
Abstract
Selectively inducing apoptosis in cancer cells is a much desired strategy when tolerance toward side effects is minimal during chemotherapy. In our search for natural products that can induce apoptosis in human cervical cancer cells (HeLa), we selected resveratrol and genistein for our study. We conducted several experiments to test whether genistein can synergistically enhance the apoptotic potential of resveratrol at doses lower than the usual cytotoxic dose. Both resveratrol and genistein were able to induce apoptosis by enhancing the activities of caspase-9 and caspase-3 by themselves and also in combination. After 24 h of exposure to resveratrol and genistein, individually or in combination, lowered mitochondrial membrane potential was observed in HeLa cells. In addition, the mitochondrial membrane potential in HeLa cells was decreased, forcing JC-1 to stay in the monomeric form. The monomeric JC-1(5,5′,6,6′ -tetrachloro-1,1′,3,3′-tetraethyl benzimedazolyl carbocyanine iodide) emitted green fluorescence. In the control group, the color of the fluorescence was red due to aggregation of JC-1 in the physiological pH. The treatment groups exhibited DNA fragmentation as the hallmark of apoptotic nuclear features. We also detected an obvious decrease in the level of HDM2 gene expression after both individual and combination treatments with resveratrol and genistein. Our findings suggest that resveratrol and genistein when combined can induce apoptosis at doses lower than usual doses, through the activation of caspases cascade, and by decreasing the expression of HDM2.
Introduction
C
In relation to cell proliferation, the HDM2 gene is found to be over-expressed in more than forty different types of malignancies, including sarcomas and leukemias, and thus qualifying itself as a classical oncogene. Although some conflicting data exist with regard to the effects of HDM2 expression on individual cancers, overall evidence convincingly explains the fact that an increased level of HDM2 expression is often related to a poor clinical prognosis. 15,16 HDM2 functions by binding and targeting p53 for proteasomal degradation; therefore, growth arrest and apoptosis caused by p53 tumor suppressor protein is minimized in these cells, shifting the equilibrium toward cell proliferation. 17 Hence, inhibiting HDM2 by using specific inhibitors or lowering the levels of HDM2 protein is expected to release p53 and increase apoptosis in HDM2-positive cancer cells.
Although many studies on the individual effects of anti-cancer mechanisms of polyphenols and isoflavone have been reported in the past, the combination effects of resveratrol and genistein toward altering HDM2 expression or enhancing the caspase-mediated apoptotic pathway have not been studied. Therefore, in this study, our aim was focused on testing the combination effects of resveratrol and genistein on HeLa cells to determine their therapeutic potential at reduced doses.
Materials and Methods
Cell line and cell culture
The HeLa cell line was purchased from American Type Culture Collection (Manassas, VA, USA). Antibiotic-antimycotic solution containing penicillin, streptomycin, and Amphoteracin-B was obtained from Mediatech Inc (Herndon, VA, USA). Fetal bovine serum was obtained from Hyclone (Logan, UT, USA). Resveratrol was obtained from MP Biomedicals, LLC (Eschwege, Germany). Genistein was purchased from Sigma-Aldrich (St. Louis, MO, USA). Other chemicals used are of reagent quality and were obtained through standard suppliers. The HeLa cells were grown as a monolayer in DMEM medium supplemented with 10% fetal bovine serum, (+)L-glutamine, 10,000 U/mL penicillin, and 10,000 μg/mL streptomycin. The cells were grown at 37°C under a humidified 5% CO2 atmosphere. Densitometry analysis was performed using Image J1. (42J) program developed by the National Institute of Health (Bethesda, MD, USA). All experiments were conducted using cells in the logarithmic growth phase.
Cell treatments
A total of five treatment groups were utilized in our experiments, including a control. The control group received only 0.5% dimethyl sulphoxide. Groups 2 and 3 were treated with resveratrol (25 μM) and genistein (5 μM), respectively. Group 4 received resveratrol (5 μM)+genistein (5 μM). Group 5 was treated with 10 μM of topotecan.
MTT assay-toxicity studies
Cell viability was measured by the MTT assay (Sigma-Aldrich), a colorimetric assay for estimating mammalian cell survival, proliferation, and activation based on the ability of viable cells to reduce yellow 3-(4, 5-dimethythiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) by mitochondrial succinate dehydrogenase. HeLa cells were plated at a density of 5×104 cells/well in 96-well plates, and incubated at 37°C under 5% CO2 for 24 h. Then, the cells were exposed to different concentrations of the test compounds (resveratrol and genistein) for 24, 48, and 72 h. After the treatment, the old medium was removed, and the cells were washed twice with fresh DMEM medium without phenol red; then, 25 μL of MTT (0.5 mg/mL in DMEM medium without phenol red) was added to each well and incubated at 37°C for 3 h. After incubation, the media were removed, and the formazan crystals were solubilized in 100 μL of acidic isopropanol (0.04 M HCl in absolute isopropanol). The converted dye was completely dissolved by pipetting up and down several times, and the dissolved dye was transferred into a 1.5 mL eppendorf tube and centrifuged at 17,900 g for 2 min. The absorbance of the supernatant was measured at 540 nm using a microplate reader.
Caspase-3 and caspase-9 activities assay
The synthetic tetrapeptides Ac-DEVD-and Ac-LEHD-pNA were used to detect caspase-3 and caspase-9 activities in HeLa cells. The release of p-nitroanilide from the peptide substrate in an in vitro spectrophotometric assay allowed for the quantification of the amount of total DEVD or LEHD cleaved by caspase activities. Cell-free extracts prepared 24 h after drug treatment (concentration mentioned in the cell treatments section) were incubated in 100 mM HEPES (pH 7.5), 20% glycerol, 0.1 mM EDTA, 1 mM DTT, and 2 mM ac-DEVD-pNA colorimetric substrate (in 10% dimethyl sulfoxide [DMSO]) for 1 h at 37°C. For determining the activity of caspase-3 and caspase 9, specific inhibitors Ac-DEVD-CHO or Ac-LEHD-CHO were used (100 μM in DMSO) for 30 min at 37°C before addition of the colorimetric substrate. The production of cleaved p-nitroanilide from the tetrapeptide substrate ac-DEVD-pNA or ac-LEHD-pNA was read at 405 nm using a Molecular Dynamics SpectraMax 250 microplate reader. Assays were performed in triplicate, and the results are presented as the average±SD.
Analysis of mitochondrial membrane potential assay (JC-1 staining)
The changes in mitochondrial membrane potential (Δψm) were measured using the 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl benzimedazolyl carbocyanine iodide (JC-1; stands for first J-aggregate-forming cationic dye) probe. Cells were seeded in 24-well plates for 12 h, then treated with the test compounds for 24 h, and stained with the JC-1 reagent from Cayman chemicals (San Jose, CA, USA) according to the manufacturer's instructions. After incubation for 15 min with the JC-1 staining reagent, JC-1 aggregates in the mitochondria (only in live cells) were detected as a red color by a fluorescence microscope using excitation/emission=540/570 nm. JC-1 monomers in the cytoplasm were detected as a green color at excitation/emission=485/535 nm.
DNA fragmentation assay
A monolayer of HeLa cells were grown in T-25 culture flasks and incubated with the test compounds for 48 h. After incubation, the cells were harvested and washed with phosphate-buffered saline (PBS). Cells were resuspended in 200 μL of PBS, and 20 μL of proteinase K was added. The DNA was extracted using the Qiagen DNeasy Kit by following the manufacturer's protocol. The samples were subjected to electrophoresis at 80 V for 2 h in 1.5% agarose gel containing 5 μL of ethidium bromide. Separated DNA fragments were viewed with a UVP image analyzer.
Reverse transcriptase-polymerase chain reaction of HDM2 mRNA
Total RNA from HeLa cells were isolated with Qiagen RNA isolation kit according to the supplier's protocol (Madison, WI, USA). The total RNA extracted from the cells (after 24 h treatment of drugs) was dissolved in nuclease-free water and quantified using a Bio-Rad Spectrophotometer. Equal amounts (2 μg) of total RNA samples, including a control, were reverse transcribed and amplified by RT-PCR reaction for 45 min at 48°C with 1.0 unit of AMV reverse transcriptase obtained from Promega (Madison, WI, USA). The RT-PCR reaction was carried out in 35 cycles under the following conditions: denaturation at 94°C for 30 sec, annealing at 53°C for 1.0 min, and elongation at 68°C for 2.0 min. Starting from cycle 21, a 15 sec auto extension was added to the elongation steps until cycle 35. The number of reaction cycles utilized was established by performing several RT-PCR reactions with equal amounts of RNA with a different number of reaction cycles. The reactions were concluded with a final extension at 68°C for 7.0 min. Amplification of HDM2 cDNA was achieved using HDM2 gene-specific primers of the following sequence: sense primer 5′-CTG GGG AGT CTT GAG GGA CC-3′; antisense primer 5′-CAG GTT GTC TAA ATT CCT AG-3′. 18 The β-actin sense and antisense primer were 5′-GTG GGG CGC CCC AGG CAC CA-3′ and CTC CTT AAT GTC ACG CAC GAT TTC-3′, respectively. 19 The RT-PCR products were electrophoresed on a 1.5% agarose gel and visualized by staining with ethidium bromide. Densitometry analysis was performed using Image J1 (42J) program developed by the National Institute of Health. The relative levels of RNA were calculated by utilizing the band intensity values.
Statistical analysis
The data were presented as mean±SD of four or more determinations. Statistical analyses were performed using a one-way analysis of variance (ANOVA) followed by Student–Newman–Keuls Multiple Comparisons tests. P values<0.05 were considered significant.
Results
MTT assay
As shown in Fig. 1, low concentrations (5 μM) of resveratrol and genistein exerted moderate cytotoxic effects on HeLa cells and showed clear time-response relationships at the specified concentration. This result suggested that resveratrol (5 μM) was directly causing toxicity to HeLa cells that got potentiated when they were combined with genistein (5 μM) during the 24, 48, and 72 h of treatment.
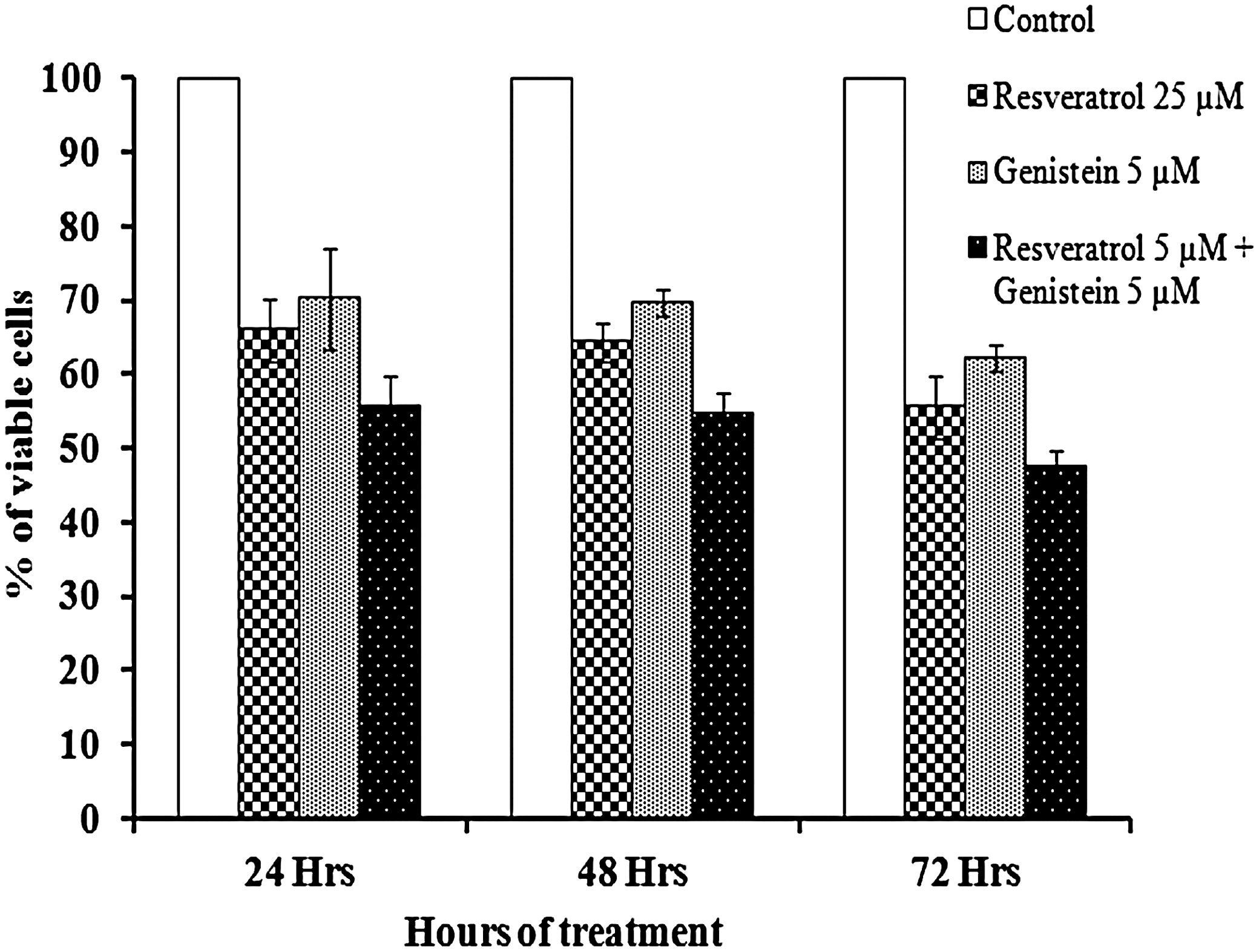
Time-dependent effect of resveratrol, genistein, and combination treatment on human cervical cancer (HeLa) cell proliferation. The cells were cultured in 10% fetal bovine serum containing Dulbecco's modified Eagle's medium and treated with 25 μM resveratrol, 5 μM genistein and the combination (5 μM of resveratrol+5 μM genistein), for 24, 48, and 72 h. The cell proliferation was monitored by an MTT assay. The percent of viable cells was calculated in comparison to the untreated cells that were taken as 100%. Each data point shown is the mean±SD of four or more experiments.
Caspase-3 and caspase-9 activity assays
Significant increases in the caspase-3 and caspase-9 levels have been observed during the 24 h time period in resveratrol and genistein combination treatment (Fig. 2). Resveratrol (25 μM) has shown a fourfold increase in caspase-3 activity and a sixfold increase in caspase-9 activity in comparison with the control. On the other hand, genistein (5 μM) alone was able to show only a twofold increase in caspase-3 activity and a threefold increase in caspase-9 activity when compared with the control cells (Fig. 2). In addition, we were able to successfully increase caspase-9 and cacepase-3 activities by combining low concentrations of resveratrol (5 μM) with genistein (5 μM) and produce an effect similar to a 25 μM concentration of resveratrol.
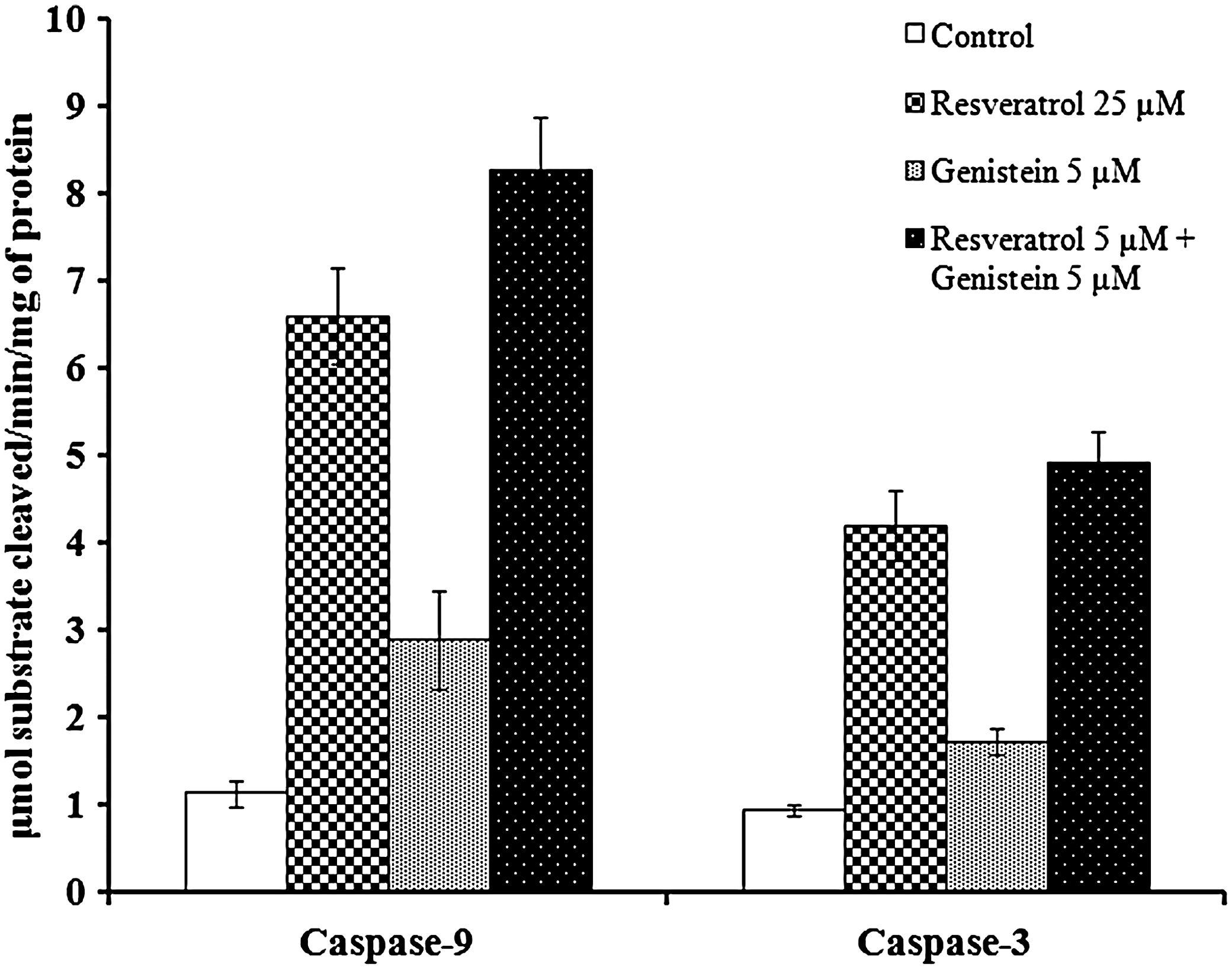
Enzymatic cleavage of Ac-LEHD-pNA and Ac-DEVD-pNA in HeLa cell lines after 24 h after drug treatment. Cell extracts were prepared 24 h after the treatment, as described in the materials and methods section. Caspase-9 and caspase-3 dependent activity using the synthetic tetrapeptide substrates Ac-LEHD-pNA and Ac-DEVD-pNA was determined. Results are the mean±SD from four or more experiments.
Mitochondrial membrane potential
The JC-1 staining technique used in our experiment confirmed the alteration in mitochondrial membrane potential that probably triggered apoptosis during genistein and resveratrol treatments. Changes in the mitochondrial membrane potential of the treated cells revealed that the cell death caused by genistein and resveratrol was indeed preceded by the lowering of the mitochondrial membrane potential which eventually led to the DNA fragmentation of the cancer cells (Fig. 3). This result was seen after 24 h of treatment with resveratrol (5 μM) and genistein (5 μM) combinations. As anticipated, the mitochondria of control cells appeared normal in which the JC aggregates were detected by the red fluorescence; while mitochondrial changes related to apoptotic effects were leaving JC in the monomeric form, and these were detected as green fluorescence. The JC-1 measurement confirmed that we were able to achieve the maximum early-stage apoptotic effect in HeLa cells at the lower concentrations of resveratrol (5 μM) by combination treatment with genistein (5 μM) (Fig. 3).
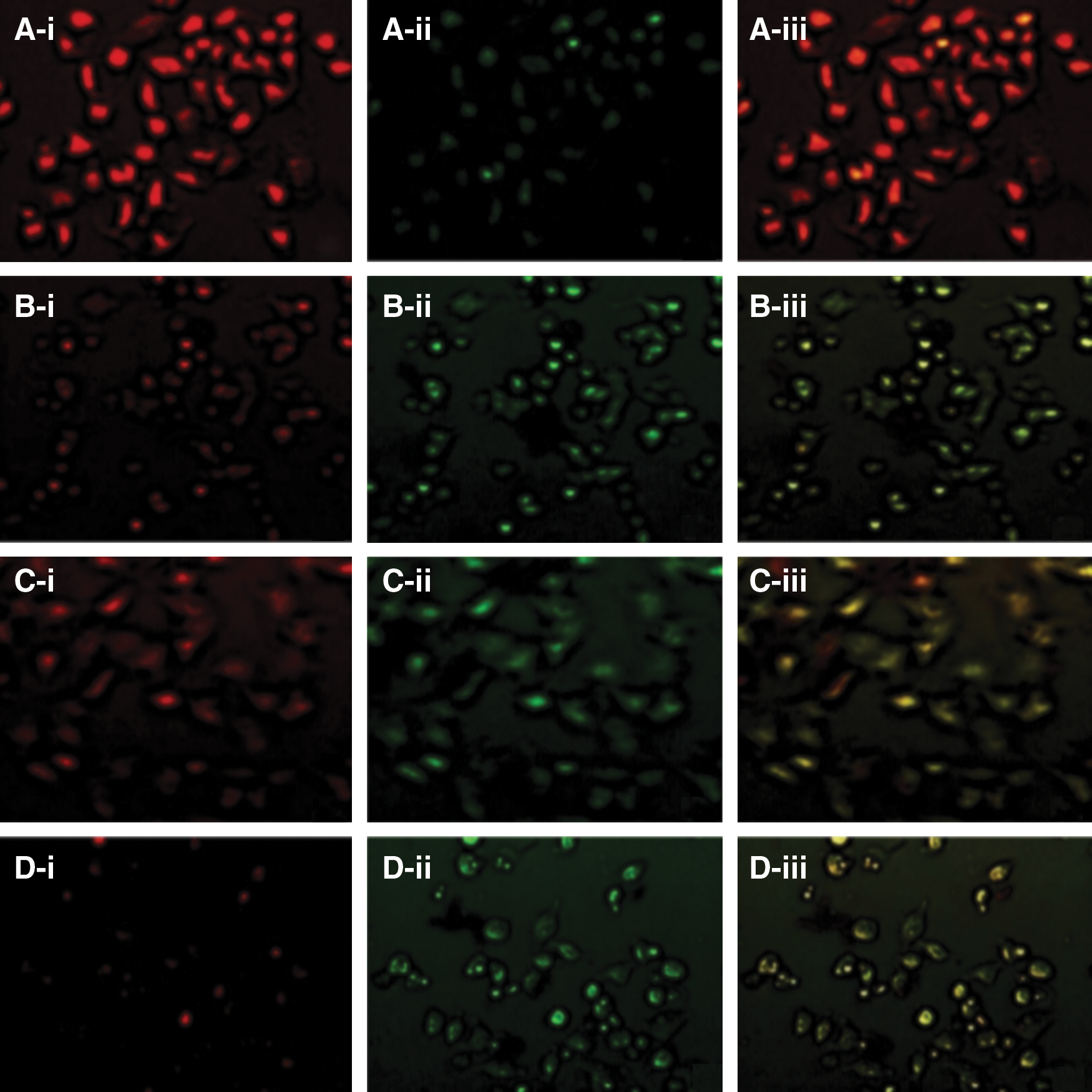
Representative photographs of control and drugs treated HeLa cells stained with JC-1 mitochondrial membrane potential stain after 24 h of treatment:
DNA fragmentation
Apoptosis induction by resveratrol and genistein in HeLa cells was ascertained by using an assay developed to measure DNA fragmentation, a biochemical hallmark of apoptosis. During later stages of apoptosis, internucleosomal cleavage of cellular DNA by endonucleases to 180 bp fragments could be detected by extraction of nuclear DNA and agarose gel electrophoresis. A typical pattern of internucleosomal fragmentation was seen as a stepwise ladder of DNA fragments. The data (Fig. 4) presented in this study show that DNA laddering is pronounced for resveratrol and genistein individually at high concentrations, and the fragmentation is also distinct in combination treatment with low concentrations of HeLa cells. These results further confirmed that when resveratrol and genistein are combined, they can induce a late stage of apoptosis in HeLa cells, thereby causing cancer cell death.
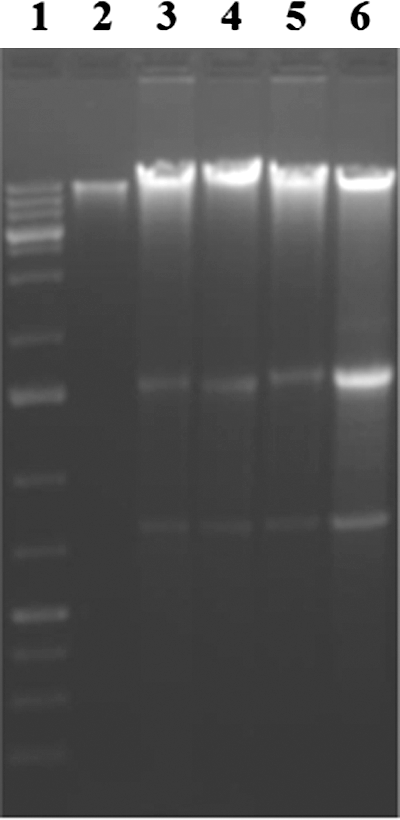
A representative picture of the agarose gel showing the DNA fragmentation in human cervical cancer cells treated with 25 μM resveratrol, 5 μM genistein, and 5 μM of resveratrol+5 μM genistein, for 48 h. Lane 1, 100 bp DNA ladder; lane 2, control cells; lane 3, cells treated with 25 μM resveratrol; lane 4, cells treated with 5 μM genistein; lane 5, cells treated with 5 μM of resveratrol+5 μM genistein; lane 6, cells treated 10 μM topotecan as positive control. The DNA was separated by electrophoresis using 1.5% agarose gel. DNA fragments stained with ethidium bromide were visualized using UVP image analyzer.
Expression of HDM2
The primer set selected for the RT-PCR of HDM2 allowed the detection of full-length HDM2 gene at 1.6 kb and 350 bp ranges. In HeLa cells, 25 μM resveratrol produced 40% reduction at 1.6 kb fragment in HDM2 expression compared with the control (Fig. 5). However, combination treatment of 5 μM resveratrol with genistein (5 μM) was able to produce 78% reduction in HDM2 gene expression at 1.6 kb fragment. In addition, we were able to observe a clear corresponding reduction at 350 bp fragment. These results clearly suggest that combination treatment of resveratrol with genistein could effectively reduce the HDM2 expression in HeLa cells.
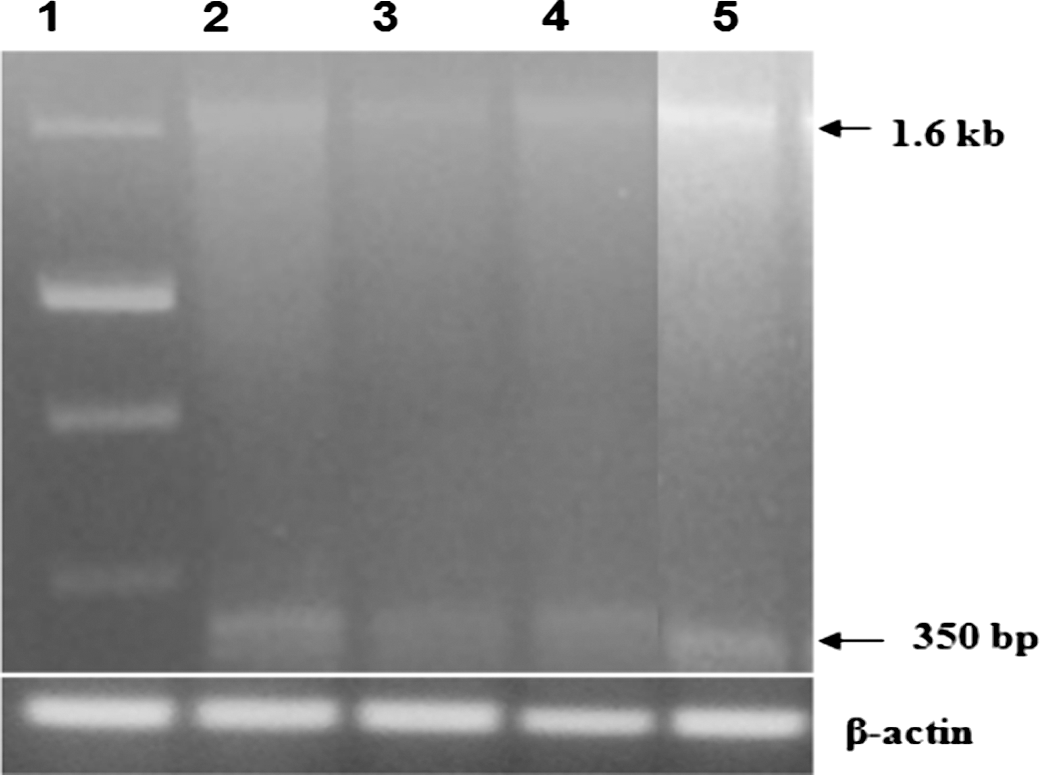
A representative agarose gel picture showing the levels of HDM2 mRNA in resveratrol- and genistein-treated HeLa cells that were amplified by RT-PCR. Lane 1, molecular weight marker; lane 2, control; lane 3, resveratrol 25 μM; lane 4, genistein 5 μM; lane 5, genistein 5 μM+resveratrol (5 μM). β-actin was reported to confirm the equal amount of sample loading. The experiment was repeated at least thrice.
Discussion
The ability to induce tumor cell apoptosis is an important characteristic of candidate anticancer drugs. The process of apoptosis is characterized by specific intracellular events and consequent morphological changes. So far, many methods have been developed to test apoptosis, including morphological study, biochemical assay, ELISA, flow cytometric analysis, and so on, of which changes in the mitochondrial membrane potential and caspases activation, especially in the early stage of apoptosis, are considered one of the most reliable methods. Previous studies conducted in our laboratory have clearly demonstrated that resveratrol can induce apoptosis in A2780/CP70 ovarian cancer cells during combination treatment with nutlin-3 and TGF-β. 20 In the present study, both resveratrol and genistein were found to have moderate to maximum cytotoxic effects on HeLa cells at the concentration ranges tested. Resveratrol has been shown to induce apoptosis in the A2780/CP70 cell line at 25 μM concentration. 21 In our experiments, the treatment of HeLa cells with a lower concentration of resveratrol (5 μM) in combination with genistein (5 μM) showed strong apoptotic effects starting from 24 h of treatment. This indicates that the concentration of resveratrol required in monotherapy is at least five times higher than the concentration typically used for effectively inducing apoptosis in cancer cells. 22
In 2001, Dörrie et al. 23 have reported that 50 μM of resveratrol revealed a four- to ninefold increase of caspase-9 activity in lymphoblastic leukemia cells, which is similar to our results presented here. Caspase-3 is an effector caspase that plays a central role in apoptosis. In our experiment, resveratrol alone and in combination with genistein induced caspase-3 as well as caspase-9 activity in HeLa cells, which are in full agreement with an earlier report, 24 that showed treatment of B16 murine melanoma cell lines with 60 μM of genistein for 72 h. Mitochondrial dysfunction such as loss of mitochondrial membrane potential (MMP) has been reported to be an early event that occurs after the induction of apoptosis. 25 Mitochondrial changes are typically rapid and irreversible events that are preceding other manifestations of the apoptosis process, such as PS (Phosphatidyl serine) externalization, chromatin condensation, and nuclear DNA cleavage. 26,27 It appears that intracellular events involved in the selective killing of cancer cells during our experiments after resveratrol and genistein treatment also involved mitochondrial dysfunction and changes in the mitochondrial membrane potential as revealed by the JC-1 aggregation. As mentioned earlier, JC-1 is a widely used dye to detect mitochondrial depolarization that occurs in the early stage of apoptosis. In the mitochondria of healthy cells, due to high MMP, JC-1 forms J-aggregates that can emit red fluorescence when they are excited at 540 nm. However, immediately after the onset of apoptosis, due to low MMP the JC-1 remains in the monomeric form that can emit green fluorescence; thus, the ratio of red to green fluorescence correlates to the ratio of high-to-low MMP. 28
The appearance of early apoptotic alterations after resveratrol and genistein treatments led us to focus on the hallmark of apoptosis, namely DNA fragmentation. Observations from our experiments clearly suggested that the mode of cell death induced by resveratrol and genistein treatment is mainly due to the collapse in the mitochondrial membrane potential. However, we attempted to identify subsequent events in the HeLa cells that completed the apoptotic process. Typically, low mitochondrial membrane potential causes the release of pro-apoptotic factors such as cytochrome C from mitochondria to cytosol, where it binds to Apaf-1 to form a procaspase-9 activating heptameric protein complex named apoptosome. During this process, cytochrome C binding to Apaf-1 induces the hydrolysis of dATP to dADP, which is subsequently replaced by exogenous dATP. The dATP hydrolysis and exchange on Apaf-1 are two required steps for apoptosome formation. 29 Such apoptosomes are responsible for the activation of caspase-9 through autocatalysis. Subsequently, the central executioner of apoptosis, namely caspase-3, is generated from the procaspase-3 through the activation of caspase-9 by the apoptosome. 30 Activation of caspase-3 eventually triggers the caspase-activated DNase, which enters the nucleus and thus causes DNA cleavage. 31 As a result, a typical internucleosomal DNA fragmentation and “ladder” formation was confirmed in HeLa cells after the treatments just mentioned. Thus, our speculation about mitochondrial membrane potential deregulation leading to DNA fragmentation and the consequent cell death during resveratrol and genistein treatment was validated with sufficient evidence. Interestingly, these results are similar to carvacrol-induced apoptosis that was previously reported in cervical cancer cells. 32
Another important additive to the apoptotic mechanism triggered by resveratrol and genistein treatment is the suppression of HDM2. As a negative regulator of p53, HDM2 is known to transform the cells into more aggressive phenotypes, independent of p53 status. 33,34 In support of this independent tumorigenic transforming potential of HDM2, it has been recently shown that at least 40 alternatively and aberrantly spliced transcripts of HDM2 mRNA have been identified in tumors so far, which presents limitless potential for producing HMD2 isoforms that impact various functions at the cellular level. 35 However, in the current study, we are reporting the detection of a 350 bp splice variant of HDM2 apart from the full-length HDM2 (1.6 kb) in HeLa cells (Fig. 5) that is getting altered during the treatment. The decrease in the expression of these two HDM2 splice variants observed during resveratrol and genistein treatment indicates that the blocking of the cell-cycle progression mediated by HDM2 could be a part of the apoptosis-causing mechanism. Our current observation is suggesting the possibility of stopping cell division by lowering HDM2 expression.
Taking these results together, it appears that resveratrol, a polyphenol, and genistein, an isoflavone, in combination can produce significant cytotoxic effects on HeLa cells and induce mitochondrial changes as was revealed by JC-1 fluorescent staining and DNA fragmentation. It is also evident that resveratrol and genistein can depolarize the mitochondria, individually or in combination, and this mitochondrial dysfunction can possibly contribute to the activation of caspase enzymes. The sequential activation of caspase-3 may eventually cleave the cellular and nuclear components, thus causing DNA fragmentation. At the same time, resveratrol and genistein may inhibit the expression of HDM2 splice variants, causing p53 activation and cell cycle arrest. Thus, the present study demonstrates that resveratrol and genistein, in combination, show stronger activation of caspase-9 and caspase-3 at lower doses and thereby pave the way for apoptotic cell death. These type of combinations of natural products possess the potential to become anticancer treatment tools, particularly in cases where complimentary therapies are required. Further animal studies and clinical trials are needed to fully evaluate therapeutic efficiencies of these combination approaches.
Footnotes
Acknowledgments
The financial support rendered by the President's Faculty Research and Development Grant of Nova Southeastern University is gratefully acknowledged. The authors would also like to thank Royal Dames of Cancer Research, Inc. (Ft. Lauderdale, FL, USA) for their financial support in conducting this research.
Author Disclosure Statement
No competing financial interests exist with this work for any of the authors who are involved in this study.