
Abstract
Valproic acid (VPA) is a potent inducer of neural tube defects in human and mouse, its teratogenicity is associated with its potential to generation of free radicals and increase oxidative stress. Furthermore, spirulina (SP) has shown pharmacological properties against teratogenicity, which are attributed to its antioxidant potential. Accordingly, the present study was performed to investigate the influence of SP on the teratogenicity of VPA in imprinting control region mice and the possible mechanisms of action. VPA (sodium valproate) was administered intraperitoneally to mice on gestation day (GD) 8 at a dose of 600 mg/kg. SP was given orally at 125, 250, and 500 mg/kg daily from GD0 through GD18. The most common finding in fetuses with VPA exposure was exencephaly. SP decreased the incidence of this and other malformations and increased levels of superoxide dismutase, catalase, and glutathione peroxidase. In conclusion, these results illustrate the protective action of SP through its antioxidant activity against VPA-induced teratogenicity.
Introduction
V
The molecular mechanism by which VPA causes NTDs has yet to be completely clarified. In general, with prolonged use of antiepileptic drugs, it has been observed that there is an increase in the formation of free radicals, which cause oxidative damage to neurons and DNA, and a decrease in the synthesis of antioxidant enzymes, such as glutathione peroxidase (GPx), superoxide dismutase (SOD), and catalase. 3
VPA is metabolized to 2-ene VPA and E2-2,Z-3-diene VPA, which is highly reactive and binds to embryonic tissues and/or fetal DNA, causing a disruption in fetal growth and development. 4
Spirulina (SP) is a cyanobacteria (blue-green alga, primarily Arthrospira platensis and Arthrospira maxima) with a high content of proteins, vitamins, minerals, amino acids, and essential fatty acids, including glucolipids and sulfolipids, 5 which make it an important source of nutrients. Additionally, several pharmacological properties of SP have been reported, including anti-inflammatory, neuroprotector, and antigenotoxic. 6 Some of these effects could be due to the antioxidant activity of SP per se, or to that of some of its extracts. 7 Among the phytochemical extracts are phycobiliproteins, such as the antioxidant agent phycocyanin, which corresponds to 14% of the dry weight of SP. 8,9 It has also been proposed that SP increases the activity of antioxidant enzymes like SOD. 10
In a study conducted by Paniagua-Castro et al., 11 it was shown that SP provided to mice during the entire gestation period prevented the teratogenic effect of cadmium administered on day 8 of pregnancy. Kapoor and Mehta 12,13 showed that SP prevented maternal blood disorders during gestation and lactation, and that the offspring had higher birth weight compared with controls.
The aim of the present study was to evaluate whether SP protects against valproate teratogenicity when given before neurolation in ICR mice, and if so, the relation between this protection and its antioxidant effect.
Materials and Methods
Test substances
VPA sodium salt (2-propyl-pentanoic acid sodium salt) was of analytical grade and was purchased from Sigma Chemical Co. (St. Louis, MO, USA). The SP sample was from a bulk production batch (5DW-9714) of standardized quality, supplied by Alimentos Esenciales para la Humanidad, S.A. de C.V. (Mexico City, DF, Mexico). The alga was identified by Prof. Catalina Mendoza González at the ENCB Herbarium of the National School of Biological Sciences, National Polytechnic Institute (Mexico City), where a voucher herbarium specimen (12584) is kept.
Animals
ICR mice (Harlan de México, Mexico City) of both sexes, weighing 25–30 g, were kept in a room at a constant temperature of 22°C±1°C and on a 12-h light–12-h dark cycle (lights on at 9:00 a.m.), with given food and water provided ad libitum. All experiments were approved by the Laboratory Animal Care Committee of our Institution and were conducted in compliance with the Mexican Official Standard (NOM ZOO–062-200-1999) regarding technical specifications for production, care, and use of laboratory animals.
Experimental design
The study was conducted under a completely randomized design open to the observer. One male mouse was housed with four nulliparous females from 6:00 to 9:00 a.m. (the last 3 h of the dark cycle). The day a vaginal plug was observed in females was designated as gestational day (GD) 0, and mice in this condition were separated.
Pregnant mice were randomly divided in six groups of 10 animals, as shown in Table 1.
SP, spirulina; VPA, sodium valproate.
SP suspended in saline was given to mice by the oral route during the entire gestational period (GD 0–17). The doses were selected on the basis of previous studies, where it was demonstrated that the highest dose was very well tolerated by the mice when administered by this route. 14 VPA (2 propylpentanoic acid sodium salt, Sigma) was dissolved in saline and injected intraperitoneally (i.p.) in doses of 600 mg/kg on GD 8, 1 h after SP administration. Pregnant mice were weighed on GD 0, 6, 8, 11, and 17.
Teratological study
On GD 17, female mice were sacrificed, and fetuses were immediately removed and weighed, sexed, and inspected for external malformations. Total implantation sites and reabsorptions were noted in the uterus. Two-thirds of viable fetuses in each litter were fixed in 95% ethanol and skeletal staining, following Peters' double-staining method. 15 The remaining fetuses were fixed in Bouin's solution and prepared for serial slices as described by Wilson. 16
Lipid lipoperoxidation and antioxidant enzyme activity
Pregnant mice were randomized into four groups of four dams each: (1) control, (2) SP 500 mg/kg, (3) VPA (2 propylpentanoic acid sodium salt; Sigma) 600 mg/kg, and (4) VPA 600 mg/kg+SP 500 mg/kg. SP was administered from GD 0 to GD 10 by the oral route; VPA was injected i.p. on GD 8. The animals were sacrificed on GD 10 and a hysterectomy was performed to obtain the embryos. Three embryos (from each litter) were homogenized and tested for lipid peroxidation by measuring the micromolar hydroperoxide concentration with a hydroperoxide kit (Sigma), catalase activity (catalase assay; Sigma), SOD activity (RANSOD; Randox, Crumlin, United Kingdom), as well as GPx activity (RANSEL; Randox) and protein content by the Coomassie blue method 17 using bovine serum albumin as a standard.
Statistical analysis
Litter/dam was used as the basic unit for statistical analysis. The χ 2-test followed by Fisher's exact test was used to evaluate fetuses with abnormalities as well as the type of external and skeletal anomalies. The remaining variables were assessed by one-way analysis of variance (ANOVA), and then by the Student–Newman Keuls test (post hoc test), or ANOVA on ranks when the normality test failed. A value of P<.05 was considered significant.
Results
The disorders induced by the VPA treatment (Table 2) include a significant increase in the number of reabsorptions and of fetuses with exencephaly, as well as a decrease in the number of fetuses. The SP treatment diminished the number of fetuses with exencephaly in a dose-dependent manner.
Data are mean±SE. Mice were treated with Spirulina (SP) 125, 250, or 500 mg/kg and sodium valproate (VPA) 600 mg/kg, n=10 for each treatment.
Significantly different (P<.05) from *control, †VPA 600 mg/kg, #SP 125 mg/kg+VPA 600 mg/kg.
Skeletal anomalies induced by the VPA treatment (Table 3) include lack of ossification in the head, sternebrae, vertebrae, and phalange regions, as well as fusion of the ribs and the appearance of the 14th rib. Cotreatment with SP diminished the number of fetuses with such alterations in a dose-dependent manner, the effect being significant (P<.5) at the doses of 250 and 500 mg/kg.
Mice were treated with Spirulina (SP) 125, 250, or 500 mg/kg and sodium valproate (VPA) 600 mg/kg.
Parietal and occipital bones.
Significantly different (P<.05) from *control, †VPA 600 mg/kg, #SP 125 mg/kg+VPA 600 mg/kg.
The visceral malformations induced by VPA include hydrocephaly, exophtalmos, micrognathia, hydronephrosis, and open eye (Table 4). SP significantly diminished the frequency of these alterations in a dose-dependent manner.
Mice were treated with Spirulina (SP) 125, 250, or 500 mg/kg and sodium valproate (VPA) 600 mg/kg.
Significantly different (P<.05) from *control, †VPA 600 mg/kg, #SP 125 mg/kg+VPA 600 mg/kg.
At GD 10, mice with VPA treatment only (administered on GD 8) were found to have a significant increase in the lipid hydroperoxide content as well as a decrease in antioxidant enzymes, including SOD, GPx, and catalase in embryos (Fig. 1). Cotreatment with SP prevented both this increase in oxidation and the decrease in antioxidant enzyme activity.
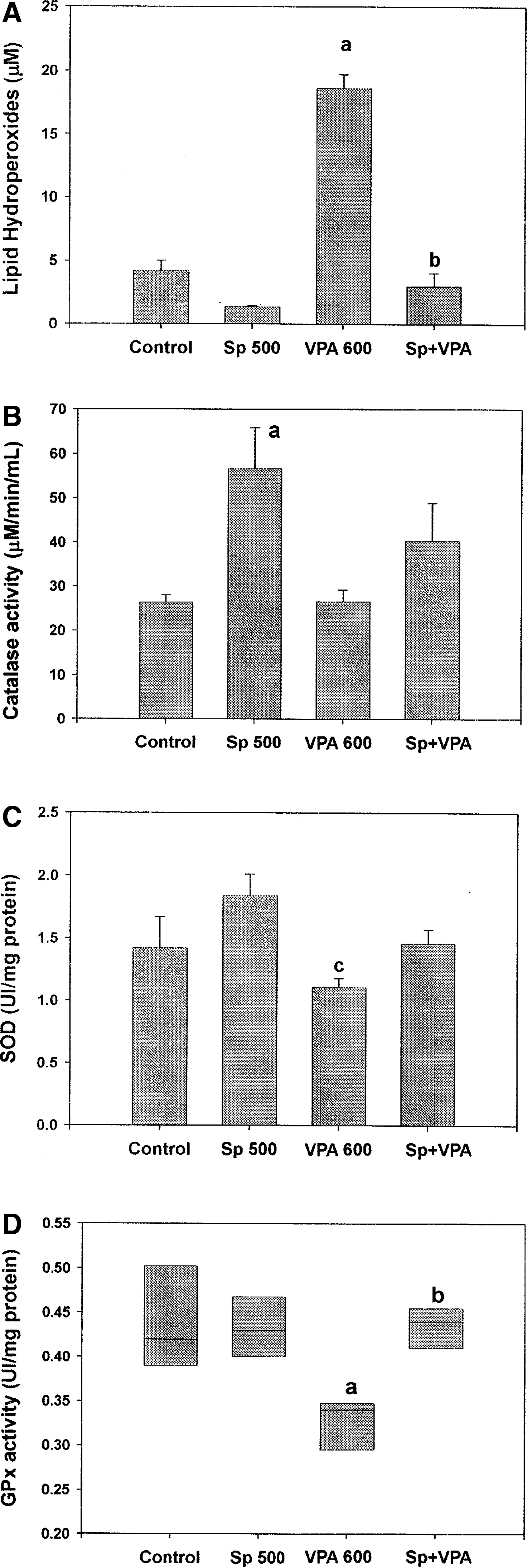
Determination of the antioxidant activity of spirulina (SP) on the embriotoxicity of sodium valproate (VPA) on mouse embryos:
Discussion
Children exposed in utero to VPA have a high risk of suffering NTDs and a syndrome that includes malformation of extremities, heart, liver, skull, and face, as well as a cerebral dysfunction with mental retardation related in some cases with autism. 18 However, the precise mechanisms that generate these disorders are not completely understood.
In the present study, it was found that VPA caused exencephaly and facial and skull malformations, such as open eye, exophtalmos, and micrognathia, which coincided with the effects reported in other studies. 4,19 Skeletal malformations were also found in the fetuses, principally in the vertebrae, sternebrae, and ribs, which also coincide with the results of other studies. 20
It is known that VPA can pass through the placenta and reach the embryo, 21,22 where it remains for ∼45 h. 23 Among other effects, this gives rise to at least two important factors in embryonic malformations: (1) the lack of expression of embryonic proteins necessary for both the closing of the neural tube and the defense against pro-oxidant agents, and (2) the generation of free radicals leading to the oxidation of molecules that are vital to embryonic development. 24 Dawson et al. 25 reported a decrease in NF-kB, Pim-1, and Myb proteins in embryos with NTD. Particularly noteworthy is that a lack of NF-kB produces cellular apoptosis in the area of the neural tube. It is known that NF-kB activates the antiapoptotic protein Bcl-2, whose action is antagonistic to the proapoptotic protein Bax. Therefore, the authors propose that the decrease of NF-kB induced by VPA causes a series of reactions in cascade that result in the diminishment of Bcl-2, which in turn leads to the generation of cellular apoptosis.
Regarding antioxidant proteins, it has been reported that the human embryo in the stage of 4 to 8 cells, and the mouse embryo in the stage of 2 cells have a low content of antioxidant enzymes, which normally provide antioxidant defenses. Such antioxidant enzymes include ascorbate, alpha-tocoferol, SOD, glutathione, GPx, catalase, and metalotioneins. 26 Various studies have reported that VPA causes a decrease in GPx, 26 SOD, 27 and glutathione. 28
VPA has also been associated with the generation of free radicals, oxidative stress, and NTDs. Embryos are highly susceptible to an increase in ROS production during their development, especially after the formation of the circulatory system and the resulting increase in aerobic metabolism. 29 This effect mainly owes itself to the fact that the tissues have a low activity of antioxidant enzymes and a high content of free radicals of oxygen and nitrogen, generating oxidative stress and embryonic damage. 30 Furthermore, it has been demonstrated that the brain is more susceptible to oxidative stress in comparison to other fetal organs, 31 resulting in the production of NTDs.
Various studies have demonstrated that treatment with antioxidant agents, such as alpha tocoferol and N,N-diphenyl-D-phenylendiamine, completely protects against hepatotoxicity and renal damage 32 induced by VPA. It has also been observed that vitamin E and pantotenic acid reduce fetal malformations induced by VPA, probably due to their antioxidant activity. 4
In a previous study, we reported that SP diminished malformations induced by CdCl2, as well as lipid hydroperoxides in 10-day-old embryos. 14 It is known that SP is an important source of protein, vitamin E, antioxidant enzymes SOD and GPx, and DE phycobiliproteins, among other components. 33 Phyicocyanin, the biliprotein with the greatest concentration, has demonstrated a high antioxidant activity in different experiments, 8,34 as well as a potent inhibitory effect on NADPH oxidase. 35 These factors are probably the major contributors to the potent antioxidant effect of SP.
In the present study, SP diminished the number of fetuses that presented malformations induced by VPA. It also decreased the levels of hydroperoxides and increased the activity of the antioxidant enzyme GPx, which suggests that its protective effect is due to its components with antioxidant action.
There is extensive interest in the progress of strategies to protect development against insults during organogenesis. Although maternal dietary antioxidant supplementation has clearly been successful in improving fetal outcomes in animal models, such as experimental diabetes 36 or after exposure to specific teratogens, 37 the available evidence of the protective ability of functional foods with antioxidant capacity against chemical teratogenesis is limited. However, it has been proved in rats that the concomitant oral administrations of garlic extract (500 mg/kg/day for 5 days/week or vitamin C (twice as the human recommended therapeutic dose of 10 mg/kg b.wt. for 5 days/week) to pregnant females on gestation days 6–15 provided significant reduction in the percentage of the fetal malformations induced by the insecticide cypermethrin. 38 These results together with those presented here are consistent evidence of potential benefit from the presence of antioxidant agents in the diet against adverse pregnancy outcomes.
We propose that treatment with SP during pregnancy may help prevent NTDs and bone malformations associated with VPA treatment. However, more studies are necessary to extrapolate these results to humans.
Footnotes
Acknowledgment
We thank Bruce Allan Larsen for reviewing the use of English in this manuscript.
Author Disclosure Statement
No competing financial interests exist.