
Abstract
C-phycocyanin (C-PC) from Spirulina has been previously shown to have anticancer properties. Here, we report on anticancer activity of C-PC that was isolated from the novel cyanobacterium Limnothrix sp. 37-2-1. C-PC from this organism exhibited anticancer properties in our in vitro systems; however, the required doses were well above the range of anticancer drugs normally used. Therefore, we conducted several experiments to test whether lower-than-usual doses of the anticancer drug topotecan (TPT) can offer the same level of cytotoxic effects as normal doses when combined with C-PC. For this purpose, cytotoxicities of C-PC and TPT were tested using the LNCaP (prostate cancer) cells. We found that when only 10% of a typical dose of TPT was combined with C-PC, the cancer cells were killed at a higher rate than when TPT was used alone at full dose. Similarly, we were also able to detect an increased level of radical oxygen species (ROS) generation as well as an increase in activities of caspase-9 and caspase-3 when these two compounds were used in combination. Taken together, our findings suggest that combining C-PC from Limnothrix sp. with the lower dose of TPT can induce apoptosis through generation of ROS and activation of caspases. In that respect, we suggest that C-PC can potentially improve the efficacy of the currently available anticancer drug, and therefore diminish its harsh side effects in the patient.
Introduction
C
C-PC is a blue-colored photosynthetic accessory pigment that absorbs light at 620 nm and emits fluorescence at about 560 nm. It is a protein containing two subunits, molecular weight of which depends on the organism from which it was isolated, and ranges between 50 and 230 kDa. 4 Subunit α carries one and subunit β has two tetrapyrol chromophores. 5
One of the first reports on beneficial effect of phycocyanin cites the Japanese patent #58-65216, according to which this blue pigment from Spirulina significantly increased the survival rates of mice that had been injected with liver tumor cells. 6,7 It has been suggested that stimulation of the immune system by phycocyanin was a mechanism that inhibited the growth of tumor cells. Liu et al. 8 also reported on antineoplastic activity of phycocyanin isolated from Spirulina. They found that this pigment significantly inhibited the growth of human leukemia K562 cells through mechanisms other than apoptosis. However, induced apoptosis was described as a mechanism of phycocyanin action by a number of authors. Reddy et al. 9 used mouse macrophage cells, which were treated with lipopolysaccharide (LPS) to express high levels of COX-2, overexpression of which is known to be responsible for downregulation of apoptosis. When these cells were exposed to C-PC, it induced apoptosis in a dose-dependent manner. A different mechanism of apoptosis induction was described in AK5 tumor cells. Pardhasaradhi et al. 10 explained the C-PC induction of apoptosis by downregulating of Bcl-2 (a known inhibitor of apoptosis) and by generation of radical oxygen species (ROS). This finding was also confirmed with the human chronic myeloid leukemia cell line K562. 11 At present, it is not known whether the whole C-PC molecule is required for the anticancer activity. Nevertheless, it was shown that the β-subunit of C-PC that was cloned in Escherichia coli had the same or even more potent power of apoptotic induction when tested on four different cancer cell lines. 12 Cells of the treated cancer cell lines underwent apoptosis with an increase in caspase-3 and caspase-8 activities with an arrest at the G0/G1 phase. Caspase-dependant apoptosis was also confirmed in C-PC–treated HeLa cells. 13
In this work, we used C-PC from our novel cyanobacterial isolate Limnothrix sp. 37-2-1 to test its anticancer properties. We have shown that Limnothrix can be a better source for C-PC than the traditionally-used Spirulina. 4 In this preliminary work, we have confirmed that C-PC from Limnothrix indeed had antiproliferative activity against a prostate cancer cell line (LNCaP). However, the required concentration of phycocyanin for anticancer activity was well above the range of anticancer drugs normally used. Therefore, we hypothesized that C-PC could be potentially used as an enhancer of existing anticancer drugs. Here, we are presenting data of in vitro experiments, which confirm our hypothesis.
Materials and Methods
Phycocyanin isolation and purification
Phycocyanin was isolated from the novel strain of cyanobacterium Limnothirix sp. 37-2-1 by a procedure described elsewhere. 4 Briefly, the cyanobacterium was grown in a 30-L photobioreactor; the biomass was harvested and extracted. The pigment was purified by fractional precipitation of the cell crude extract by ammonium sulfate and treated with activated carbon and chitosan. The pigment was desalted and concentrated by tangential flow filtration. The purity of C-PC was checked by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and by calculating the ratio of the absorbencies at 620 and 280 nm. Only those preparations of phycocyanin that had the 620/280 ratio greater than 4.0 were used for further experiments. The purified phycocyanin was freeze-dried and kept in a refrigerator until use.
Cell line and reagents
The prostate cancer (LNCaP) cell line was a generous gift from Dr. Thomas Powell (Cleveland Clinic Foundation, Cleveland, OH, USA). The RPMI-1640 growth medium, amphotericin-B,
Cell culture and treatment
The LNCaP cells were grown as a complete monolayer in RPMI-1640 growth medium supplemented with 10% fetal bovine serum, 10,000 U/mL of penicillin, 10,000 μg/mL of streptomycin, 1% (+)-
Determination of cell death and ROS
To determine the cytotoxic effect of C-PC and TPT, the monolayer culture of LNCaP cells was used. The cells were incubated in the presence of C-PC (250 and 500 μg/mL) and TPT (1 and 10 μM), individually and in combination. All incubations were carried out for 16 h. After the incubation period, cell death was determined by using trypan blue dye exclusion test. To determine the levels of ROS generation, the cells cultured in six-well plate were incubated with different concentrations of C-PC and TPT as described above. At the end of incubation period, levels of ROS generated were measured using the nitro blue tetrazolium (NBT) chloride reduction assay.
Assay of caspase-9 and caspase-3 activities
To determine the activity of caspase-9 and caspase-3, LNCaP cells were treated with different concentrations of the C-PC (250 μg/mL) and TPT (1 and 10 μM). After the incubation time, cells were harvested, washed, and resuspended in a cell lysis buffer and kept on ice for 10 min. The cell lysate was centrifuged at 1400 g for 5 min; the supernatant was assayed for protein concentration and further diluted with the lysis buffer to adjust the protein concentration. Equal amounts of protein from each sample were added to 96-well plates and mixed with the 2× reaction buffer (100 mM HEPES [pH 7.4], 200 mM NaCl, 20 mM dithiotheritol, 2 mM ethylenediaminetetraacetic acid, and 0.2% chaps) and 2 mM of the respective substrates acetyl-Leu-Glu-His-Asp-p-nitroaniline (Ac-LEHD-pNA) and acetyl-Asp-Glu-Val-Asp-p-nitroaniline (Ac-DEVD-pNA). For measuring the specific activity of caspase-9 and caspase-3, the respective inhibitors Ac-LEHD-CHO and Ac-DEVD-CHO were used. Release of the cleaved p-nitroanilide from the tetrapeptide substrates was measured using the 96-well microplate reader at 405 nm.
DNA fragmentation assay
DNA fragmentation assay was performed by using a monolayer of LNCaP cells that were grown in T-25 culture flasks and incubated with different concentrations of C-PC and TPT, individually and in combination. After incubation, the cells were harvested and washed with phosphate-buffered saline (PBS). Cells were resuspended in 200 μL of PBS, and 20 μL of proteinase K was added. The DNA was extracted using the Qiagen DNeasy Kit, following the manufacturer's protocol. The samples were subjected to electrophoresis at 80 V for 2 h in a 1.5% agarose gel containing 5 μL of ethidium bromide. Separated DNA fragments were viewed with an UVP image analyzer.
Statistical analysis
The data presented here represent mean±standard deviation values from at least four separate experiments. Statistical analyses were performed using one-way analysis of variance, followed by Student–Newman–Keuls multiple comparison tests. Values of P<.05 were considered as significant and presented in results.
Results
Treatment of LNCaP cells with C-PC and TPT resulted in a significant inhibition of cell growth (Fig. 1). By incubating the cells in the presence of C-PC at two applied concentrations (250 and 500 μg/mL), the survival rate of the cells was between 65% and 70%. However, this effect was more pronounced when C-PC was combined with TPT. When LNCaP cells were treated with 10 μM of TPT alone, 62% cells were killed in 18 h, whereas at the concentration of 1 μM, TPT caused death in only 3–4% cells within the same time frame. Therefore, to test our hypothesis that C-PC enhances the activity of the anticancer drug, we have chosen 1 μM concentration of TPT for combination treatments. When 1 μM of TPT was combined with 500 μg/mL of C-PC, 80% of cells were killed due to the additive effect. Similarly, the treatment of LNCaP cells with C-PC and TPT showed a significant increase in the rate of ROS generation (Fig. 2). The highest ROS production was obtained when the cells were incubated with a combination of C-PC (500 μg/L) and TPT (1 μM).

Viability of LNCaP cells after treatment with phycocyanin (C-PC) and topotecan (TPT), individually and in combination. Dimethyl sulfoxide was used as a control. Values are mean±SD of four individual experiments. SD, standard deviation.
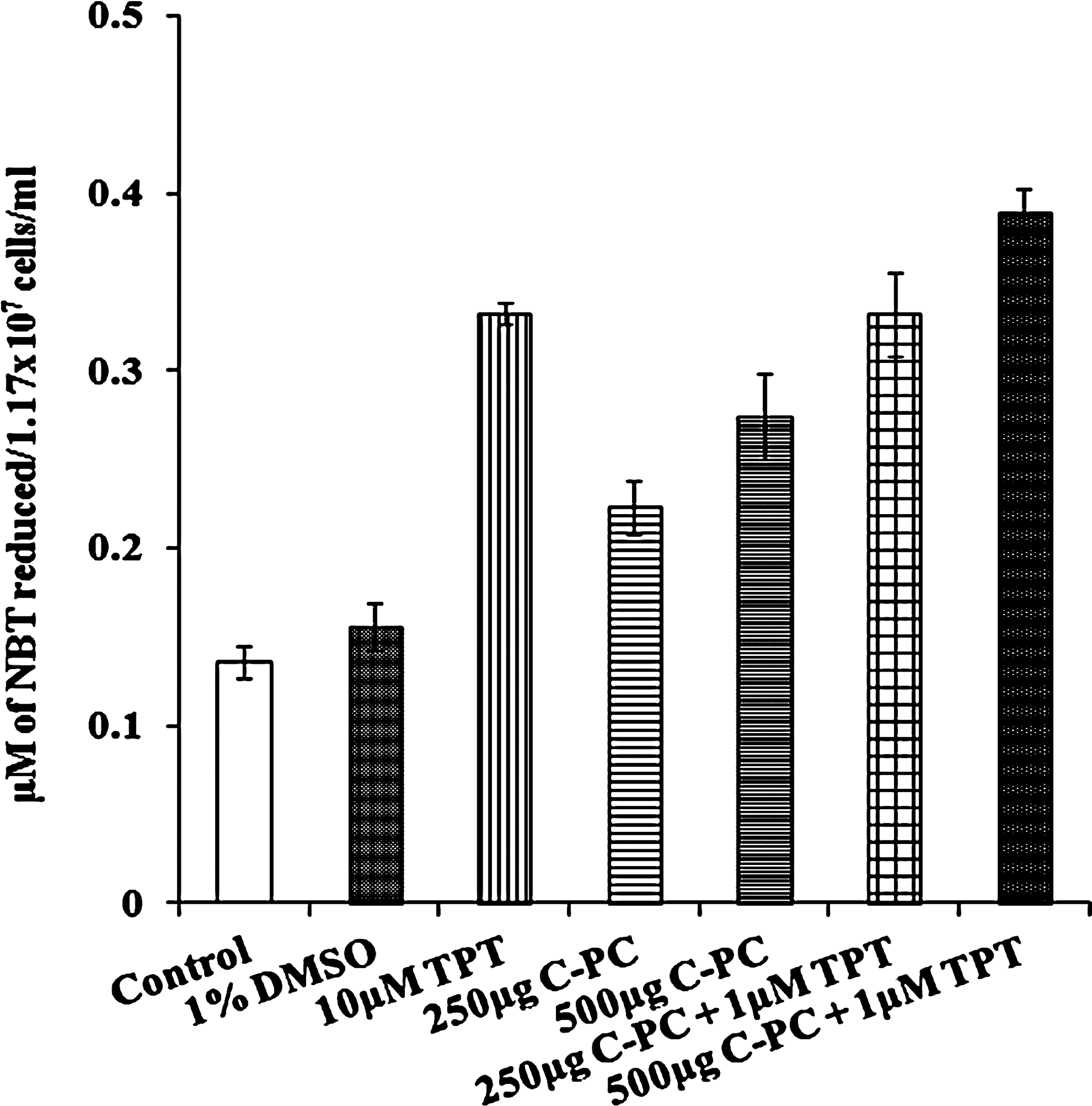
Levels of ROS in C-PC– and TPT-treated LNCaP cells. ROS were measured based on the nitro blue tetrazolium (NBT) chloride reduction assay. Values are mean±SD of four individual experiments. ROS, radical oxygen species.
The inhibition of cell viability could have resulted from the induction of apoptosis. To confirm this, the cells treated with C-PC and TPT were subjected to DNA fragmentation analysis. Figure 3 clearly shows internucleosomal DNA fragmentation of cells treated with different concentrations of C-PC and TPT. The DNA fragmentation was the most pronounced when the C-PC–TPT combined treatment was applied.

A representative photograph of agarose gel showing DNA fragmentation in LNCaP cells treated with C-PC and TPT. The DNA was separated by electrophoresis using a 1.5% agarose gel. DNA fragments stained with ethidium bromide were visualized using an UVP image analyzer. Lane 1, DNA ladder; lane 2, control; lane 3, TPT 1 μM; lane 4, TPT 10 μM; lane 5, C-PC 250 μg/mL; lane 6, C-PC 500 μg/mL; lane 7, C-PC 250 μg/mL+TPT 1 μM; lane 8, C-PC 500 μg/mL+TPT 1 μM.
To identify whether caspases were involved in C-PC–induced apoptosis, the C-PC– and TPT-treated cell lysates were analyzed for the caspase-3 and caspase-9 activities by using colorimetric assay. As expected, caspase-3 and caspase-9 (Fig. 4) activities in C-PC– and TPT-treated cells were found increased when compared to untreated control cells. Our data suggest that C-PC–induced apoptosis is characterized by the caspase-dependent pathway.
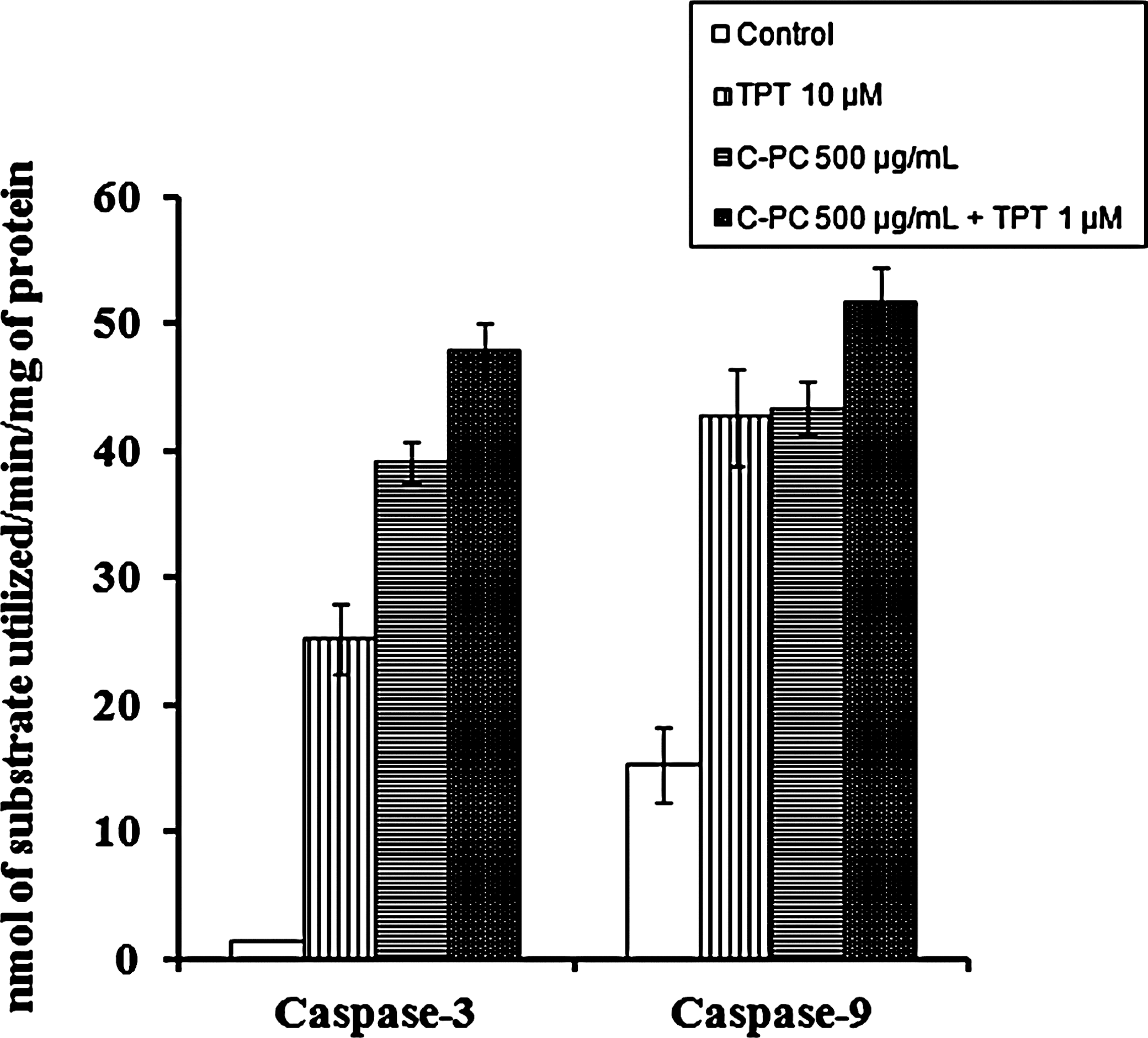
Activities of caspase-3 and caspase-9 in LNCaP cells treated with C-PC and TPT. Caspase-9 and caspase-3 activities were measured using the synthetic tetrapeptide substrates acetyl-Asp-Glu-Val-Asp-pNA and acetyl-Leu-Glu-His-Asp-pNA, respectively. Values are mean±SD of four individual experiments.
Discussion
The use of natural products to inhibit carcinogenesis and to treat cancer is an important and rapidly growing field of cancer research. There has recently been a surge of interest in diverse bioresources as potential novel sources of anticancer compounds. One of these includes cyanobacteria (blue-green algae), which were shown to produce a number of anticancer compounds such as scytophycins, 14 tolyporphin, 15 apratoxin A and antillatoxin B, 16,17 malevamide D, curacin D and symplostatin 3, 18 lyngbouilloside, 19 and cryptophycin. 20 However, in addition to these toxic compounds, cyanobacteria produce a blue pigment C-PC, which is a nontoxic compound, 21 yet it has anticancer properties. 11,22 This compound is readily available as a component of Spirulina biomass, which is sold in health food stores worldwide.
In this article, for the first time, we present our preliminary data on antineoplastic activity of C-PC that was isolated from the novel cyanobacterial strain Limnothrix sp. 37-2-1, and also we showed that when combined with the standard chemotherapeutic agent TPT, C-PC enhanced the cancer cell death. Specifically, when only 10% of typical dose of TPT was combined with 500 μg/L of phycocyanin (Fig. 1), the cancer cells were killed at a much higher rate than when TPT and phycocyanin were used separately.
Accumulative evidence suggests that defects in the process of apoptosis may be closely associated with carcinogenesis, and that many cancer cells have defective machinery for self-destruction. 23 Apoptosis is a specific mode of cell death recognized by a characteristic pattern of morphological, biochemical, and molecular changes. Most of the existing cancer drugs target apoptosis. Our finding that C-PC affects the LNCaP cells by inducing apoptosis is confirmation of previous reports in which C-PC used was obtained from Spirulina. 9,11 In our experiments, apoptosis was confirmed by DNA fragmentation (Fig. 3), shown both when C-PC and TPT were used individually and in combination. One of the probable causes of cell death during C-PC–anticancer drug combined treatment is induction of ROS production, leading to caspase-3 activation and DNA fragmentation. Our study confirms earlier report 10 that C-PC indeed induces ROS-dependent apoptosis. The highest level of ROS production was observed when the LNCaP cells were treated with the combination of TPT and C-PC. Furthermore, C-PC treatment increased the activities of both caspase-3 and caspase-9, which are the most important proteases found in apoptotic cells. 23 Further in vivo and in vitro molecular studies will be needed to identify the appropriate application conditions and doses of C-PC in the treatment process.
Combining the known anticancer drugs with natural products for the treatment of cancer patients is apparently a new avenue in cancer research. This kind of approach and its benefits have been reported recently by He et al., 24 who showed that resveratrol, a unique polyphenol produced by plants, potentiates the growth inhibitory effect of rapamycin. Whether used in a combined treatment or in the form of a hybrid molecule, 25 this approach deserves further research.
In conclusion, the results suggest that the use of the C-PC–TPT combination can reduce the effective dose of TPT, and still maintain its efficacy in inducing significant apoptotic cell death in LNCaP cells. Based on these preliminary data, it is suggested that the C-PC–TPT combination may have a therapeutic value.
Footnotes
Acknowledgment
The authors would like to thank the Royal Dames of Cancer Research Inc. (Ft. Lauderdale, FL, USA) for their generous financial support.
Author Disclosure Statement
No competing financial interests exist with this work.