
Abstract
Polyphenols have been demonstrated to have clear antioxidant activities in vitro. However, in complex biological systems, they exhibit additional properties, which are yet poorly understood. The apple is among the most consumed fruits worldwide, and several studies suggest that apple polyphenols could play a role in the prevention of degenerative diseases. The present study aimed at evaluating the Annurca apple polyphenol extract (APE) effects both proliferation and apoptosis on HaCaT cells. The data indicate that apple polyphenolic compounds had significant antiproliferative action on HaCaT cells. The fluorescence-activated cell-sorting analysis showed that APE induced cell apoptosis in a dose-dependent manner. Moreover, apple polyphenols induced apoptosis in epithelial cells by triggering a death receptor-associated extrinsic pathway p53-independent. APE was also capable of inducing morphological changes as evidenced by nuclear condensation. The cellular, morphological, and molecular data unequivocally demonstrated that induction of cellular apoptosis was mainly responsible for the previously observed antiproliferation-induced APE on HaCaT keratinocytes. Our experimental results suggest that apple polyphenols are a promising source from which a natural-based topical agent could be developed for skin diseases treatment.
Introduction
P
The Annurca is an apple variety widely cultivated in southern Italy. The main classes of polyphenols in apple are flavonoids, including quercetin, (−)-epicatechin and (+)-catechin, procyanidins, and anthocyanidins; dihydrochalcones such as phloretin and phloridzin; and other phenolic compounds such as chlorogenic acid. Apple polyphenols have shown high antioxidant capacity in vitro, 6,7 and it has been reported that the consumption of apple juice increases the antioxidant status of blood. 8 Furthermore, it has been shown that apple polyphenols inhibit the proliferation of cancer cells 9 and also exhibit an antiarteriosclerosis activity by reducing low-density lipoprotein oxidation. 10 A dose-dependent antiproliferative activity on colon as well as liver cancer cells has been observed using extracts from fresh apples. 11
There is a steadily growing interest nowadays in the use of natural products, as polyphenols, to combat several skin diseases such as psoriasis, 12,13 a common chronic inflammatory skin disease that affects, at varying degrees of severity, ∼1%–3% of the population worldwide. 14,15 Indeed, many commonly prescribed antipsoriatic drugs such as dithranol, methotrexate, and vitamin D3 analogs exert their therapeutic actions through counteracting keratinocyte hyperproliferation and/or modulating keratinocyte differentiation. 16 –18 Compounds that inhibit keratinocytes proliferation and modulate their differentiation are potentially useful in the treatment of psoriasis, because a balanced homeostatic control of growth and differentiation of this cellular line is crucial for recovery from psoriatic to normal epidermis. Traditional Chinese medicine has been widely used for the treatment of psoriasis in China, 19,20 and data derived from clinical investigations have indicated that Chinese herbal medicines are effective and safe in the management of psoriasis. Moreover, tea extracts have been traditionally used in alternative medicine as anticarcinogenic/antibiotic agents or in the treatment of conditions such as psoriasis. 21,22
The preset study aimed to investigate the effects of polyphenol extracts from Annurca apple fruits on the proliferation of keratinocytes using cultured HaCaT cells as a psoriasis-relevant experimental model.
Materials and Methods
Materials
The Dulbecco's modified Eagle's medium (DMEM) with 4.5 mg/mL glucose, fetal calf serum (FCS), nonessential amino acids, N-(2-hydroxyethyl) piperazine-N-2-ethanesulfonic acid, and phosphate-buffered saline (PBS) tablets were purchased from Gibco Life Science Technologies. Folin–Ciocalteu reagent, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazoliumbromide (MTT), propidium iodide (PI), sodium carbonate, and trypan blue 0.5% were purchased from Sigma Chemical Co. Rabbit polyclonal anti-caspase-3, rabbit polyclonal anti-caspase-8, rabbit polyclonal anti-caspase-9, and rabbit polyclonal anti-p53 were purchased from Santa Cruz Biotechnology.
Polyphenol extraction
Forty grams of Annurca apple flesh were homogenized for 5 min by a Tefal Rondo 500 homogenizer using 40 mL of 80% methanol and 20% water plus 0.18 N HCl (15 mL 12 N of HCl/L). After centrifugation (18,000 g for 25 min), the slurry was dried under vacuum by using the Univapor Concentrator Centrifuge, model Univapo 100 H (Uni Equip). The dried extracts were dissolved in 10 mL of PBS and frozen at −80°C until an analysis. The chemical characterization was performed in HPLC as reported in D'Angelo et al. 6 All experiments were carried out in triplicate.
Polyphenolic content
The total polyphenolic content of apple extracts was assessed approximately by using the Folin–Ciocalteu phenol reagent as described in Singleton et al. 23 The extracts (100 μL) were mixed with the Folin–Ciocalteu phenol reagent (0.5 mL), deionized water (0.9 mL), and Na2CO3 (7.5% w/v, 4 mL). The absorbance at 765 nm was measured 1 h after incubation at room temperature using a Cary ultraviolet–visible spectrophotometer (Varian). The measurement was compared to a standard curve of prepared catechin solutions and expressed as milligrams of catechin equivalent per 100 g (SD apple component).
Cell culture
HaCaT cells, an immortalized cell line of human epidermal keratinocytes 24 have been extensively used as an in vitro model for studies on the pathogenesis of psoriasis and evaluation of antipsoriatic drugs. 25 –27 This cell line was routinely grown at 37°C, in a humidified incubator with 5% CO2/95% air atmosphere, in the DMEM, supplemented with 10% FCS, glutamine (2 mM), penicillin (100 units/mL), streptomycin (100 units/mL), and 1% nonessential amino acids. All cell culture experiments were carried out when the culture was 60%–90% confluent.
Proliferation assay
The apple polyphenol extract (APE) together with HaCaT cells was cultured in six-well plates, with each well containing 2×105 cells in 2 mL of DMEM. By serial dilution, the final concentration of APE ranged from 100 to 500 μM. The treated HaCaT cells were incubated for 24 and 48 hours, and the proliferation rates under the influence of polyphenol extract were determined by the MTT assay. 28
In brief, after APE-treatment, 100 μL of the DMEM, containing 5 mg/mL MTT, was added to each well (1 mL of the final medium). Three hours later, the DMEM–MTT solution was removed, and the converted dye was solubilized with acid isopropanol (0.1 N HCl in isopropanol). The resulting absorbance from each well was measured at a wavelength of 570 nm with background subtraction at 630 nm. 28
The number of cells surviving to the treatment with APE was also evaluated by vital cell count in Trypan Blue (0.5% solution) in a Burker chamber.
Evaluation of apoptosis by DNA-flow cytometry
For apoptosis evaluation, subconfluent cells were treated, for 24, by adding various amount of APE (100, 200, 250, and 500 μM). Apoptotic cells, which appeared as floaters in the culture medium, were harvested by centrifugation at 1000 g for 5 min, whereas adherent cells remaining on the culture dish and control cells were trypsinized, and subsequently collected by centrifugation.
This assay is based on PI (from Sigma Chemical Company) staining of cells followed by the flow cytometry analysis, as described in literature. 29 Briefly, HaCaT cells (106) were incubated for 4 h at 4°C in 2 mL hypotonic solution containing 50 μg/mL PI, 0.1% sodium citrate, 0.1% Triton X-100, and 20 μg/mL DNase-free RNase A. The stained cells were analyzed, for relative DNA content, on a FACScan flow cytometer (Becton Dickinson). Cells with subdiploid DNA content (sub-G0/G1 peak) were considered apoptotic cells. All experiments were done in triplicate.
Western blotting analysis
For the Western blot analysis, HaCaT cells treated with APE as described above were lysed at 4°C for 1 h in a lysis buffer (50 mM Tris/HCl, pH 7.5, 150 mM NaCl, and 1% TRITON X-100), supplemented with a cocktail of phosphatase and proteinase inhibitors (1 mM sodium ortho-vanadate, 1 mM phenylmethylsulfonyl fluoride, 10 mg/mL leupeptin, 10 mg/mL pepstatine, and 10 mg/mL aprotinin). After centrifugation of the lysates at 13,000 g for 10 min, the supernatants were quantified for protein content by the Bradford method. Aliquots containing 60 μg of protein per lane were subjected to sodium dodecyl sulfate/10% PAGE under reducing (5% β-mercaptoethanol) conditions and electroblotted onto nitrocellulose membrane filters. The blots were blocked with 5% nonfat milk in 20 mM Tris/HCl, pH 7.5, 500 mM NaCl plus 0,1% Tween (TBS-T). The membranes were subsequently incubated in agitation at 4°C overnight in the 1% bovine serum albumin-TBS-T buffer containing, for each experiment, one of the specific antibodies against p53, caspase-8, -9, and -3. After four times washing with TBS-T, the blots were incubated for 1 h at room temperature with the second antibody conjugated to peroxidase, washed four times with TBS-T, developed with ECL detection reagents (Amersham) for 1 min, and exposed to X-Omat film (Eastman Kodak Co.).
Statistical analysis
The statistical analysis was performed by the Student's t test. The results are presented as the mean±standard deviation. Differences were considered to be significant at P<.05.
Results
Action of APE on HaCaT cell proliferation
To evaluate the toxicity of APE on the investigated system, HaCaT cells were incubated for 24 and 48 h with different concentrations of polyphenol extracts equivalent to 100 and 500 μM polyphenols.
The effects of APE on HaCaT cell viability determined by the trypan blue exclusion method is shown in Figure 1A. It is evident that the APE was able to exert a potent citotoxicity action on the HaCaT keratinocytes in a dose- and time-dependent manner both 250 and 500 μM (at concentrations below 250 μM, we did not observe any effect).
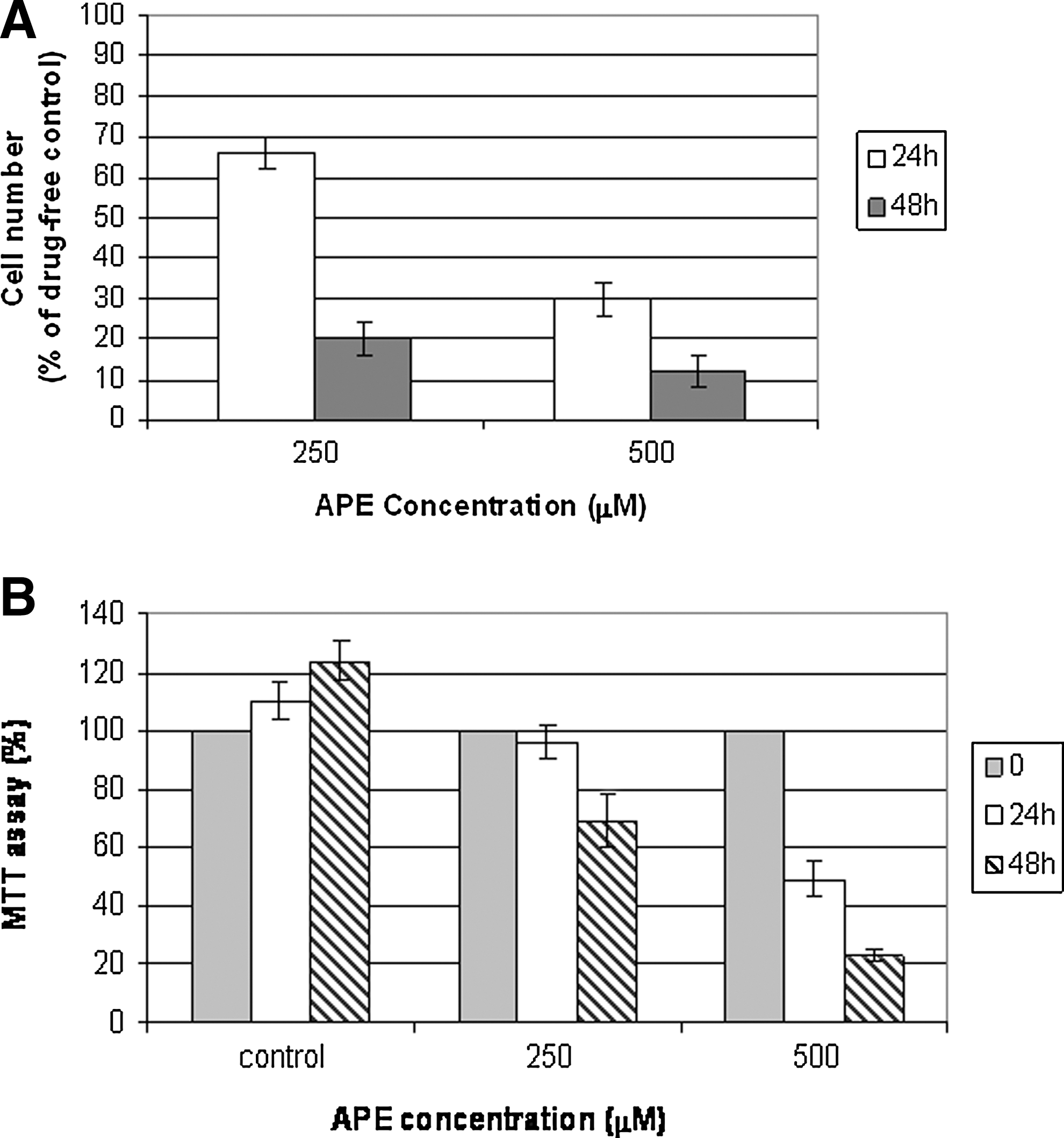
Effects of apple polyphenol extract (APE) on HaCaT cell growth.
The antiproliferative effect of apple polyphenols was subsequently tested by the MTT assay. 30
Figure 1B shows how the apple polyphenolic extract exerted a potent antiproliferative action on the HaCaT keratinocytes and also in this experimental test, the percentage of damaged cells varies not only as a function of APE concentration used, but also in relation to incubation time.
These results are in general agreement with our previous findings, regarding the growth inhibitory effect of different polyphenols on HaCaT keratinocytes. 12,30,31 Moreover, these data confirm the capacity of APEs to inhibit cell proliferation in vitro. 11
The percentage of damaged cells varies not only as a function of APE concentration used, but also in relation to incubation time.
Alteration of cellular morphology
Following 24 hours of incubation with different concentration of APE, morphology of HaCaT cells was observed under an inverted phase-contrast microscope. The number of treated HaCaT cells was obviously reduced, as compared with the control (vehicle containing 0.1% DMSO). Furthermore, morphological changes, including detachment from growing surface, nuclear fragmentation, and reduction of cell volume, could be observed in the treated cells (Fig. 2).
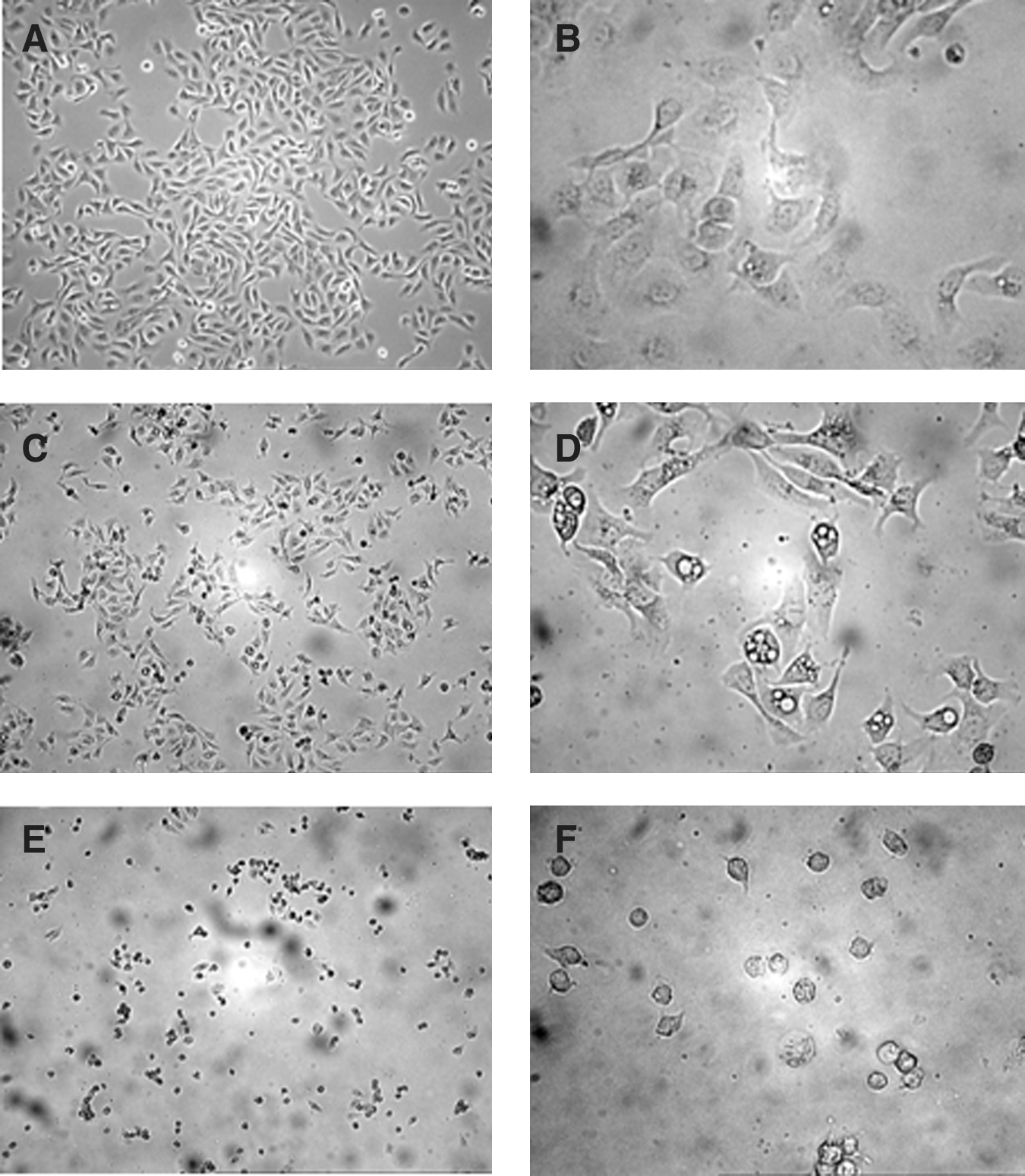
Action of APE on HaCaT morphology as examined by phase-contrast microscopy.
Evaluation of apoptosis by DNA-flow cytometry
HaCaT cells were exposed to various concentrations of APE (100, 200, 250, and 500 μM), harvested at different time points, and examined for DNA content. Apoptosis was investigated by DNA distribution as reveled by flow cytometry, demonstrating hypodiploid DNA as described in Materials and Methods. Figure 3 shows the results of experiments performed to evaluate the apoptotic response. When HaCaT cells were cultured alone, <2% was apoptotic, while the exposure of cells to apple polyphenol solutions caused a dose-dependent apoptosis. In fact, at concentrations of 100 μM, no effects on cell apoptosis were found, while 200 μM induced apoptotic death in about 19.8% of cells and 250 μM in about 51.5%, 24 h after incubation (Fig. 3). Finally, by using 500 μM, all the cells underwent detachment and death (data not shown). These results indicate that apple polyphenol induces massive keratinocytes apoptosis.
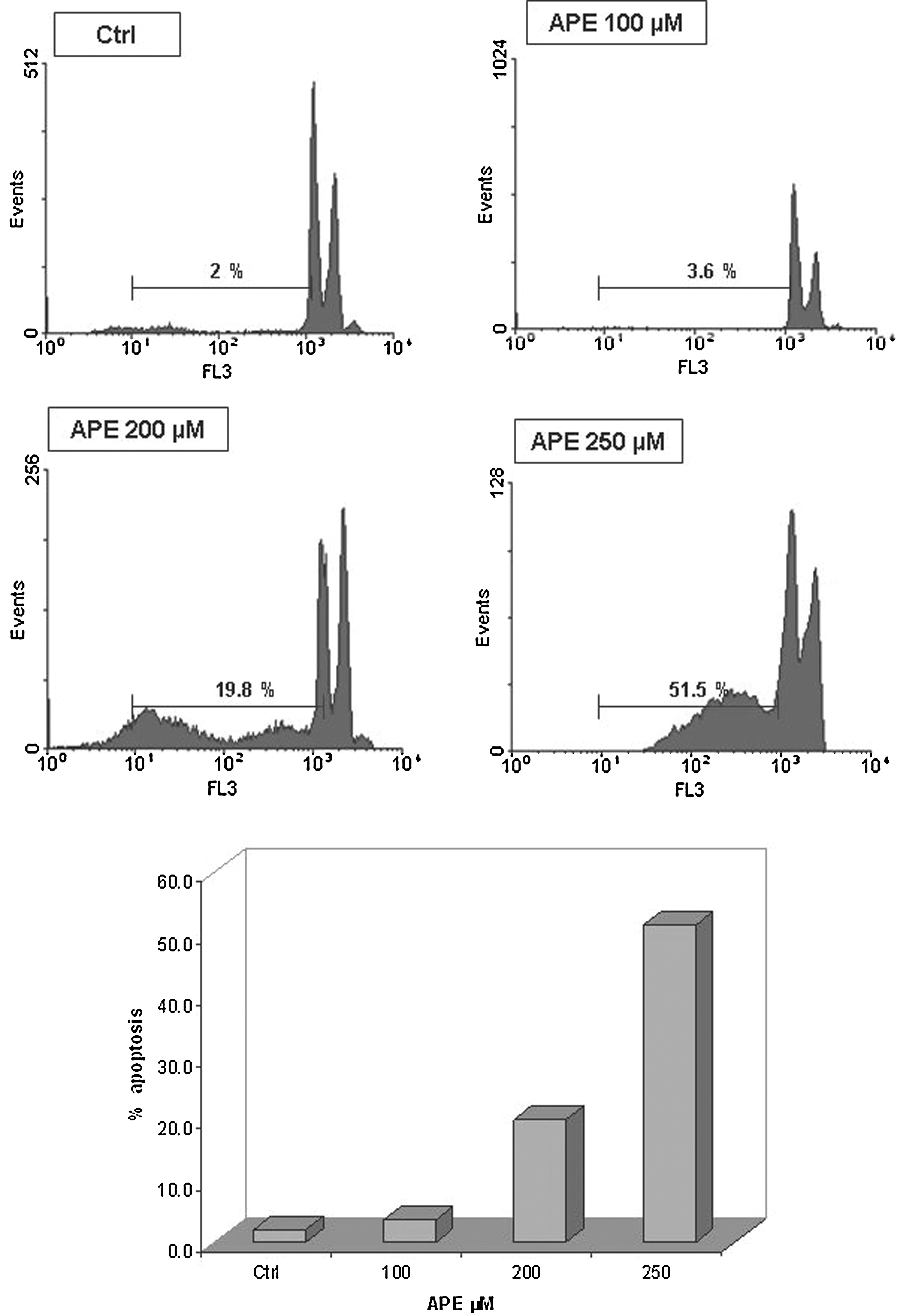
Effect of different APE concentrations on apoptosis of HaCaT cells. Flow cytometry analysis of DNA content of HaCaT cells exposed to various concentration of APE (100, 200, and 250 μM) for 24 h. Cells were collected and labeled with propidium iodide and analyzed as described in Materials and Methods. Apoptotic cells are characterized by low DNA stainability and appear below the G1 peak in the distribution, which is summarized in the bottom quantitative graph. Three experiments were performed with similar results.
Expression of p53 in APE-treated HaCaT cells
The tumor suppressor p53 protects mammalian cells from malignant transformation by preventing the propagation of genetic aberrations required for tumorigenicity. In cells that are exposed to such insults, p53 can induce either a cell-cycle arrest or apoptosis, depending in part on the severity of cellular damage incurred. 32 –34 Here, the role of p53 in apple polyphenols-mediated apoptosis of HaCaT cells was evaluated. Lysates derived from control and treated cells were subjected to Western blotting, using specific antibodies against p53. As shown in Figure 4, when cells were exposed to APE (100, 200, 250, and 500 μM), the p53 protein levels remained unaffected. These data indicate that APE causes HaCaT apoptosis through the p53-independent pathway.

Western blotting analysis of p53 expression. Cells were cultured for 24 hours with APE at the indicated doses and then lysed to prepare protein extracts. Electroblot on the nitrocellulose filter was incubated with p53-specific antibodies and detected by chemiluminescence. Staining of tubulin was used as an equal loading control. Data are representative of three independent experiments for each antibody.
APE-induced apoptosis is mediated by caspase activation
To investigate the apoptotic pathways activated by APE in HaCaT cells, the activity of the initiators caspase-8, caspase-9, and the effector caspase-3 was monitored.
Two major apoptotic pathways have been described for mammalian cells. The first one involves caspase-8, which is activated through membrane death receptor-mediated extrinsic pathways, whereas the second one implicates mitochondria-emanated intrinsic pathways, being mainly characterized by activation of caspase-9 via cytochrome c release into the cytosol and apoptosome formation. Subsequently, activated caspase-3, as the major effector component of both pathways, initiates a program of proteolytic cleavages of critical cellular proteins, including poly(ADP-ribose)polymerase (PARP) and Lamin A/C, eventually resulting to apoptosis. 35,36
The activity of these caspases was examined as a decrease in the proenzyme level and an increase in active fragments, using the Western blotting analysis.
The obtained results showed that exposure of HaCaT cells to APE for 24 hours led to the cleavage of caspase-8 and caspase-3 in a dose-dependent manner (Fig. 5), while no activation of caspase-9 was revealed by the immunoblot analysis (data not shown). These results suggest that the extract induced apoptosis through the extrinsic pathway.
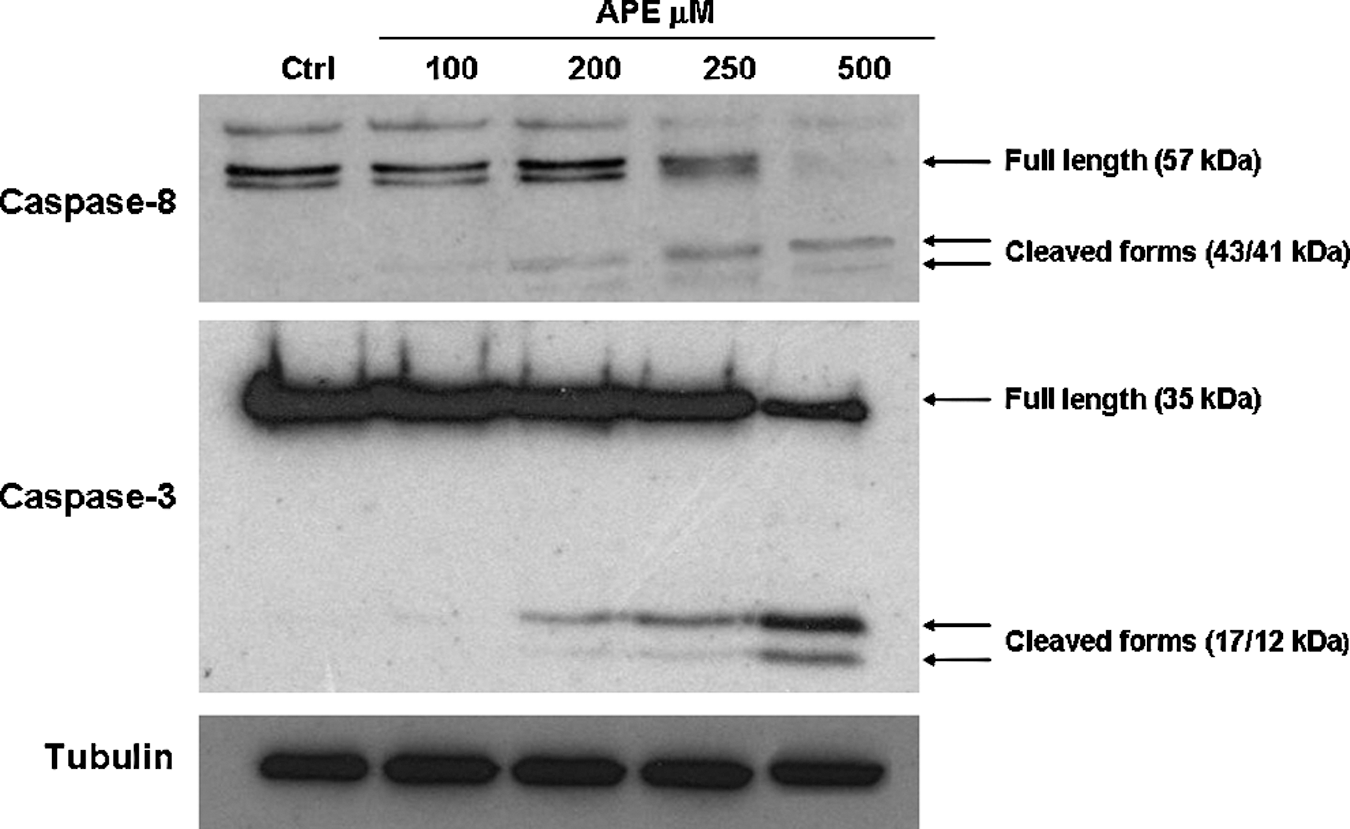
Western blotting analysis of caspase-8 and caspase-3 activation. Cells were cultured for 24 hours with APE at the indicated doses and then lysed to prepare protein extracts. Electroblot on the nitrocellulose filter was incubated with caspase-8, -3-specific antibodies and detected by chemiluminescence. Staining of tubulin was used as equal loading control. Data are representative of three independent experiments for each antibody.
Discussion
In the present study, we investigated the effects of Annurca apple polyphenols on proliferation and apoptosis in a human HaCaT keratinocytes, and analyzed the intracellular apoptotic signal transduction pathway linked to the activation of caspase. Recently, apple polyphenols have been found to induce apoptosis in cancer cells. In fact, Kern et al. (2007) have demonstrated that APE induced the caspase-3 activity and DNA fragmentation in HT29 cells. 37
Compounds that inhibit proliferation and modulate differentiation in keratinocytes are potentially useful in the treatment of psoriasis, because a balanced homeostatic control of keratinocyte growth and differentiation is crucial for recovery from psoriatic to normal epidermis. Our experimental results demonstrated that APE possesses potent inhibitory action on the growth of keratinocytes.
This favorable toxicity profile of the APE is important because it enables formulating topical applications of apple polyphenols that could exert significant therapeutic effect without evoking harmful side effects on normal skin cells.
The elucidation of the underlying cellular and biochemical mechanisms for the observed growth inhibitory action is necessary for the bioactive polyphenolic compounds to be developed as an effective therapy for psoriasis treatment. Because cellular apoptosis and/or necrosis could be responsible for growth inhibition of cultured cells, experiments were designed to elucidate, at morphological, molecular, and biochemical levels, whether induction of cellular apoptosis is responsible for the APE-mediated growth inhibition on human keratinocytes. It is well recognized that hyperproliferation of epidermal keratinocytes seen in psoriasis is the result of the aberrant expression of many regulatory molecules associated with proliferation, and defects in apoptosis are believed to play an important role in the pathogenesis of psoriasis. 38 Polyphenols that are able to inhibit keratinocyte proliferation and induce keratinocyte apoptosis would conceivably possess good potential for being developed into effective agents for treating psoriasis.
The results obtained here have indicated that apple polyphenols induced apoptosis in HaCaT as demonstrated by internucleosomic DNA fragmentation and quantitized by the fluorescence-activated cell-sorting (FACS) analysis after nuclear dye with PI. The phenomenon was dose-dependent and observed at concentrations as little as 200 μM; maximal effect was observed at 500 μM of extract.
Then, it seemed appropriate to study in detail the apoptotic molecular mechanism associated with the apple polyphenol action. We investigated whether the p53 protein was involved in the signaling pathway of HaCaT apoptosis, and we found the existence of a p53-independent signaling pathway to regulate the apoptotic response. In this regard, we hypothesized that alternative pathways are responsible for APE-induced cell death. It has been known that the extrinsic pathway involves engagement of particular death receptors that belong to the tumor necrosis factor receptors family and, through the formation of the death-inducing-signaling-complex, leads to a cascade of activation of caspases, including caspase-8 and caspase-3, which in turn induce apoptosis, while caspase-9 is the apical caspase in the intrinsic pathway. As displayed in the results, APE caused caspase-8 and caspase-3 activation, while caspase-9 was not activated, suggesting that the extrinsic pathway is dominant. Therefore, apple polyphenol induces apoptosis of HaCaT cells by triggering the death receptor-associated extrinsic pathway without recruitment of p53.
All our results unambiguously demonstrated that induction of cellular apoptosis is mainly responsible for the apple polyphenols mediated HaCaT keratinocyte growth inhibition, and this effect is dose- and time-dependent. The successful identification of APE as potent antiproliferative and apoptogenic agents not only places the traditional use of apple extracts for psoriasis on a scientific footing, but also renders them promising candidates for further development into topical therapeutic formulae for psoriasis treatment. In fact, in psoriasis an abnormal resistance of keratinocytes to apoptosis has been implicated as one of the key pathogenetic mechanism, leading together with the increased keratinocyte turnover rate to epidermal acanthosis. 39 –41 This suggests that the induction of keratinocyte apoptosis could be a common pathway of different therapeutic agents active in psoriasis via a direct effect on epidermal hyperplasia.
Collectively, our experimental results confirm that APE is capable of inducing programmed cell death in cultured HaCaT keratinocytes. The apoptotic actions observed in the present study provide an explanation for the underlying mechanism of the potent antiproliferative property exhibited by apple polyphenols on HaCaT cells.
Our experimental data suggest that apple polyphenols are a promising source from which a natural-based topical agent could be developed for skin diseases treatment.
Footnotes
Author Disclousure Statement
No competing financial interests exist.