
Abstract
Estrogens, and particularly glucuronides such as ethinylestradiol (EE), have been shown to cause cholestasis in animal studies, by reducing bile acid uptake by hepatocytes. The aim of the present article was to investigate anticholestatic activity of resveratrol (RES) against liver cholestasis induced by EE in adult female rats. The daily oral administration of the RES at a concentration of 25 mg/kg body weight for 15 days to rats treated with EE (100 μg/kg body weight for 5 days) resulted in a significant protection against EE-induced decrease in both serum cholesterol and bile acid levels as well as against an increase of serum bilirubin concentration. The treatment also resulted in a significant increase in hepatic superoxide dismutase, glutathione peroxidase, glutathione reductase, and catalase activities as well as hepatic protein-bound and nonprotein sulfhydryl groups. RES inhibited serum alkaline phosphatase, alanine aminotransferase, pi-glutathione-S-transferase, gamma-glutamyl transpeptidase, and alpha-glutathione-S-transferase activities, as well as reduced serum tumor necrosis factor-alpha, nitric oxide, and hepatic malondialdehyde as compared to EE-treated rats. The results clearly suggest that RES has a powerful prophylactic action in cholestasis induced by EE. Taken together, RES has potential as a preventive and therapeutic agent for cholestasis and deserves clinical trial in the near future as an adjuvant therapy in women treated with estrogen.
Introduction
C
Materials and Methods
Chemicals
EE was obtained from Sigma Chemical Co. (St. Louis, MO, USA). RES was purchased from Nutrabio Co. (Middlesex, NJ, USA). All other chemicals used in this study were of the analytical grade, preserved under standard situation, and were provided from standard commercial suppliers. Propylene glycol was produced by Panreac Quimica, SA (Barcelona, Spain).
Experimental setup
This experiment was carried out to examine the prophylactic effect of RES against EE-induced liver cholestasis. This experiment was conducted in accordance with guidelines established by the Animal Care and Use Committee of October 6th University. Adult female albino rats weighing around 200±20 g were purchased from the Faculty of Veterinary Medicine, Cairo University (Cairo, Egypt). They were individually housed in cages in an air-conditioned room with a temperature of 22°C±2°C, a relative humidity of 60%, and an 8:00 a.m. to 8:00 p.m. light cycle. During the acclimatization period, each animal was raised on a regular diet (Dyets, Inc., Bethlehem, PA, USA) ad libitum.
The animals were divided into six groups with eight animals in each. The treatment groups are described in Table 1. 25,26
b.w., body weight; EE, ethinylestradiol; RES, resveratrol.
Biochemical assays
Serum level of cholesterol, 27 bile acids, 28 bilirubin, 29 alanine aminotransferase (ALT), 30 alkaline phosphatase (ALP), 31 and gamma-glutamyl transpeptidase (γ-GT) 32 were determined using Reflotron® Plus Analyzer and Roche kits, respectively. Serum pi-glutathione-S-transferase (π-GST), α-GST, and tumor necrosis factor-alpha (TNF-α) were quantitatively estimated by enzyme-linked immunosorbent assay (ELISA) according to Platz et al., 33 Flendrig et al., 34 and Corti et al., 35 respectively. Serum nitrate concentration as a stable end product of NO was estimated by the Griess reaction after quantitative conversion of nitrate to nitrite by nitrate reductase according to the method of Moshage et al. 36
Measurement of lipid peroxidation
A thiobarbituric-acid-reactive substance assay kit (ZeptoMetrix) was used to measure the lipid peroxidation products, malondialdehyde (MDA) equivalents. 37 In brief, liver tissues were homogenized with 0.1 M sodium phosphate buffer (pH 7.4). One hundred microliters of homogenate was mixed with 2.5 mL reaction buffer (provided by the kit) and heated at 95°C for 60 min. After the mixture had cooled, the absorbance of the supernatant was measured at 532 nm using a spectrophotometer. The lipid peroxidation products are expressed in terms of MDA equivalents.
Measurement of antioxidant enzymes
Superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione reductase (GR), and catalase (CAT) activities were determined using commercially available assay kits (BioDiagnostics Inc., River Falls, WI, USA). Briefly, liver tissues were weighed and homogenized with appropriate buffers (provided by the kits). The homogenates were then determined following the procedures provided by the respective manufacturers. The Superoxide Dismutase Assay Kit utilizes a tetrazolium salt for detection of superoxide radicals generated by red formazan dye reduction produced. 38 One unit (U) of SOD activity is defined as the amount of enzyme needed to exhibit 50% dismutation of the superoxide radical. The Glutathione Peroxidase Assay Kit measures GPx activity indirectly by a coupled reaction with GR. 39 Oxidized glutathione, produced upon reduction of hydroperoxide by GPx, is recycled to its reduced state by GR and NADPH. The oxidation of NADPH to NADP+ is accompanied by a decrease in absorbance at 340 nm. Under conditions in which the GPx activity is rate limiting, the rate of decrease in the A340 is directly proportional to the GPx activity. One unit (U) of GPx activity is defined as the amount of enzyme that will cause the oxidation of 1.0 nmol of NADPH to NADP+ per minute at 25°C. The specific activities of the various enzymes in the rat liver are expressed in U/μg of the protein with the protein content determined as stated above. GR activity was assayed as described by Horn, 40 with some modifications, by measuring the oxidation of NADPH at 340 nm. The reaction mixture consisted of 0.1 M sodium phosphate buffer (pH 7.5), 1 mM ethylenediamine tetraacetic acid (EDTA), 0.63 mM NADPH, and 0.15 mM GSSG.
The Catalase Assay Kit utilizes the peroxidative function of CAT for determination of enzyme activity. 41 The method is based on the reaction of the enzyme with methanol in the presence of an optimal concentration of H2O2. The generated formaldehyde is assayed spectrophotometrically with 4-amino-3-hydrazino-5-mercapto-1,2,4-triazole as the chromogen. One unit (U) of CAT activity is defined as the amount of enzyme that will cause the formation of 1.0 nmol of formaldehyde per minute at 25°C.
Estimation of sulfhydryl groups
The activity of nonprotein (NP) sulfhydryl (SH) group was measured according to the method of Sedlak and Lindsay. 42 Liver tissue was homogenized in ice-cold 0.02 M EDTA. Aliquots of 5 mL of the homogenates were mixed in 15-mL test tubes with 4 mL of distilled water, and 1 mL of 50% trichloroacetic acid was added. The tubes were shaken intermittently for 10–15 min and centrifuged at 3000 g. Two milliliters of supernatant was mixed with 4 mL of 0.4 M Tris buffer, pH 8.9, and 0.1 mL of 0.4% 5,5-dithio-bis-(2-nitrobenzoic acid) (DTNB) was added, and the sample was shaken. The absorbance, using a spectrophotometer, was read within 5 min of addition of DTNB at 412 nm against a reagent blank with no homogenate. Finally, the protein content of liver tissue was measured by applying the method of Lowry et al. 43
Histological assessment
The liver was sliced, and pieces were fixed in 10% buffered formaldehyde solution for histological study. The fixed tissues were processed by automated tissue processing machine. Tissues were embedded in paraffin wax by conventional methods. Sections of 5 μm in thickness were prepared and then stained with hematoxylin and eosin for light microscopy analyses according to the method of Bancroft and Steven. 44 After that, the sections were observed under the microscope for histopathological changes, and their photomicrographs were captured.
Statistical analysis
The results were expressed as mean±SD for eight separate determinations. All the data were statistically evaluated with SPSS/13 Software. Hypothesis testing methods included one-way analysis of variance, followed by least significant difference test. P values of<.05 were considered to indicate statistical significance.
Results
Biochemical and antioxidant parameters
Tables 2 and 3 show the specific cholestatic and liver function biomarkers. EE (100 μg/kg body weight [b.w.]) administration for 5 days led to significant increase of biochemical marker levels for ALT (16.17%), ALP (43.66%), pi-GST (67.91%), γ-GT (123.33%), α-GST (63.61%), total bilirubin (103.95%), and MDA (77.39%), while significantly decreasing total cholesterol and bile acids to 36.96% and 35.38%, respectively, as compared with the normal control group (P<.01), indicating acute hepatocyte damage. Pretreatment of animals with RES significantly reduced the level of liver function biomarkers ALT (9.01%, P<.05), ALP (35.27%, P<.01), and bilirubin (44.56%, P<.01) and antioxidant parameters π-GST (23.30%, P<.01), γ-GT (41.57%, P<.01), α-GST (32.14%, P<.01), and MDA (36.27%, P<.01), and also significantly increased cholesterol and bile acids to 42.29% (P<.01) and 40.47% (P<.01), respectively, as compared with the EE group. Likewise, post-treatment of animals with RES significantly reduced the level of liver function biomarkers ALT (4.70%, P<.05), ALP (27.41%, P<.01), and bilirubin (27.17%, P<.01) and antioxidant parameters π-GST (11.88%, P<.05), γ-GT (25.80%, P<.05), α-GST (31.67%, P<.01), and MDA (8.82%, P<.05), and significantly increased cholesterol and bile acids to 47.99% and 23.80%, respectively, as compared with the EE group. The cholestatic effect of EE was controlled in the rats treated with RES (25 mg/kg) as demonstrated by the restoration of the levels of the liver biomarkers. The effect was more pronounced in the case of simultaneous administration of RES pretreatment compared to administration of RES post-treatment.
Values are given as mean±SD for groups of eight animals each.
RES and EE groups were compared with normal control B rats.
RES+EE and EE+RES groups were compared with EE-treated rats.
Values are statistically significant at * P<.05, ** P<.01.
ALP, alkaline phosphatase; ALT, alaninamino transferase.
Values are given as mean±SD for groups of eight animals each.
RES and EE groups were compared with normal control B rats.
RES+EE and EE+RES groups were compared with EE-treated rats.
Values are statistically significant at * P<.05, ** P<.01.
GST, glutathione-S-transferase; GT, glutamyl transpeptidase; MDA, malondialdehyde.
Tables 4 and 5 show serum TNF-α and nitric oxide (NO) and liver SOD, GPx, GR, and CAT levels in different groups of rats. In the EE group, the serum TNF-α and NO levels were twofold higher than in the normal control group (P<.01). Pretreatment with the RES (25 mg/kg b.w.) reduced serum TNF-α and NO levels to 47.74% and 29.84%, respectively (P<.01). Likewise, post-treatment with the RES reduced serum TNF-α levels and NO to 34.76% and 30.92% (P<.05), respectively, as compared with the EE-treated group. In addition, significantly (P<.01) decreased activities of liver antioxidant enzymes were observed in the EE-treated rats compared with the normal control group (P<.01). RES pre- and post-treatment at 25 mg/kg b.w. significantly (P<.01, P<.05) enhanced the liver antioxidant enzymes activities (SOD, GPx, GR, and CAT) in rats, respectively, as compared to the EE-treated group. As shown in Table 6, a marked decrease in levels of hepatic total, protein-bound, and nonprotein sulfhydryl groups was recorded after EE treatment. Pretreatment of animals with RES (25 mg/kg) significantly elevated the levels of total, protein-bound, and nonprotein sulfhydryl groups in liver homogenate by 68.57%, 66.78%, and 79.66%, respectively, as compared with the EE-treated group. In addition, post-treatment with RES increased hepatic total, protein-bound, and nonprotein sulfhydryl groups significantly (P<.01) by 46.33%, 44.79%, and 56.95%, respectively, as compared with the EE-treated group.
Values are given as mean±SD for groups of eight animals each.
RES and EE groups were compared with normal control B rats.
RES+EE and EE+RES groups were compared with EE-treated rats.
Values are statistically significant at * P<.05, ** P<.01.
NO, nitric oxide; TNF, tumor necrosis factor.
Values are given as mean±SD for groups of eight animals each.
For SOD, one unit of activity was taken as the enzyme reaction which gave 50% inhibition of NBT reduction in 1 min/mg protein.
GPx is reported in μg of GSH consumed each min/mg protein.
GR is reported in nmol of NADPH oxidized each min/mg protein.
CAT is reported in μmol of H2O2 utilized eac min/mg protein.
RES and EE groups were compared with normal control B rats.
RES+EE and EE+RES groups were compared with EE-treated rats.
Values are statistically significant at * P<.05, ** P<.01.
CAT, catalase; GPx, glutathione peroxidase; GR, glutathione reductase; SOD, superoxide dismutase.
Values are given as mean±SD for groups of eight animals each.
RES and EE groups were compared with normal control B rats.
RES+EE and EE+RES groups were compared with EE treated rats.
Values are statistically significant at ** P<.01.
NP, nonprotein; PB, protein-bound; SH, sulfhydryl.
Histopathology examination
Histopathological examination of liver sections of the normal groups (A and B) showed regular cellular architecture with distinct hepatic cells, sinusoidal spaces, and a central vein. The hepatocytes are polygonal cells with a well-preserved cytoplasm and a nucleus with prominent nuclei (Fig. 1a, b). Likewise, in RES-treated animals, liver sections showed normal hepatic architecture (Fig. 1c). On the other hand, in the hepatotoxic-positive EE-treated control group, histological examination showed marked proliferation of the bile ducts. Bile components were not observed in the hepatocytes or the bile canaliculi, hepatic cell necrosis and loss of architecture, inflammation, and congestion with cytoplasmic vacuolation, fatty change, sinusoidal dilatation, centrilobular necrosis, and displayed bundles of collagen surrounding the lobules, which resulted in huge fibrous septa and distorted tissue architecture (Fig. 1d). Histopathological examination also showed good recovery of EE-induced necrosis by RES as compared with the EE-treated group. RES pre- and post-treatment at 25 mg/kg b.w. showed remarkable histological regeneration compared to those of the EE group (Fig. 1e, f). They showed nearly ordinary patterns with an increase normal hepatocyte parenchyma and a reduced development of fibrous septa and lymphocyte infiltration. Results of the gross and histopathological examination are shown in Figure 1.
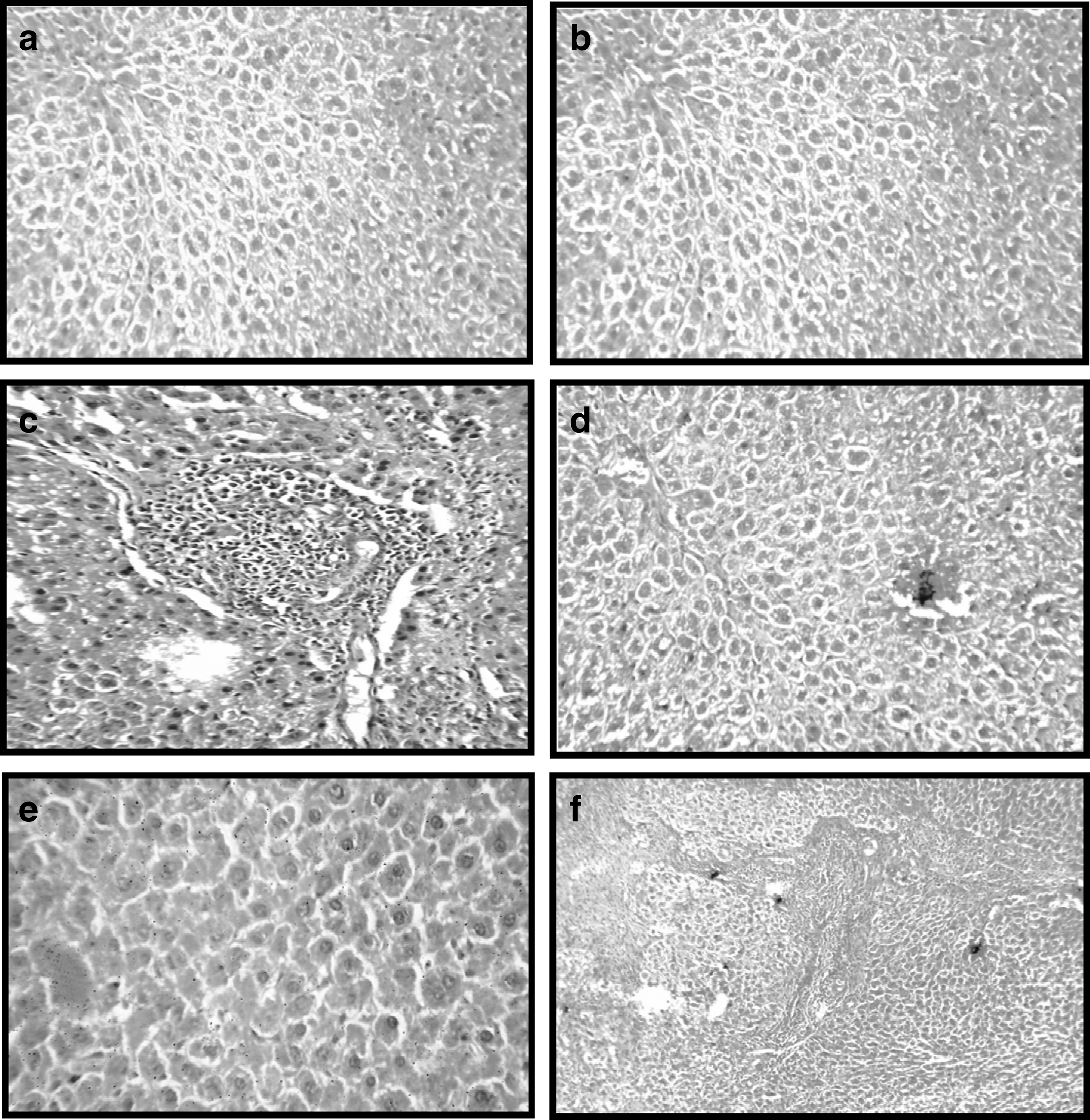
Effect of resveratrol (RES) on the histopathology of liver in rats treated with ethinylestradiol (EE).
Discussion
Estrogens are well-known to cause reversible intrahepatic cholestasis in susceptible women during pregnancy, administration of oral contraceptives, and postmenopausal replacement therapy. 4 In the present study, rats exposed to EE (100 μg/kg b.w.) had significant reductions in serum cholesterol and bile acids and elevations of biochemical markers ALT, ALP, π-GST, γ-GT, and α-GST, total bilirubin, and MDA levels (Tables 2 and 3). Liver cholestasis is associated with cellular necrosis 45 and increases in tissue lipid peroxidation resulted in oxidative stress as MDA, ALT, ALP, π-GST, γ-GT, α-GST, and bilirubin were elevated. Cholestatic activity of EE appears to act through reduction of biliary excretory activity of the canalicular membrane 46 and decreased elimination of conjugated bilirubin and an increase in the conjugated level in plasma, a symptom diagnostic for obstructive jaundice. 47 EE was reported to decrease serum cholesterol level, which is accompanied with an increase in hepatic cholesterol level. 48 The hypocholesterolemic effect of EE appears to act through stimulating low-density lipoprotein receptor activity, increasing the binding of lipoproteins to the liver plasma membrane, 49 and increasing hepatic catabolism of low-density lipoprotein. 50 Also, the decrease in bile flow due to cholesterol precipitation may be responsible for the reduced level of bile acids in serum after EE administration. 51 In the present study, oral administration of RES (25 mg/kg b.w.) showed significant protection against induced decrease in serum cholesterol and bile acids through inhibiting squalene monooxygenase, a rate-limiting enzyme in cholesterol biosynthesis. Also, RES stimulates cholesterol-7α-hydroxylase activity, which is responsible for bile acids synthesis and reduces liver cholesterol levels. 52,53
The present results show that RES could inhibit serum TNF-α and NO levels in the EE-treated group (Table 4). Free radicals are involved in the regulation of cell proliferation and death, as well as gene expression such as TNF-α, NO, and MDA. 54 Evidence indicates that free radicals, oxidative stress, and lipid peroxidation are present in cholestatic damage. 55,56 EE causes changes in the equilibrium between antioxidant and pro-oxidant activity, favoring the latter, since it increased production of hepatic MDA and reduced free-radical–scavenging activities. It has been shown that in chronic cholestasis, the increased intrahepatic concentration of bile acids induces mitochondrial toxicity and free-radical generation. 57 TNF-α, TGF-β1, and interleukin-6 are the most extensively studied mitogenic and fibrogenic factors. RES is also able to inhibit proinflammatory cytokine expression. 58 Taken together, these results indicate that the antifibrotic effect of RES is associated with the blockade of mitogenic and/or fibrogenic signaling. TNF-α was reported to induce NO formation. 59 The increased NO production is recognized as an important mediator of physiological and pathological processes, 60 as a result of these inflammatory and destructive processes. In addition, RES is a potent reactive oxygen species (ROS) scavenger 61 and normalized the oxidative stress biomarkers (π-GST, γ-GT, α-GST, NO, and MDA), resulting in reduced oxidative stress, which contributes to suppression of hepatocyte inflammation by EE. In the present study, the significant decrease in hepatic SOD, GPx, GR, and CAT activity were detected after EE administration (Table 5). The reductions of hepatic SOD and CAT activities in EE-induced cholestasis rats when compared with normal rats were reported in this study due to production of NO; as a result of inflammation and destructive processes, 60 the oxidative stress on the hepatic cells was increased leading to depletion of antioxidant enzymes that scavenge the toxic superoxide and hydrogen peroxide radicals that promote lipid peroxidation, whereas the RES-treated groups showed a significant increase in the hepatic SOD, GPx and GR, and CAT activities of the EE-induced cholestatic rats (Table 5). It was reported that the in vivo antioxidant property of RES probably arises from its ability to protect against the molecular effects of lipid peroxidation, free radicals, and ROS, and it also delays the progress of many chronic diseases. 62,63 It has also been reported that the antioxidant role of RES can reduce the risk of CVD, likely through its prevention of low-density lipoprotein oxidation and modulation of lipid metabolism, as well as inhibition of eicosanoid synthesis and platelet aggregation. 64 –66 Since then, it has been investigated as an effective treatment for metabolic complications. 67
As an indicator of oxidative stress, liver content of total, protein-bound, and nonprotein SH groups (Table 6). The NP–SH concentration in the liver tissue decreased significantly in the EE-treated rats, while RES administration prevented the decrement in NP–SH induced by EE. RES contributed significantly to the intracellular antioxidant defense system by acting as a powerful consumer of singlet oxygen and hydroxyl radicals. Obviously, the reduced level of NP–SH groups is due to the involvement of reduced glutathione in rendering the harmless estradiol metabolites by the formation of conjugates. Treatment with RES results in stimulation of hepatic total, protein-bound, and nonprotein sulfhydryl group biosynthesis. These results were agreement by the finding of Lixian et al., 68 who demonstrated a significant increase in reduced glutathione by treatment with RES. Finally, histopathological examination showed a marked degree of bile duct proliferation in EE-treated rats (Fig. 1d). Comparing the beneficial effect of RES with that of EE-induced cholestasis, RES showed anticholestatic activity as indicated by the measured biochemical parameters and the histopathological examination of liver. In the present study, the histological findings prove that RES affected the recovery of the liver structure in rats with EE-induced liver cirrhosis. Indeed, there was remarkable reduction in fibrosis extent and a decrease of stellate infiltration in rats treated with RES groups compared to the control EE group. Histological studies confirmed the hepatoprotective effect of RES. EE-treated rat liver sections showed fatty degeneration of hepatocytes and necrosis of cells. RES treatment (25 mg/kg) almost normalized these effects in the histoarchitecture of the liver. Furthermore, the severe fatty changes in the livers of rats caused by EE were treated in the EE treatment groups. Therefore, from this study, the RES could be a hepatoprotective against EE-induced liver damage in rats. In addition, the most novel and relevant finding was that RES supplementation was accompanied by the alleviation of bile duct proliferation and ductular reaction in this model. RES was also able to reduce newly formed bile ducts (Fig. 1). Since the proliferation of bile ducts is an early event in cholestasis-related changes, the attenuation of hepatic injury and fibrosis in rats by RES might be associated with alleviation of ductular reaction. Prophylactic effect of RES against liver cholestasis induced by EE has not been reported earlier to my knowledge, and this study is perhaps the first observation of its kind.
In conclusion, the present study showed that RES has powerful anticholestatic activity against liver cholestasis induced by EE, in addition to its antioxidant action and free-radical-scavenging activities.
Footnotes
Acknowledgment
Thanks to Dr. Ahmed Saed (Faculty of Applied Medical Sciences, October 6th University) for his involvement in histopathological examination, and Mr. Abdelkhalek Ahmed for animal care assistance.
Author Disclosure Statement
No competing financial interests exist.