normal cell division. The tumor-suppressive function of p53 must be dampened sufficiently to allow normal growth and development. There are many mechanisms through which p53 is regulated. 27 The major mechanisms include regulation of p53 protein levels, control of the localization of p53 protein, and modulation of the activity of p53, in particular its ability to function as a sequence-specific transcription factor. 28,29 We detected with western blot analysis that C. galaticus, L. candidum, and C. antalyensis total extracts lead to increased p53 protein accumulation (Fig. 3). Because we want to examine whether p53 accumulation comes from increased p53 transcriptional activity or not, the effects of the three plant extracts on p53 mRNA expression were also examined by quantitative real-time PCR. Our results indicate that total extracts of C. galaticus, L. candidum, and C. antalyensis induced p53, p21, and bax mRNA expressions in human breast carcinoma cell line MCF-7 cells. p53 is a transcriptional activator, and it is well known that p53 can induce expression of apoptosis-mediator genes. 30 Hastak et al. 30 reported that p53 can also induce the expressions of p21 and bax mRNAs. Our results are in good agreement with previously published results.
Here we report that C. galaticus, L. candidum, and C. antalyensis total extracts have strong cytotoxic effects on cellular proliferation, and this cytotoxicity resulted through the p53-mediated apoptosis. However, it has been reported that p53 gene mutation rate is 20–30% in breast cancer, 31 –33 which means that p53 is in its wild-type form in 70–80% of breast carcinoma patients. 33 p53-mediated apoptosis is a very important pathway to kill the breast carcinoma cells. Because oncologists have very limited weapons for the battle against breast cancer, it is very important to find new compounds that potentially can be used as a new drug on cancer treatment. Our results did not show that C. galaticus, C. antalyensis, and L. candidum total extracts can be used as a treatment against breast carcinoma, but clearly indicate that these three plant extracts potentially include an chemical compound or compounds against breast cancer cells. Therefore, these compounds still need to be isolated and identified in future studies.
Conclusions
Our data indicate that C. galaticus, C. antalyensis, and L. candidum have strong anticancer effects on human breast cancer cell line MCF-7 cells and that these effects come from induction of apoptosis through accumulation of p53. Our data indicate that these three plant species potentially may include active compounds that may be improved as a therapeutic agent for human breast cancer, after isolation and identification studies in the future.
Footnotes
Author Disclosure Statement
No competing financial interests exist.