
Abstract
Hypoxia induces a wide range of deleterious effects at the cellular level due to an increased production of reactive oxygen species (ROS). Polyphenols from grape seeds, which are potent antioxidants might protect the brain against oxidative stress produced by hypobaric hypoxia. The brain effects of three doses of grape seed extract intraperitoneally (i.p.) administered in rats after exposure to hypobaric hypoxia corresponding to 5500 m altitude were investigated. Some oxygen and nitrogen reactive species, inflammatory cytokine (IL-6) and molecules involved in angiogenesis (vascular endothelial growth factor [VEGF], matrix metalloproteinase 2 [MMP2], and tissue inhibitors of metalloproteinase 1 [TIMP1]) were determined. Forty-two rats were divided in seven groups: group 1, control; groups 2, 3, and 4 were exposed to hypobaric hypoxia for 24 h in a hypobaric chamber; groups 5, 6, and 7 were exposed to hypobaric hypoxia for 5 days. After returning to normal atmospheric pressure, rats from groups 2 and 5 were sacrificed without other treatment. Animals from groups 3 and 6 were i.p treated with carboxymethyl cellulose (CMC) vehicle and those from groups 4 and 7 were i.p. treated with grape seed extract (GSE) (50 mg gallic acid equivalents/kg body weight in 0.5 mL CMC suspension/animal). The treatment was applied at 2, 24, and 72 h from returning to normoxia. Hypobaric hypoxia produced increased brain levels of ROS, nitric oxide (NO), IL-6, and VEGF after both time intervals (P<.05). The MMP2 concentration was significantly increased in groups treated only with vehicle, whereas TIMP1 was slightly changed. GSE produced a significant reduction of ROS and NO levels proving its antioxidant capacity. It also decreased IL-6 and MMP2 concentrations to values similar to controls. The VEGF concentration was also significantly reduced. These effects are indicative for anti-inflammatory and antiangiogenic properties of GSE.
Introduction
T
It has been reported that hypoxia causes oxidative stress involving both reactive oxygen species (ROS) and reactive nitrogen species (RNS). 3,4
Free radicals act as signaling species in various normal physiological processes. Excessive production of free radicals causes damage to biological material and is an essential event in the etiopathogenesis of various diseases. 5,6
The effects of ROS can be particularly evident in certain tissues such as brain. Because the brain consumes about 20% of the basal oxygen, more than other tissues also undergoing mitochondrial respiration, the potential for ROS exposure is increased. The brain is particularly affected by oxidative species because it has a high content of polyunsaturated fatty acids, which easily undergo oxidation. In addition, the presence of redox-active metals (Cu, Fe) increases the ROS level. On the other hand, the brain has a relatively low level of antioxidant capacity. 7
In the last years, much effort has been undertaken to develop therapies to reduce the effects of oxidative stress, which either causes or aggravates different diseases.
Considerable evidence now shows that antioxidants can exert protecting action on a variety of illnesses, including heart disease, cancer, and neurodegenerative diseases. There are many investigations, which suggest that polyphenolic compounds contained in fruits and vegetables are powerful antioxidants, metal chelators, and cell signaling molecules. They have a broad spectrum of pharmacological and therapeutic effects as a result of their anti-inflammatory or antimicrobial activities. 8,9 Although the mechanism of action of polyphenols is disputed by certain investigators, in vitro, in vivo, and epidemiologic studies suggest that polyphenols possess beneficial health properties.
Phenolic compounds from grapes, wine, and grape seeds are very potent antioxidants and have a high ability to scavenge free radicals. Therefore, these compounds might exhibit positive effects on health. 10,11
The aim of the present study was to investigate the effects of grape seed extract (GSE) administration after exposure to hypobaric hypoxia on the brain levels of some oxygen and nitrogen reactive species and inflammatory cytokine and molecules involved in the angiogenesis process.
Materials and Methods
Reagents
Phenylmethanesulfonyl fluoride (PMSF), Tris-(hydroxymethyl)aminomethane (TRIS), glucose-6-phosphate, glucose-6-phosphate dehydrogenase, NADPH-dependent nitrate reductase, β-nicotinamide adenine dinucleotide phosphate reduced tetrasodium salt (NADPH), N-(1-naphthyl)ethylene diamine hydrochloride, sulfanyl-amide, 2′,7′-dichlorofluorescein–diacetate (DCF-DA), 2′,7′-dichlorofluorescein (DCFH) were purchased from Sigma-Aldrich (Sigma-Aldrich Chemicals GmbH, Inc., Seelze, Germany), orthophosphoric acid from Merck KgA (Darmstadt, Germany), KH2PO4, K2HPO4.3 H2O were from Chimopar (Bucharest, Romania). IL-6, vascular endothelial growth factor (VEGF), and tissue inhibitors of metalloproteinase 1 (TIMP1) ELISA kits were from R&D systems (Minneapolis, MN, USA), and matrix metalloproteinase 2 (MMP2) kit from Abnowa (Taipei City, Taiwan).
Grape seed extract
The hydroethanolic extract from grape seeds (Vitis vinifera L.), variety Burgund Mare (Romania) was prepared from a 0.1 g/mL mixture of finely powdered dried seeds and water/ethanol 50/50 (v/v). 12 The solution was concentrated in vacuo (10-fold). The total polyphenolic content (TPC) of the extract, assessed by the Folin–Ciocalteu colorimetric reaction, was 3400 mg gallic acid equivalents/l. 13 The total catechin content was 2.06 mg/mL. Proantocyanidins represented 35.76% of the TPC with a concentration of 1.073±0.03 mg/mL. Anthocyanidine concentration was 3.17×10−3 mg/mL. HPLC analysis detected peaks with maximums for epigallocatechin and epicatechin at 193 nm, catechin hydrate at 196 nm, procyanidin B at 203 nm, and gallic acid at 212 nm. 14 The antioxidant capacity of GSE evaluated by the ABTS colorimetric test was 52.89±0.02 mM Trolox eq. 15
For this study, GSE was suspended in a 0.5% carboxymethyl cellulose (CMC) solution as vehicle.
Animals
Forty-two male Wistar rats weighing 225±5 g were used throughout the experiment. The animals were housed in the animal facility of the physiology department from the “Iuliu Hatieganu” University of Medicine Cluj-Napoca under a 12-h light/12-h dark cycle and 24°C±1°C temperature and had free access to water and food. All the experiments were in agreement with the guidelines for animal research of the Ethics Committee of “Iuliu Hatieganu” University.
Treatment of animals
Hypobaric hypoxia was accomplished by keeping the rats in an automated hypobaric chamber at a pressure corresponding to 5500 m altitude. Forty-two male Wistar rats were randomly divided in seven equal groups. The control group (1) was kept at normal atmospheric pressure. Animals from groups 2, 3, and 4 were exposed to hypobaric hypoxia for 24 h and those from groups 5, 6, and 7 were exposed for 5 days. Animals from groups 2 and 5 were sacrificed at 1-h intervals after returning to normal atmospheric pressure. Animals from groups 3 and 6 were intraperitoneally (i.p.) injected with three doses of 0.5 mL CMC vehicle/animal. Rats from groups 4 and 7 were i.p injected with three doses of GSE: 50 mg gallic acid equivalents/kg body weight in 0.5 mL CMC suspension/animal. The i.p. treatment was applied at 2, 24, and 72 h after the end of the hypoxia exposure. Rats from groups treated with vehicle or GSE were sacrificed at 24 h after the last injection. The brain was rapidly removed, frozen, and stored at −80°C until assayed.
Whole brain was homogenized in a TRIS (50 mM)-EDTA (10 mM) buffer (pH 7.4) containing 10 mg/mL PMSF with a Polytron homogenizer. The assays were performed in supernates obtained after centrifugation at 9200 g for 15 min.
Estimation of ROS and nitric oxide
Free radical determination
The presence of free radicals in supernates was estimated with the 2′,7′-dichlorofluorescein–diacetate (DCF-DA) method. 16 This method is based on the hydrolysis of DCF-DA by cellular esterases to nonfluorescent 2′,7′-dichlorofluorescin, which is oxidized by ROS in fluorescent DCFH. The readings were performed fluorometrically (excitation wavelength 488 nm and emission at 525 nm). A standard curve was obtained with DCFH (25–400 pmoles/mL) and results were expressed as pmoles DCFH/mg protein.
NO determination
Nitric oxide (NO) was evaluated by estimating the total nitrite level in supernates using the Griess method. Briefly, nitrates were reduced with nitrate reductase to nitrites, amd then proteins were precipitated. After centrifugation, the supernate was treated with the Griess reagent. The absorbance was measured at 543 nm. Nitrite concentrations were established using a standard curve obtained with NaNO2 solutions. The results were expressed in μmol/mg protein. 17
Estimation of IL-6, VEGF, MMP2, and TIMP1 was performed in supernates of brain homogenates using ELISA methods according to the manufacturer's protocol.
The protein content in homogenates was determined using the Bradford method. 18
Statistical analysis
The results are expressed as mean±standard deviation. The data analyses were performed using SPSS13.0 software with the Mann–Whitney U and Kruskal–Wallis nonparametric tests. The criterion for statistical significance was P<.05.
Results
Free radicals
The brain levels of free oxygen radicals after 24 h and 5 days of hypobaric hypoxia were significantly increased (144.7±29.1, respectively, 177.03±28.8 pmoles/mg) comparatively with controls (21.04 pmoles/mg protein) (P<.02). After 5 days of hypoxia, the ROS level was higher compared with the 24-h group, but the difference was not statistically significant. In animals treated only with vehicle, the values declined but remained significantly higher versus controls (93.48±19.6 pmoles/mg at 24 h, respectively, 85.85±13.4 pmoles/mg at 5 days of hypoxia) (P<.05). GSE significantly reduced free radical levels in both treated groups (46.58±11.4 pmoles/mg for the 24-h group and 43.13±12.57 pmoles/mg for the 5-day group) as compared with animals after 24 h and 5 days of hypoxia (P<.02) and those treated only with vehicle (P<.05) (Fig. 1A).
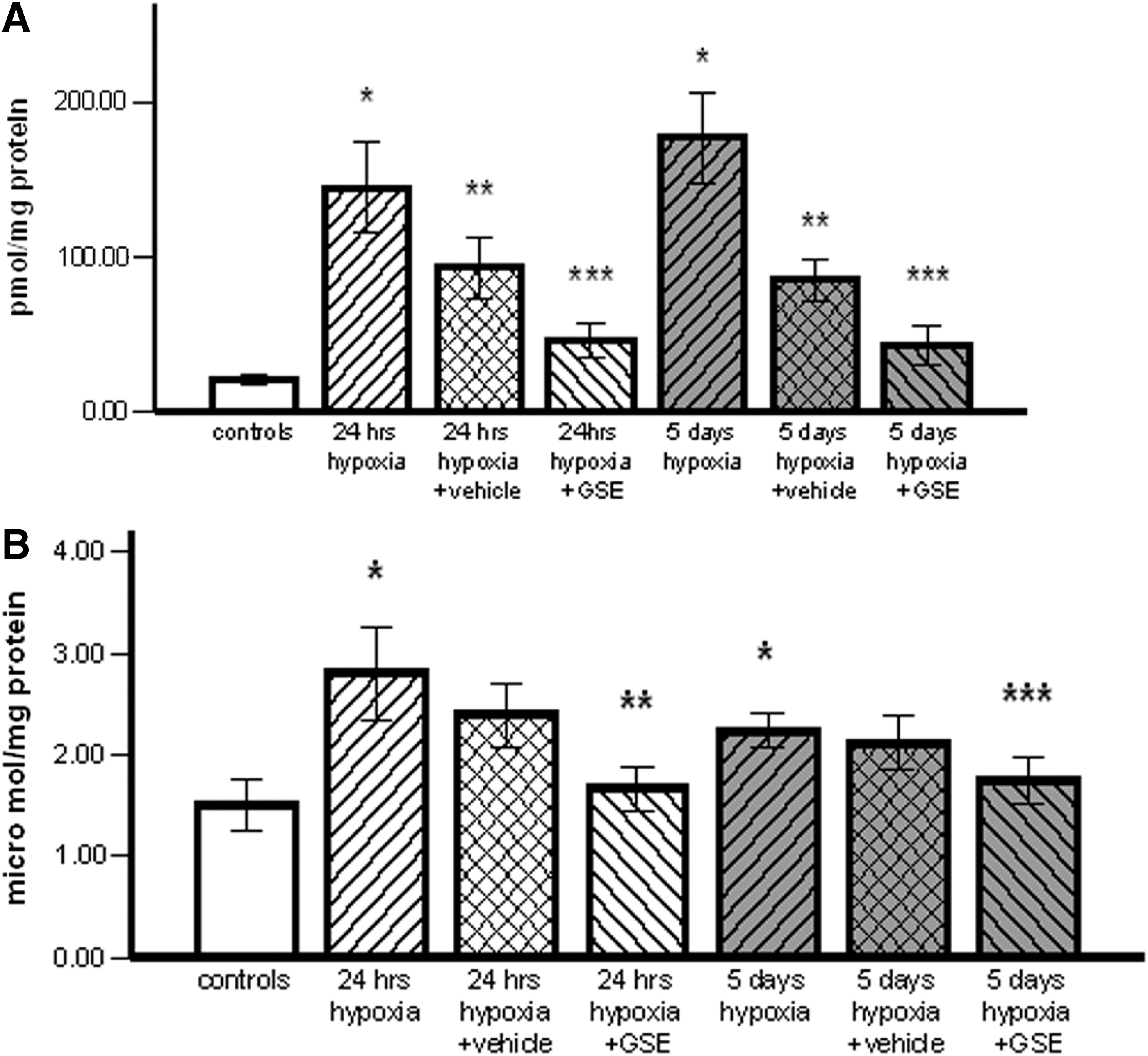
NO is a multifunctional free radical presenting cytotoxic or cytoprotective effects on mammalian cells. In this experiment, hypoxia induced increased brain NO concentrations at both time periods. In the 24-h group, the NO level was insignificantly higher than in the 5-day hypoxia group (2.8±0.46 and 2.23±0.16 μmol/mg protein in the 24-h, respectively, 5-day hypoxia groups vs. 1.5±0.25 μmol/mg protein in controls) (P=.05). NO levels slightly decreased in groups treated only with CMC. GSE treatment induced a significant reduction of NO concentrations in both treated groups: 1.76±0.22 μmol/mg after 24 h of hypoxia (P<.02) and 1.75±0.23 μmol/mg after 5 days of hypoxia (P<.05) (Fig. 1B).
The IL-6 cytokine level, measured in whole brain supernate, was significantly increased after 24 h and 5 days of hypoxia versus controls (55.54±5.1, respectively, 49.61±5.2 vs. 36.04±6.09 pg/mg protein) (P<.02). The difference between the groups exposed only to hypoxia was not significant. IL-6 concentrations insignificantly decreased in groups treated with vehicle. GSE treatment additionally reduced brain IL-6 to 42.94±5.11 and 43.72±7.01 pg/mg after 24 h and 5 days of hypoxia (Fig. 2).

The IL-6 concentration in brain homogenates. Hypobaric hypoxia induced increased brain levels of IL-6 in animals of both 24-h and 5-day groups (*P<.02 vs. controls). GSE treatment determined a decline in the IL-6 concentration.
VEGF has a key role in angiogenesis in health and disease. Hypoxia produced a significant increase (P<.05) of VEGF versus controls (5.1±1.1 pg/mg). At 24 h, the VEGF level (8.9±0.3 pg/mg) was insignificantly higher comparatively to that at 5 days (7.6±2.2 pg/mg). VEGF levels decreased in the brain of rats treated only with CMC, especially after 5 days of hypoxia (P<.05). GSE determined the lowering of VEGF concentrations at both times of hypoxia (6.3±2.5 in the 24-h group, respectively, 1.9±0.4 pg/mg in the 5-day group). The VEGF level was significantly lower in rats treated with GSE after 5 days of hypoxia comparatively to those treated only with vehicle (P<.01) (Fig. 3A).

MMP2 involved in the degradation of the extracellular matrix had an increased level after 24 h (842.0±83.4 pg/mg) and 5 days (818.5±121.9 pg/mg) of hypobaric hypoxia as compared with the normoxia group (659.4±67.9 pg/mg). The difference between these groups was statistically insignificant. The MMP2 level significantly increased in groups treated only with vehicle to 1313.1±73.8 pg/mg in the 24-h group (P<.02) and 936±64.37 pg/mg in the 5-day group (P<.05). The MMP2 concentration decreased in groups treated with GSE (797.94±36.2 pg/mg in the 24-h hypoxia group, respectively, 841.6±31.2 pg/mg in the 5-day group). These levels were higher than those of controls (Fig. 3B).
TIMP1, one of the four tissue inhibitors of metalloproteinases, acts in the control of proteolysis of the extracellular matrix. The TIMP1 levels were insignificantly changed after 24 h and 5 days of hypoxia (44.04±3.7 pg/mg, respectively, 35.4±4.1 pg/mg) comparatively with normoxia controls (48.6±10 pg/mg). In both groups treated with vehicle after hypobaric hypoxia, the levels were similar: 43.4±7.8 for 24 h of hypoxia, respectively, 35.7±3.4 pg/mg for 5 days of hypoxia. TIMP1 concentrations insignificantly increased in both the GSE-treated groups (56.2±12.6 and 56.2±11.4 pg/mg for 24 h of hypoxia, respectively, 5 days of hypobaric hypoxia) (Fig. 3C).
Discussion
Hypobaric hypoxia affects the central nervous system leading to many physiological alterations: mental retardation, memory deficit, and motor impairment. 3,19 Hypobaric hypoxia was demonstrated to cause oxidative stress, which may be involved in neurodegeneration and memory loss. 20 In hypoxia conditions, oxidative stress produces strong deleterious effects on cells due to damage in lipids, proteins, and DNA molecules. 21,22
Reactive oxygen and nitrogen species were demonstrated to be involved in a large number of normal physiologic and pathological processes. They appear as a consequence of aerobic metabolism and function at the cellular level to modulate the redox status as well as to signal for the generation of different reactions chains.
The increased level of ROS in hypoxia is the result of the accumulation of reduction equivalents in the mitochondrial electron transport system due to the lower level of oxygen. The accumulated electrons attack O2 molecules forming superoxide radical anion, which leads to H2O2 and hydroxyl radicals (HO·−). 3,6,7
Polyphenols, which are broadly distributed in plants are potent antioxidants capable of scavenging free radicals. In this way, they interfere with the initiation and propagation of free radical chain reactions, which might produce oxidative effects on biomolecules. The antioxidant power of these compounds has been shown to be higher than those of vitamin C or E. 8
Many in vivo and in vitro models of neurodegenerative disorders, which are associated to oxidative stress (e.g., Alzheimer's disease, Parkinson's disease), evidenced that polyphenols had neuroprotective effects due especially to their antioxidant properties. 23 –25 Polyphenolic compounds have also the capacity to chelate iron. 26 In this way, ROS production, n as a result of catalytic processes involving redox–active metals, is prevented. Additionally, these compounds contribute to the induction of antioxidant enzymes. 27 All these beneficial effects of polyphenols in brain hypoxia and neurodegenerative diseases are linked to their capacity to permeate the blood–brain barrier. 28,29
The present study reports that hypobaric hypoxia corresponding to an altitude of 5500 m resulted in an increase of free radical levels in the brain after 24 h. A higher level was found after 5 days of hypoxia, but the difference between these two groups of animals was not statistically significant. Another study revealed that hypobaric hypoxia in rats determined time-dependent increasing of ROS levels. 3 Additionally, we observed that in groups injected with vehicle although ROS levels decreased, significantly higher levels of ROS compared to the normoxia group were present even after 96 h from the ending of the hypoxia period. CMC injected as vehicle in this and other experiments 30,31 is a physiologically inert polymer. Therefore, the decline in the parameter level measured in the vehicle-treated groups is a result of their natural reduction after the returning to normoxia.
Oxidative stress involves also RNS. The nitric monoxide system was demonstrated to be involved in increasing RNS after hypobaric hypoxia. 22,32,33 NO is a multifunctional free radical presenting an important role as a messenger for physiologic processes like immune response, vasodilatation, and neurotransmission. Pathophysiological processes, such as neurodegeneration or inflammation, also involve NO action.
Hypoxia leads to an increased Ca2+ level in cells as a result of glutamatergic stimulation. Neuronal nitric oxide synthase (n-NOS) becomes activated and induces an increased synthesis of NO. This results in an increased oxidative stress producing DNA damage and leads to cell death. NO can also react with proteins forming nitrosylated and nitrated products. 22
In this experiment, significantly increased concentrations of NO were evidenced in the rat brain just after 24 h. They were insignificantly lower after 5 days of hypobaric hypoxia (Fig. 1). In the other study, the highest level of NO was observed at 7 days of hypobaric hypoxia corresponding to 6500 m altitude. 22 These results suggest that the hypoxia level influences the dynamics and the level of the brain NO synthesis.
Neuronal cells seem to be protected against ROS- and RNS-induced damages by naturally occurring polyphenols. This study demonstrates that administration of three doses of GSE determined the reduction of brain ROS. This result is probably a consequence of its antioxidant properties. GSE used in our study had a strong antioxidant capacity in vitro as revealed by ABTS assay. This activity is related to the high content of polyphenols: proanthocyanidins, anthocyanidines, and catechins evidenced in the extract.
Antioxidant and free radical scavenging effects of GSEs were demonstrated by reduction of lipid peroxides and DNA alteration in the animal brain. 34,35 Additionally, red wine polyphenols were demonstrated to inhibit brain NADPH-oxidase activation, an important source of superoxide radical. 10
The reduction of the brain NO level after GSE administration was also evidenced in our study. Probably, this extract had an inhibitory effect on n-NOS a result of its epigallocatechin gallate content. Flavonoid reduction of NO and cytokine concentration involved in neuroinflammation was demonstrated. 36 –38 Resveratrol, oxyresveratrol, and quercetin were proved to protect against NO cytotoxicity. 39,40 Moreover, hypoxia-induced NADPH-d/n-NOS expression was depressed by high doses of epigallocatechin gallate. 41
Hypobaric hypoxia was demonstrated to have a long lasting impact on the immune function. Therefore, studies were undertaken to establish the influence of hypoxia on inflammatory cytokines. It was found that high-altitude hypoxia determined an upregulation of circulating interleukin-6, interleukin-1 receptor antagonist, and C-reactive protein and was suggested that these cytokines are involved in high-altitude pulmonary edema. 42
IL-6, a multifunctional cytokine, is involved in the regulation of brain responses to inflammation and degenerative diseases 43 and in the regulation of neuronal survival. 44 IL-6 becomes elevated in brain injuries and inflammation. 43
In vitro studies demonstrated that hypoxia induces a dramatic increase in the expression of IL-6 of mouse endothelial cells 45 and rat neonatal cardiac myocytes. 46 It was also suggested that IL-6 can promote angiogenesis and may contribute to the induction of VEGF production in hypoxia. 47 In our in vivo experiment, hypobaric hypoxia determined a significant increase of brain IL-6 level, especially after 24 h. Three doses of GSE determined the decline of this cytokine concentration. This effect demonstrates that our extract from grape seeds presents potentially anti-inflammatory properties. It was shown that flavonoids attenuate microglia and/astrocyte-mediated neuroinflammation through modulation of protein and lipid kinase signaling pathways. 48 In vitro and in vivo studies demonstrated that luteolin, a natural flavonoid, inhibits LPS-induced IL-6 production in the brain by inhibiting the JNK signaling pathway and activation of AP-1 in microglia. 49
A compensatory mechanism to increase oxygenation in tissues that underwent hypoxia is the formation of new blood vessels. Angiogenesis is involved in both physiological and pathological processes. The angiogenic process is controlled by two major proangiogenic factors: VEGF and MMPs. The stimulatory effect of hypoxia involves the stabilization of hypoxia-inducible factor (HIF)-1, resulting in an increased expression of VEGF. This stimulates the formation of new blood vessels. 50,51
MMPs are a family of zinc endopeptidases capable to degrade components of the extracellular matrix. Once activated the MMPs are a subject of inhibition by TIMPs. 52
Our experiment showed that the VEGF level increased significantly after 24 h of hypoxia. The level was also significantly higher comparatively with controls after 5 days of hypobaric hypoxia although lower than after the 24-h interval. However, the level in the 24-h group treated only with vehicle remained significantly higher than in the 5-day group, suggesting that 24 h of hypobaric hypoxia produced a long-standing stimulatory effect on VEGF expression. In a study investigating the time course of the expression of brain VEGF mRNA and protein levels in mice maintained for a prolonged period in hypoxia, a maximum increase of VEGF mRNA and VEGF protein were obtained at 2 days of hypoxia. Increases in protein levels lasted for a longer period. It was suggested that VEGF expression is regulated by tissue metabolic balance rather than by oxygen partial pressure. 53
The dynamics of the MMP2 level in this experiment showed that MMP2 expression was slightly increased only in the group treated with vehicle after 24 h of hypoxia, demonstrating that this proteinase was activated in a longer interval from the returning animals to normal oxygen pressure comparatively with VEGF. In the same time, TIMP1 levels were similar to controls.
GSE treatment reduced VEGF brain concentrations, especially in animals which were kept for 5 days in a hypobaric chamber. MMP2 expression fell back to those in controls in both the treated groups, while polyphenolic treatment slightly increased TIMP1 to exert its inhibitory function on MMP.
Studies in vitro and in vivo demonstrated that polyphenols from red wine and green tea have potent antiangiogenic properties by preventing the expression of proangiogenic factors like VEGF most likely due to their antioxidant properties. 54 –56 It was demonstrated also that GSE inhibits VEGF expression by blocking HIF-1α protein synthesis through inhibiting the PI3/Akt signaling pathway. 57 Polyphenols were also demonstrated to inhibit expression and activity of MMP2 in different cell types. 55,58 On the other hand, in a study regarding postischemic neovascularization, it was demonstrated that red wine polyphenols have a dual effect as a function of dose: they proved to be proangiogenic at a low dose and antiangiogenic at a higher dose. 59 The results obtained in this study show that three doses of our grape seed extract have an antiangiogenic effect acting on the biomolecules involved in the angiogenesis process.
In conclusions, hypobaric hypoxia determines a strong oxidative stress in the animal brain evidenced by significantly increased levels of reactive oxygen and nitrogen species. In the same time, hypoxia produced increased levels of IL-6 cytokine as well as of VEGF and MMP, which are factors involved in the angiogenic process. The brain concentration of these parameters and their dynamics depend on the length of exposure to hypobaric hypoxia.
Three doses of GSE administered after hypobaric hypoxia produced a significant reduction of brain ROS and NO levels and acted as an anti-inflammatory agent by reducing IL-6 cytokine concentrations. GSE demonstrated antiangiogenic properties evidenced by decreased VEGF and MMP2 and slightly increased TIMP1 brain levels. GSE might possess beneficial effects in protecting the brain against deleterious effects of hypobaric hypoxia.
Footnotes
Acknowledgment
Authors acknowledge technical support for animal treatment from veterinary technician Remus Moldovan.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.