
Abstract
Recent studies indicate that an oil extract from Salvia sclarea may provide clinical benefits in various pathological conditions. In comparison to extracts from other Salvia species, S. sclarea oil contains twice as much omega-3 fatty acids, which are involved in eicosanoid synthesis pathways, and has been found to contain significant levels of the psychoactive monoterpane linalool. In the present study, we examined the mood stabilizing and anxiolytic-like effects of chronic food administration of S. sclarea oil extract on behavioral and physiological parameters of mice with prominent dominant and submissive features in behavioral assays used to test mood stabilizing and antidepressant drugs. Experimental animals received oil supplemented food from the age of 4 weeks or from conception via their pregnant dams. Each age group received either S. sclarea oil– or sunflower oil–enriched feed. Dominant animals, whose pregnant mothers received S. sclarea oil–enriched feed from the date of conception, showed a significant reduction of dominant and anxiety-like behavior, in comparison to their sunflower oil–treated counterparts. S. sclarea oil–treated submissive animals exhibited a similar tendency, and showed a significant reduction in blood corticosterone levels. These findings enforce the hypothesis that S. sclarea oil possesses anxiolytic properties.
Introduction
S
The potential health advantages of omega-3 fatty acids are currently under vigorous study and are believed to stem from their anti-inflammatory effects achieved through modulation of eicosanoid synthesis pathways. 5 –8 Another factor mediating the potential effects of dietary supplementation with omega-3 is the ratio of omega-6 to omega-3 fatty acids consumed, since the two types of acids are in direct competition for desaturase and elongation enzymes. Nutritional guidelines recommend a dietary ratio not exceeding 1:4 in favor of omega-6, to encourage the biosynthesis of anti-inflammatory prostaglandins, thromboxanes, and leukotrienes. 6,8,9 The ratio of omega-3 to omega-6 fatty acids in S. sclarea oil extract is approximated to reach 3:1 in favor of omega-3, while typical western diets are estimated to contain fatty acids at a ratio of from 1:10 to as much as 1:20 in favor of omega-6. 2,8
Investigation of the therapeutic properties of omega-3 acids has revealed encouraging evidence regarding their use in the secondary prevention and supplementary treatment of cardiovascular disease, arthritis, renal disease, type 2 diabetes, inflammatory bowel disease, as well as certain cancers. 5,6 In addition, a number of studies may suggest a possible therapeutic effect of the omega-3 fatty acids DHA and EPA upon mood disorders. 10 A reduction in depressive symptoms was seen among patients suffering from clinical depression and bipolar disorder as a result of EPA supplementation alone 11,12 and as adjunct therapy to standard psychiatric medications. 13 –15 Also, DHA supplementation has been shown to increase membrane fluidity and to encourage signal transduction initiation required for the production and function of serotonin and dopamine in the brain. 6,7
As opposed to EPA and DHA, ALA has not been shown to be directly involved in the regulation of the inflammatory response, and its involvement in brain development is not clear. 6 –8 While analysis of the brain fatty acid composition of animals used to model depression found elevated levels of the proinflammatory arachidonic acid, 16,17 only limited research has been dedicated to the potential effects of ALA upon mood disorders, with a lack of conclusive results. 6,10,18 In the present study, we examined the effect of food administration of S. sclarea oil on behavioral and biochemical parameters of animals with prominent dominant and submissive features. These animals were developed using a selective breeding approach, based on the Dominant–Submissive Relationship (DSR) behavioral paradigm 19 –21 for antidepressant and mood-stabilizing drug testing. 21 As a comparison group, animal feed fortified with sunflower [Helianthus annuus L. (Asteraceae)] oil was used. The effect of prolonged oil supplementation was evaluated upon animals that received supplemented food (1) from the age of 4 weeks or (2) from conception via their pregnant dams. Each age group was further divided into two treatment groups, each of which received either S. sclarea oil– or sunflower oil–enriched feed.
Materials and Methods
Animals
The populations of dominant (Dom) and submissive (Sub) mice used in this study are descendants of the outbred Sabra line, which freely develop relationships of dominance and submissiveness in the DSR test, and whose behavioral and biochemical characteristics were described recently. 22 These animal populations were selectively bred for ten generations at the Ariel University on basis of their behavior in the DSR test 19 (described in the DSR testing section). Animals were given their specifically designed rodent chow (described in the Animal feeds section) and water ad libitum. During DSR testing, chow was provided according to the DSR protocol. The colony room was maintained on a 12-h light–12-h dark cycle (lights on 7 a.m.–7 p.m.). The experiments were conducted in compliance with the NIH/USDA guidelines, under the approved Institutional Animal Care and Use Committee.
Animal feeds
Both groups of animals were fed standard commercial rodent food (2018SC; Harlan Laboratories, Madison, WI, USA) enriched with either S. sclarea (Tekland TD10359; Harlan Laboratories) or sunflower (Tekland TD10360; Harlan Laboratories) oil to the level of 160 mg oil per kg of isocaloric chow. Both chows contained an identical amount of total fat (5.9% by weight). Based upon food consumption, supplemental oil levels were approximated at 225 mg/day. The S. sclarea oil additive was derived from S. sclarea, which is approximated to contain 38.6% ALA and 12% linalool. 3 The sunflower oil additive was found to contain 5.9% ALA. The ratio of omega-3 to omega-6 fatty acids in the S. sclarea oil is approximated to be 2:1 in favor of omega-3, while the sunflower oil contains fatty acids at a ratio of 1:10 in favor of omega-6.
As presented in Table 1, animals were exposed to oil-enriched chows according to two regimens, in the first of which (From Weaning), males were fed enriched chow from weaning at the age of four weeks, until the end of the experiment at the age of 3 months. In the second regimen (From Conception), females received the enriched chow from the first day of pregnancy, and their male offspring continued to receive oil enrichment until the end of the experiment. Animals from each age group (From Weaning and From Conception) received either S. sclarea– or sunflower-enriched feed (n=10 each for Dom and Sub).
DSR testing
The DSR test is a food competition paradigm identifying dominant and submissive behavior, developed as a screening test for antidepressants and mood-stabilizing agents described previously. 19,20,23 Animal pairs, whose drinking times have at least a 40% disparity between them, are considered to have developed a DSR. 19 In short, the apparatus, made from Plexiglas, consists of two identical chambers (12 cm×8.5 cm×7 cm) joined by a tunnel (2.5 cm×2.5 cm×27 cm) with a 0.5-cm-diameter hole in its bottom center. A self-refilling feeder is connected to the tunnel, allowing a constant supply of sweetened milk (3% fat, 10% sugar), to which only one animal has access at any given moment. The tunnel has narrow slits cut on both sides of the feeder for easy gate insertion and removal. In this way, the paired mice have an equal starting position at the beginning of each session.
DSR tests were carried out on five consecutive days. During each 16-h period preceding testing, the mice were deprived of food, but water was provided ad libitum. Pairs of submissive mice from different home cages were matched for similar weight and were tested against each other in the DSR test daily. Dominant animals were also paired according to the same protocol. During each 5-min DSR session, milk drinking times were recorded manually by a human observer.
Open Field
The Open Field test is used to estimate spontaneous locomotor (horizontal) and exploratory (vertical) activity. 19,22,24 The Open Field apparatus consists of a clear glass box (30 cm×40 cm) whose floor is divided by lines drawn into 20 equally sized squares. Each mouse was placed individually in the center of the apparatus. The horizontal (number of squares crossed) and vertical (number of rearings) activity was recorded manually for 6 min. Between subjects, the apparatus was cleaned with 70% ethanol.
Elevated Plus Maze
The Elevated Plus Maze (EPM) test is used to assess anxiety-like behavior. 19,22,25 The apparatus is comprised of two enclosed (10 cm×45 cm×40 cm) and two open (10 cm×45 cm) arms that extend from a common central platform (10 cm×10 cm). The black apparatus is elevated to a height of 60 cm above the floor level. The tests were conducted during the period of illumination in the mice's colony room. For habituation purposes, all animals were placed in the experimental room 1 h before the test. Each animal was placed at the center of the maze, facing one of the enclosed arms. Each mouse spent a 5-min session in the EPM, during which the number of entries into open and closed arms, as well as time spent in the open and closed arms were manually scored. Between subjects, the apparatus was cleaned with 70% ethanol.
Blood corticosterone detection
Immediately following decapitation, blood samples were collected and centrifuged at 3500 g for 7 min. Serum was removed to a new tube and stored at −20°C for future use. Corticosterone in serum was determined using a Corticosterone ELISA kit according to the manufacturer's instructions (Assaypro EC3001-1).
Data analysis
Results are expressed as the mean±SEM, and statistical significance between S. sclarea– and sunflower-treated mice was assessed using the unpaired t-test or the two-way analysis of variance with post hoc Bonferroni t-test analysis, as appropriate (GraphPad Prism version 5.02).
Results
Effect of S. sclarea oil administration upon animals' behavior in the DSR test
At the age of 16 weeks, dominant and submissive mice treated with S. sclarea oil were tested against sunflower oil–treated animals of their same behavioral phenotype in the DSR test. Among animals who received oil supplementation from age 4 weeks, no differences were found between the drinking times of S. sclarea and sunflower groups of submissive or dominant animals (data not shown). Among animals who received oil supplementation from conception (whose dams received oil supplementation from the first day of pregnancy), S. sclarea oil–treated dominant animals drank significantly (P<.05) less milk than their sunflower oil–treated dominant counterparts in the fourth and fifth days of DSR testing (Fig. 1B). Submissive animals treated with S. sclarea oil from conception also showed a nonsignificant reduction in drinking time in comparison to sunflower oil–treated submissive counterparts (Fig. 1A).

Behavior of animals receiving food oil fortification from conception in the Dominant–Submissive Relationship (DSR) test. DSR testing showed no effects of oil fortification upon submissive mice
Effect of S. sclarea oil administration upon animals' behavior in the open field test
After completion of the DSR testing, the effect of chronic S. sclarea oil administration upon animals' spontaneous locomotor and exploratory activity was measured in the Open Field test. S. sclarea oil administration did not alter the activity of mice treated with S. sclarea oil from weaning in comparison to their sunflower oil–fed counterparts (data not shown). Animals fed oil-fortified food from conception presented a similar picture in locomotion (Fig. 2A), while a nonsignificant tendency was seen among S. sclarea oil–treated dominant animals to exhibit more rearing (Fig. 2B).
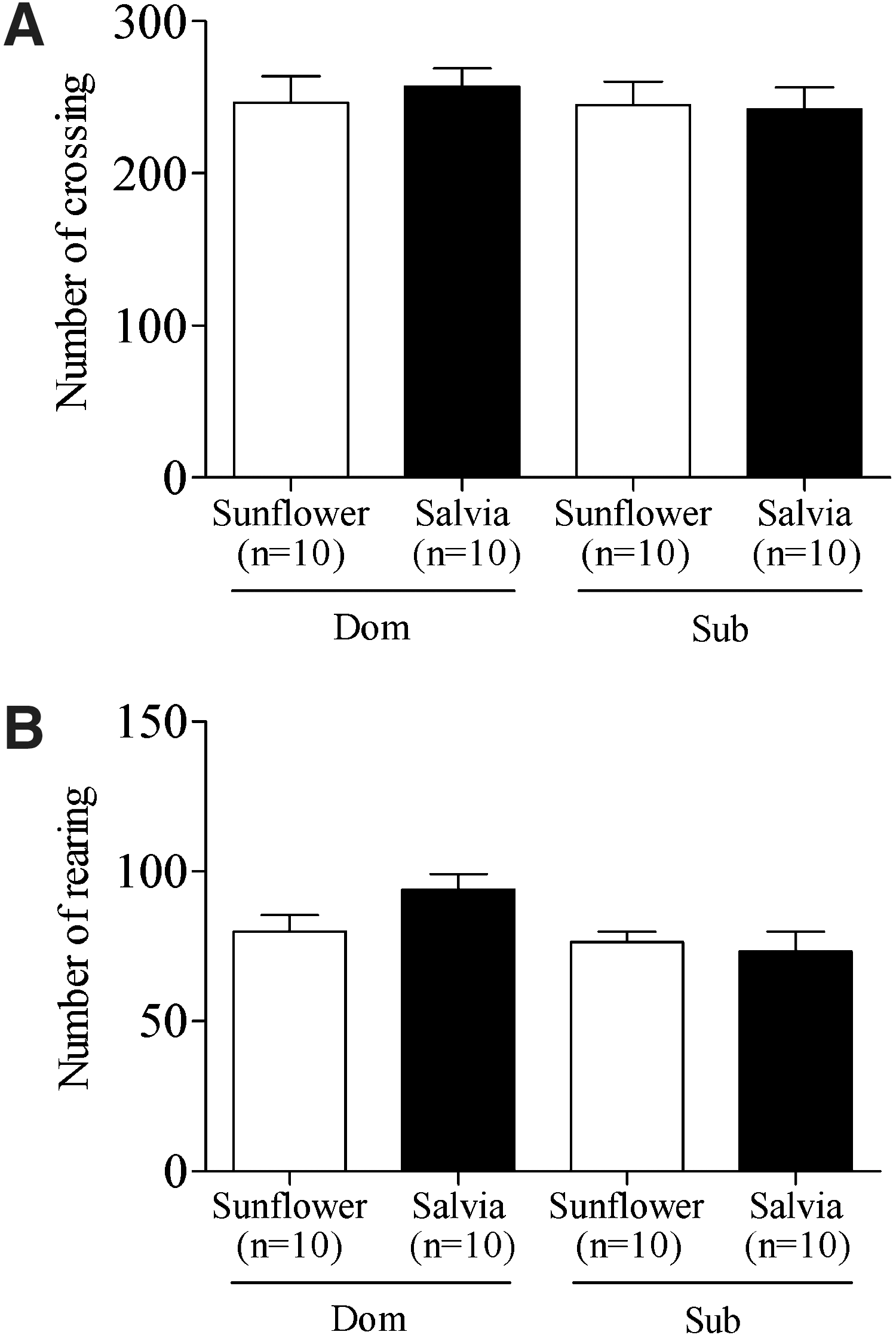
Food oil fortification did not alter animals' behavior in the Open Field test. The Open Field testing of animals fed oil-enriched food from conception revealed no discrepancies among spontaneous ambulatory
Effect of S. sclarea oil administration upon animals' behavior in the EPM
Following the Open Field test, animals' anxiety-like behavior was tested in the EPM. The number of entries to the open and closed arms, and the time spent there, were recorded. Animals receiving oil fortified food from weaning showed no differences between the S. sclarea oil– and sunflower oil–treated animals' activity in the EPM test (data not shown). Among animals fed fortified food from conception, the Total Entries and the Open Time/Total Time indices showed no differences between the S. sclarea and sunflower oil treatment (Fig. 3A, B), while the Open Entries/Total Entries index of dominant animals that received S. sclarea oil–fortified feed was significantly (P<.01) higher (Fig. 3C).

Anxiety-like behavior of animals receiving S. sclarea oil fortification from conception in the elevated plus maze (EPM). EPM testing of animals given oil-enriched food from conception found no difference in anxiety-like behavior measured by the Total Entries
Effect of S. sclarea oil administration upon animals' serum corticosterone levels
After completion of the behavioral studies, the animals were culled and serum samples taken for corticosterone level analysis. Animals receiving oil-fortified food from weaning showed no differences in corticosterone levels based upon S. sclarea or sunflower oil treatment (data not shown). Among animals fed fortified food from conception, the unpaired t-test found that submissive animals fed S. sclarea oil–fortified feed had significantly (P<.05) lower corticosterone levels than their sunflower-fed submissive counterparts (Fig. 4). No differences were observed in the corticosterone levels of dominant animals (Fig. 4).
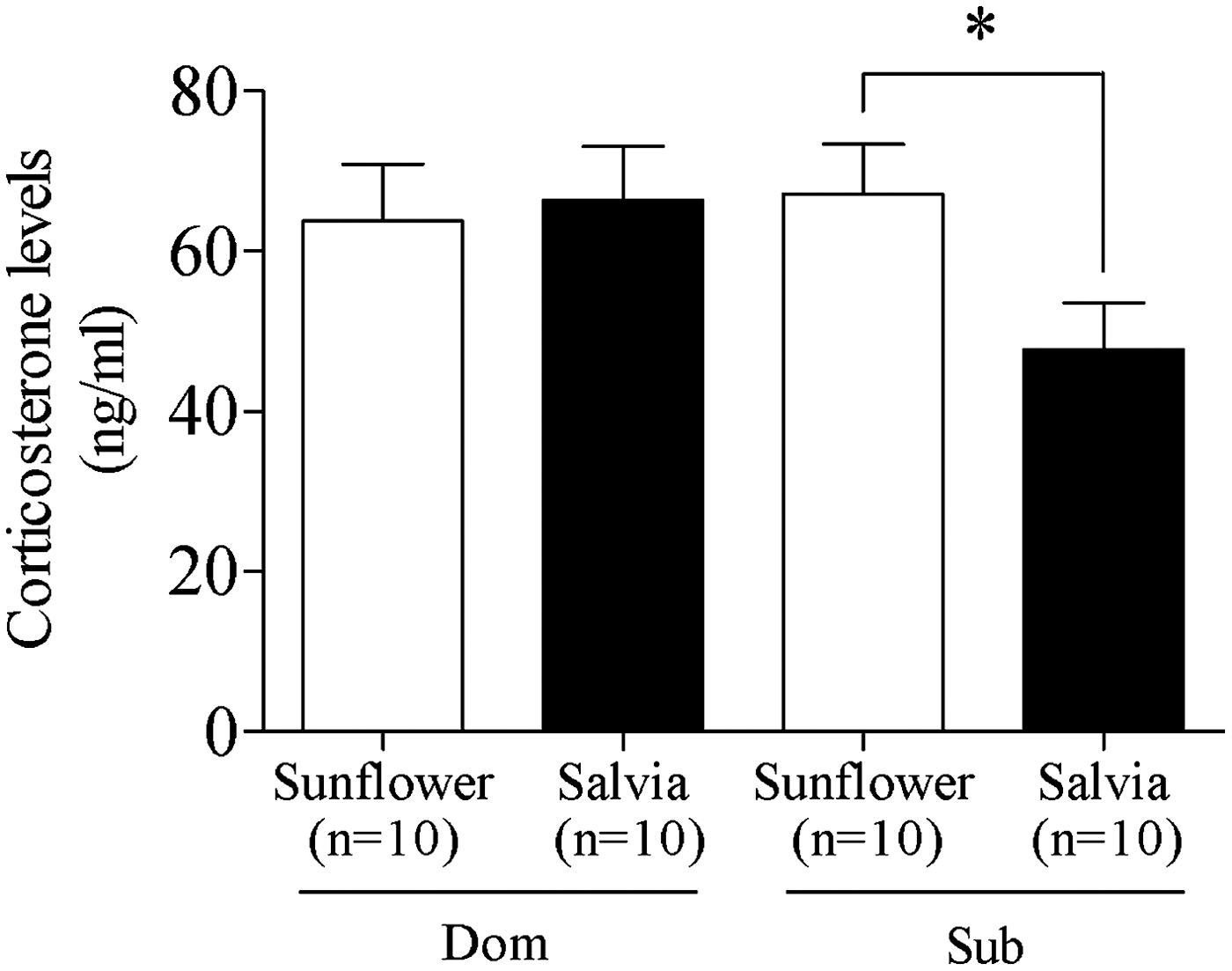
Corticosterone levels of animals receiving food oil fortification from conception. Submissive animals who received S. sclarea oil had significantly (P<.05) lower serum corticosterone levels than their counterparts receiving sunflower oil–enriched feed. Significances were shown using the unpaired t-test (*P<.05).
Discussion
This work's major finding is that chronic S. sclarea oil supplementation significantly reduced animals' dominant behavior in the DSR test and their levels of anxiety in the EPM test. These effects were observed only among animals whose pregnant mothers began treatment on the first day of conception, as seen in other studies that found beneficial effects on postnatal mental development from prenatal maternal dietary supplementation with omega-3 acids. 6,26 The present study is the first to describe anxiolytic-like effects from S. sclarea oil ingestion in an animal model of affective disorders.
Also noteworthy is the finding that submissive animals treated from conception with S. sclarea oil had significantly (P<.05) lower corticosterone levels than their sunflower-treated counterparts (Fig. 4). Submissive animals in the DSR paradigm acquire a consistently subservient social status in relation to their dominant counterparts, reminiscent of social defeat paradigms designed to induce the stress response. 27 Social stress is a factor with well-established effects upon the hypothalmus–pituitary–adrenal (HPA) axis, in which stress induces the release of the corticotropin-releasing factor (CRF) and vasopressin from the hypothalamus. CRF then activates the synthesis of proopiomelanocortin in the anterior pituitary, whose post-translational cleavage yields the adrenocorticotropic hormone, which stimulates the secretion of glucocorticoid hormones such as cortisol in humans and corticosterone in rodents from the adrenal cortex. 28 Corticosteroids reach every organ through the circulatory system, thus enabling coordination between brain and body functions aimed at management of stress, recovery, and adaptation. 28 However, exposure to chronic stress may disrupt the normal, adaptive stress response. 29 In individuals with perturbed HPA axis signaling, chronic stress exposure may act as a trigger for the development of behavioral disorders, such as depression and anxiety. 30 Submissive animals have recently shown an exaggerated corticosterone response to stress. 31 In the present study, S. sclarea oil significantly attenuated submissive animals' hormonal response to social interaction-related stress.
One of the challenges encountered in the course of experimentation based on dietary intervention is the introduction of potential confounding factors, such as differences in caloric intake or the balance of macronutrients. To minimize such confounding factors, in the present study, control animals received feed fortified with sunflower oil at concentrations isocaloric to the S. sclarea oil–enriched feed. The sunflower oil–fortified feed contained certain fatty acids, most prominently omega-6 linoleic acid (LA 18:2), in amounts much higher than in S. sclarea oil. Thus, one could claim that the sunflower oil strengthened the dominant phenotype of the mice who received this food. However, sunflower oil has not been found to possess psychoactive properties. 10 Moreover, the fatty acid composition of sunflower oil–supplemented chow was almost identical to that of regular laboratory chow in standard use. Therefore, this qualification does not appear to detract from S. sclarea's moderating effect upon dominant behavior in the DSR paradigm.
Indeed, in the DSR testing conducted between dominant animals, which underwent dietary intervention from the day of conception, S. sclarea oil significantly reduced the drinking time of dominant mice. Previous studies have validated the use of the DSR paradigm for the detection of behavior representing mood disorders, in which submissiveness represents depressive-like behavior and dominance represents manic-like behavior, 19 –21,32 and clinical antidepressants and mood-stabilizing drugs reduce submissive and dominant behavior, respectively. 19,23 The reduction of dominant behavior found among S. sclarea oil–treated dominant mice therefore may be explained as a mood-stabilizing effect of S. sclarea oil treatment. The lack of effects upon submissive mice in the DSR test may point to a selective mood-stabilizing action of S. sclarea oil treatment upon dominant animals.
The mood-stabilizing effect of S. sclarea oil upon dominant animals in the DSR paradigm was correlated with its anxiolytic effects in the EPM test. The Open Entries/Total Entries ratio of these dominant animals who received S. sclarea oil was significantly (P<.01) higher than of those who received sunflower oil (Fig. 3C). This seeming anxiolytic effect of S. sclarea oil was also reflected in a nonsignificant tendency among dominant animals to exhibit more rearing activity in the Open Field test. A broad range of antidepressant compounds have been shown to increase spontaneous locomotion in the Open Field test, of which the vertical movement is considered to be more sensitive than the horizontal movement to pharmacological treatment. 24 Thus, this tendency seen among S. sclarea oil–treated dominant mice may be a nascent expression of the antidepressant-like effects of S. sclarea oil upon these animals. At the same time, the lack of difference in the horizontal activity between treatment groups supports the claim that the increased entries into the open arms of the EPM does not result from a generalized increase in activity.
S. sclarea oil may exert its mood stabilizing and anxiolytic-like effects by the modulation of inflammatory signaling pathways attributed to omega-3 fatty acids. 6,7 A growing body of evidence supports the role of chronic inflammation in the etiology of depression, both in clinical research 33 and in animal models, 34,35 suggesting that inflammatory mechanisms may offer new targets for intervention in the treatment of affective disorders. 36,37 Specifically, a recent study found that dietary administration of a plant-derived omega-3-fatty acid extract with very high levels of ALA (60% of the extract) protected the mitochondrial function in the brain, although the behavioral parameters of the experimental animals were not measured. 38 Additionally, future studies of dietary S. sclarea oil administration should measure the bioavailability of linalool, which was found to strengthen adaptive aspects of the physiological stress reaction in animals when inhaled. 39 –41
In conclusion, we would like to stress that the anxiolytic effect of S. sclarea oil was seen among animals whose dams were treated with enriched feed from the first day of conception. We may speculate that the anxiolytic effect of S. sclarea oil upon pregnant dams contributed to their offspring's behavioral phenotype. There are many studies showing that prenatal stress exposure induced HPA axis dysregulation, leading to anxiety-like behavior upon offspring, in both clinical 42 –44 and animal 45 –47 research. Such effects are believed to be mediated by proinflammatory cytokines and/or corticosteroids, both of which have been identified as potential targets of omega-3 fatty acids' biological activity. 5,6 Among humans, the effects of maternal stress upon children's susceptibility to behavioral disorders is subject to great interest, and the mechanisms potentially linking pregnant animals' dietary enrichment with S. sclarea oil and behavioral characteristics of offspring merit further study.
Footnotes
Acknowledgments
This work was supported by the Israeli Ministry of Immigrant Absorption. Food supplements were provided by Magnetika Ltd. Neither of the aforementioned had a role in the study design, collection, analysis and interpretation of data, or in the writing of the report.
Author Disclosure Statement
No competing financial interests exist.