
Abstract
Polyphenols, including green tea catechins, are secondary plant compounds often discussed in the context of health-promoting potential. Evidence for such effects is mainly derived from epidemiological and cell culture studies. The aim of the present study was to investigate antidiabetic, antiadipogenic, and anti-inflammatory effects at nonpharmacological doses in an obese diabetic mouse model that exerts early relevant clinical signs of non–insulin-dependent diabetes mellitus. Female db/db mice received a flavonoid-poor diet either without additive, with rosiglitazone (RSG, 0.02 g/kg diet), or with green tea extract (low-dose green tea extract [LGTE] and high-dose green tea extract [HGTE], 0.1 and 1 g/kg diet). Food and water were freely available. The body weight was monitored weekly. Blood was sampled (12-h fasted) from the tail vein on day 28 and analyzed for glucose, cholesterol, triacylglycerol, nonesterified fatty acids, insulin, adiponectin, and soluble intercellular adhesion molecule-1 (sICAM-1). Blood glucose was also analyzed on day 14. Furthermore, sICAM-1 release was investigated in tumor necrosis factor alpha-stimulated EAhy926 cells. After 14 days, fasting glycemia was improved by RSG or HGTE supplementation compared to controls. However, at the end of the study (day 28), only RSG exhibited glucose-lowering effects and induced plasma adiponectin concentrations, paralleled by higher body weight gain and reduced periuterine fat pads compared to controls. However, only GTE treatment reduced sICAM-1 release in vitro and in vivo. Nonpharmacological HGTE supplementation in db/db mice caused (1) no adiponectin-inducing or antiadipogenic effects, (2) reduced sICAM-1 release, thereby potentially exerting anti-inflammatory effects in the progressive diabetic state, and (3) a transient improvement in glycemia.
Introduction
N

Plasma glucose concentrations of db/db mice in the 12-h fasted state after 14 and 28 days of feeding either without additive (control [C]) or with the addition of rosiglitazone (RSG, 0.02 g/kg diet) or green tea extract (0.1 or 1 g/kg diet, low-dose green tea extract [LGTE] and high-dose green tea extract [HGTE]). Data are means+standard error of the mean (SEM; n=8), two-way analysis of variance (ANOVA), *P<.05 compared to controls.
Materials And Methods
Animals and diets
Female db/db mice (BKS.Cg-m+/+Lepr<db>/J; n=32), initial body weight 30.1±0.5 g (Charles River Laboratories, Brussels, Belgium), were housed in cages with sawdust-covered solid flooring in a controlled environment (22°C±2°C, humidity 65%) with a 12-h light/dark cycle. Animals had free access to food and tap water. During the first week, all animals were fed a flavonoid-poor diet (C1000 control diet; Altromin GmbH, Lage, Germany; Table 1) without additives; thereafter, animals were randomly assigned to one of four groups and received the same flavonoid-poor diet either without additive (C), with the addition of rosiglitazone (RSG, 0.02 g/kg diet; Avandia®, GlaxoSmithKline, Munich, Germany) or green tea extract (0.1 or 1 g/kg diet, low-dose green tea extract [LGTE] and high-dose green tea extract [HGTE]; Polyphenon 60, Sigma-Aldrich Chemie GmbH, Steinheim, Germany, Table 2) for 28 days. Diets were prepared daily, and supplement-added, just before, feed was offered (daily at the beginning of the dark phase after removal of feed residues). The body weight was monitored weekly. Blood was collected from the tail vein for fasting (12 h) glucose measurements (HemoCue® System; HemoCue GmbH, Grossostheim, Germany) on days 14 and 28. After the 28-day feeding period, at the end of the dark phase and the 12-h fasting phase, blood from the tail vein was also collected in EDTA tubes (2.6 mL; Sarstedt AG & Co., Nürnbrecht, Germany) for plasma preparations. Thereafter, animals were sacrificed under anesthesia (CO2) by decapitation, and all tissues and organs were immediately dissected and snap-frozen in liquid nitrogen for storage at −86°C. Fresh weight of the liver and periuterine adipose tissue was determined before freezing. Animal experiments were approved by the Ministry of Agriculture, the Environment, and Rural Areas of Land Schleswig-Holstein, Germany (No. V 312-72241.121-25).
Information as provided by the manufacturer (Altromin GmbH).
Information as provided by the manufacturer (Sigma-Aldrich Chemie GmbH).
Determination of plasma metabolites and hormones
Plasma triacylglycerols (TG), nonesterified fatty acids (NEFA), and total cholesterol were quantified spectrophotometrically (Konelab 20i; Konelab Corporation, Espoo, Finland) using commercially available kits (TG and cholesterol: Thermo Fisher Scientific GmbH, Dreieich, Germany; NEFA: Wako Chemicals, Neuss, Germany). Plasma concentrations of insulin, adiponectin, and soluble intercellular adhesion molecule-1 (sICAM-1) were measured by enzyme-linked immunosorbent assays (ELISA) using commercial kits (insulin: ELISA Kit; DRG Diagnostics, Marburg, Germany; adiponectin: ELISA Quantikine® mouse Adiponectin/Acrp30; R&D Systems GmbH, Wiesbaden, Germany; sICAM-1: Quantikine® mouse ELISA sICAM-1; R&D Systems GmbH).
Cell culture
EAhy926 endothelial-like cells were kindly provided by Dr. Edgell (University of North Carolina, Chapel Hill, NC, USA) and cultured in the Dulbecco's modified Eagle's medium containing glucose (4 g/L), supplemented with
sICAM-1 release in EAhy926 cells
For determination of sICAM-1 in TNF-α-stimulated endothelial cells, 200,000 EAhy926 cells per well were seeded in a 12-well plate. After 24-h cultivation, the culture medium was replaced with a medium containing GTE (10 μg/mL), EGCG (25 μmol/L), or RSG (1 μmol/L; Biozol Diagnostics, Eching, Germany). This RSG concentration was proven to activate PPAR-γ in a reporter gene assay using dimethyl sulfoxide (0.1%) as a negative control. We confirmed that these GTE, EGCG, and RSG concentrations had no adverse effects on cell viability (determined by neutral-red assay). After 24 h of incubation, the medium was aspirated, and the cells were washed once with prewarmed PBS to completely remove test substances. Subsequently, PBS was replaced, and cells were stimulated for 24 h with 1 ng/mL human TNF-α (Invitrogen GmbH, Karlsruhe, Germany). Thereafter, supernatants were collected and aliquots stored at −80°C for sICAM-1 analysis. Supernatant sICAM-1 concentrations were assessed by commercial sandwich ELISA (Quantikine Human ELISA sICAM-1/CD54; R&D Systems GmbH). The undiluted samples were applied to a 96-well plate with immobilized monoclonal antibodies against human sICAM-1. ELISA was performed according to the manufacturer's protocol. For the determination of sICAM-1, three independent experiments in triplicate were carried out.
Statistical analysis
Results are reported as means with the standard error of the mean. Statistical analysis was performed using GraphPad Prism (GraphPad Software, Inc., version 4.01, 2004; San Diego, CA, USA). The group means were compared by one-way or two-way analysis of variance (ANOVA) with subsequent Bonferroni's multiple comparison test (dietary treatment against control) or Bonferroni post-tests comparing replicate means, respectively. Significance was set at P<.05.
Results
Plasma parameters
Feeding db/db mice for 14 days with RSG or HGTE resulted in reduced fasting plasma glucose concentrations compared to controls (Fig. 1). After 28 days, however, the blood glucose-lowering effect was only observed in the RSG group (Fig. 1, Table 3), whereas a reduced plasma sICAM concentration was found only in the HGTE group (Table 3). After 28 days, no differences were found in fasting plasma TG, NEFA, cholesterol, or insulin levels between the RSG or L/HGTE groups and controls (Table 3). In RSG-fed mice, lower fasting plasma glucose concentrations were paralleled by elevated plasma adiponectin concentrations compared to controls (Table 3). A two-way ANOVA indicated that effects on fasting plasma glucose concentrations are mainly attributable to the dietary treatment (32.44%, F=17.13, P<.0001) and the duration of dietary treatment (26.43%, F=41.71, P<.0001).
Reported as mg liver tissue/g body weight.
Data are means±standard error of the mean (n=8), one-way analysis of variance, * P<.05 compared to controls.
ACRP30, full-weight adiponectin; C, control; CHOL, total cholesterol; HGTE, high-dose green tea extract (1 g/kg diet); LGTE, low-dose green tea extract (0.1 g/kg diet); NEFA, nonesterified fatty acids; RSG, rosiglitazone (0.02 g/kg diet); sICAM, soluble intercellular adhesion molecule-1; TG, triacylglycerols.
Body weight and liver and periuterine fat mass
All animals continuously gained body mass during the feeding period (Fig. 2). However, at the end of the feeding period (days 21 and 28), the average body weight was higher in the RSG-fed group compared to controls (Fig. 2A). Conversely, body weights of L/HGTE-treated db/db mice did not differ compared to controls (Fig. 2). The mean liver wet weight was lowered by HGTE feeding (Table 3), while periuterine fat mass was reduced by ∼25% by RSG treatment compared to controls, despite these mice having the highest body weights (Fig. 2A, B).
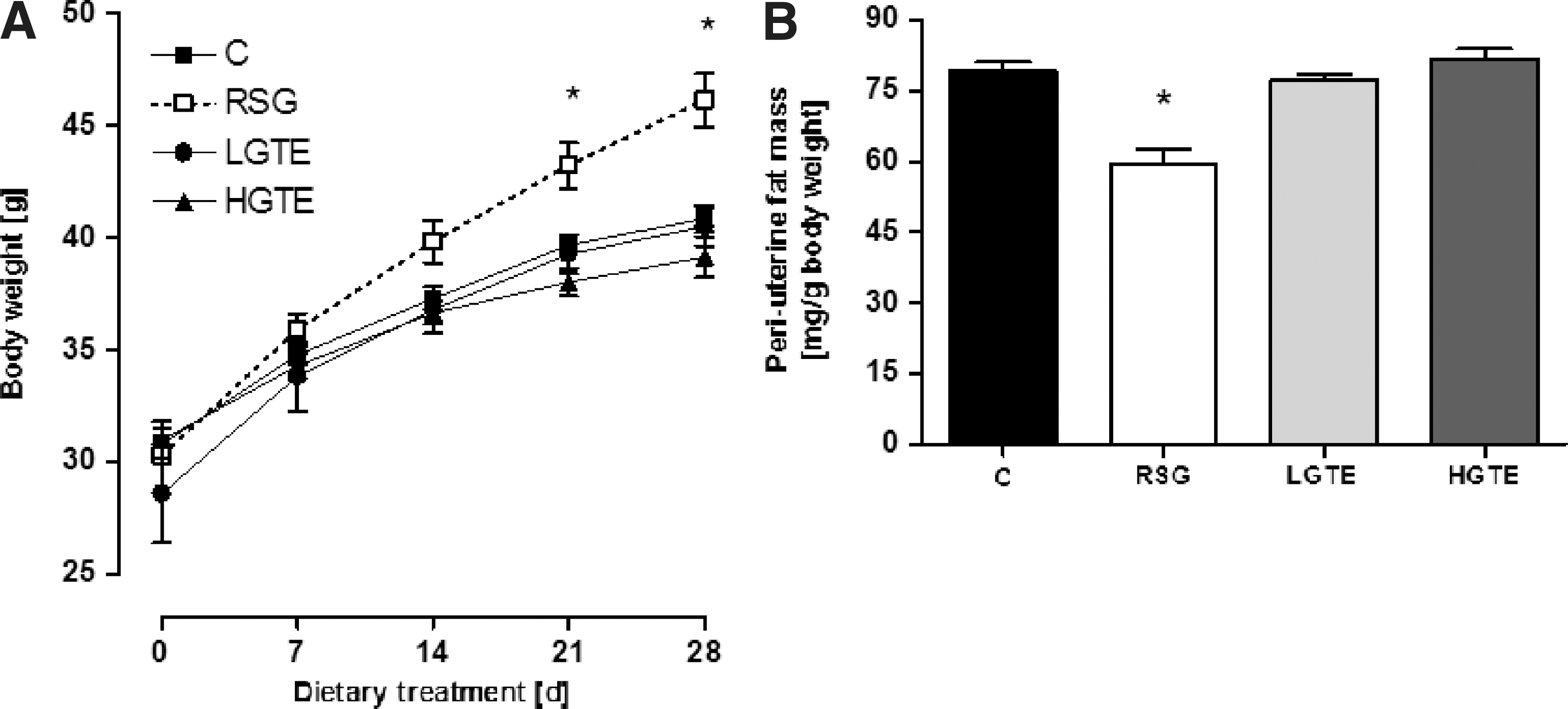
Body weight development of db/db mice over 28 days
sICAM-1 release in stimulated EAhy926 endothelial-like cells
No sICAM-1 was detected in the supernatants of unstimulated EAhy926 endothelial cells. However, stimulation of EAhy926 cells with TNF-α markedly increased the release of sICAM-1. While RSG had no effect on sICAM-1 release in stimulated EAhy926 cells, GTE and EGCG reduced the release of sICAM-1 in stimulated EAhy926 cells (Fig. 3).
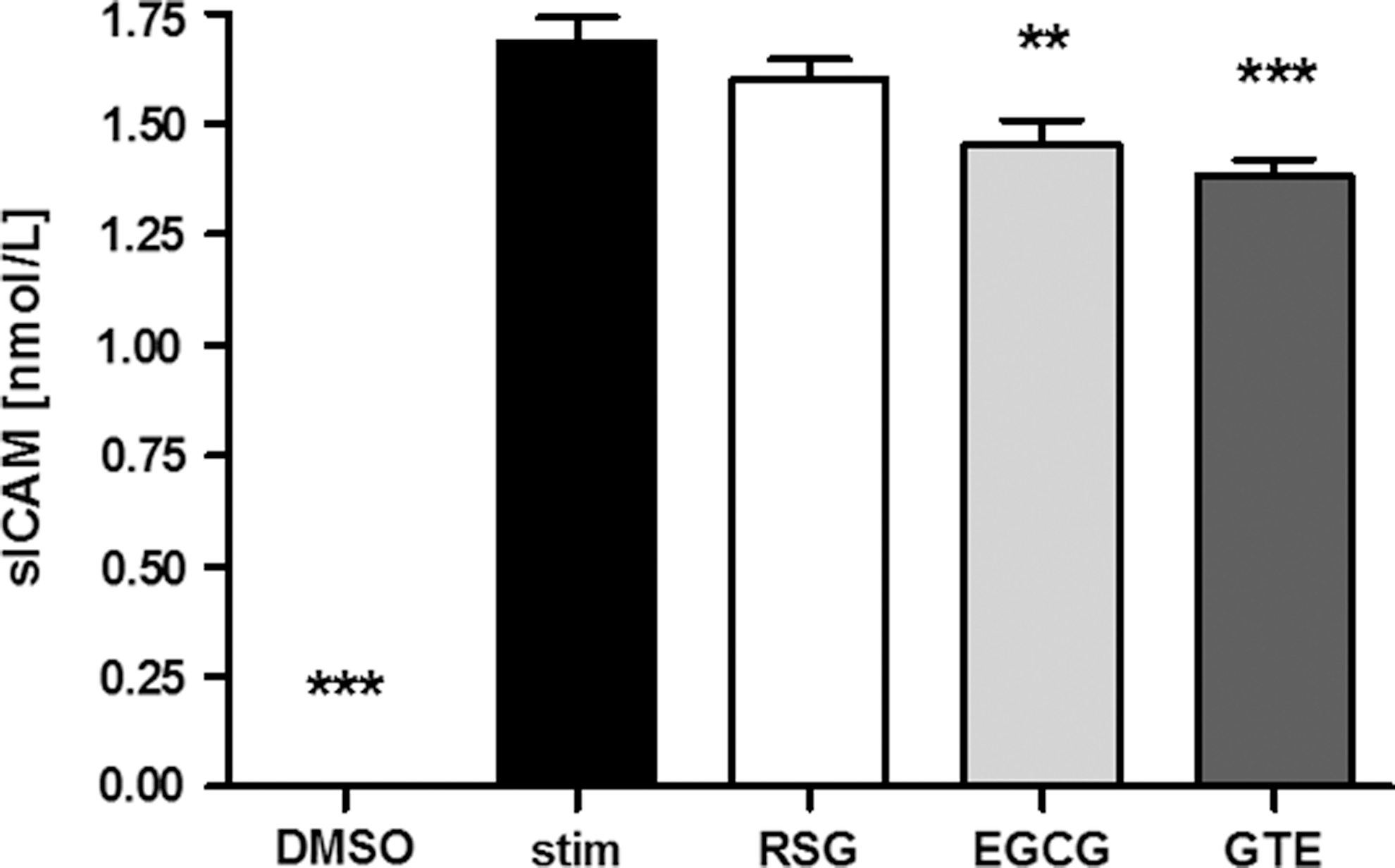
Effect of green tea extract on soluble intercellular adhesion molecule 1 (sICAM-1) release in tumor necrosis factor alpha (TNF-α)-stimulated EAhy926 cells. EAhy926 cells were preincubated (24 h) with dimethylsulfoxide (DMSO, 0.1%) as a negative control, RSG (1 μmol/L), green tea extract (GTE, 10 μg/mL), or epigallocatechin-3-gallate (EGCG, 25 μmol/L) after 24-h stimulation with TNF-α (1 ng/mL). Data are means+SEM (n=9), one-way ANOVA. **P<.01, ***P<.001 compared to stimulated control (stim).
Discussion
Although beneficial health effects on obesity, NIDDM, and/or coronary heart diseases due to the consumption of flavanol-rich green tea or oral EGCG supplementation have been suggested, 10,20 –22 the underlying mechanisms remain largely elusive. Thus, we investigated the glucose-lowering, antiadipogenic, and anti-inflammatory effects of dietary GTE in an animal model used extensively for the screening of antidiabetic agents. 23 The obese db/db mouse reliably simulates the development of insulin resistance, demonstrating progressive beta-cell failure with pronounced hyperglycemia, adiposity, and hypoadiponectinemia similar to that found in human NIDDM. In the present study, diet was supplemented with GTE (containing 34% EGCG; 0.1 or 1 g GTE/kg diet). Corresponding daily intake was ∼6 mg EGCG/kg body weight (LGTE) or ∼60 mg EGCG/kg body weight (HGTE), respectively. The lower dose was chosen to simulate an EGCG intake achievable by fortified daily green tea (nutrient data bank [NDB] No. 99070) consumption (0.5 L), 24 but was explicitly nonpharmacological. After 14 days of dietary intervention, body weights of mice did not differ. Groups supplemented with RSG or the high dose of GTE had improved fasting glycemia compared to controls, while fasting plasma glucose concentrations were not reduced in mice fed the low GTE dose. Thus, it seems that even in the early stages of insulin resistance, low doses of GTE do not exhibit glucose-lowering effects in female db/db mice. Continuous feeding of the respective diets resulted in higher body mass gain in mice fed RSG compared to controls, while mice fed GTE did not differ from controls. However, as insulin resistance advances, improved fasting glycemia, paralleled by unreduced plasma adiponectin, was only found in mice fed RSG after the 28-day feeding period. Neither of the two dietary GTE doses exhibited glucose-lowering effects in female db/db mice after 28 days. Thus, we assume that administration of nonpharmacological EGCG doses only transiently lowers plasma glucose as a consequence of progressive pancreatic β-cell failure in this animal model of obesity and diabetes. To date, neither epidemiological nor interventional studies provide clear-cut evidence for glucose-lowering effects of EGCG consumption in humans. 25 –28 Also, antiadipogenic effects of EGCG, reported in previous in vitro investigations, 15.29 –31 were not observed in this study. In the present study, animals were housed in groups of four with free access to food and water, and thus individual food intake was not assessed. Additionally, animals were fed a powdery diet resulting in extensive food wastage, and thus accurate measurement of a group-based feed consumption was not possible. However, the impact of RSG on feed intake and body weight gain has been investigated before. Despite reduced feed intake, RSG intake was paralleled by an increase in body weight, a known critical side effect of RSG intake. 32 Nevertheless, the impact of EGCG on body weight remains controversial. Intraperitoneal injection of EGCG has been found to have dose-dependent antiobesity effects in Sprague-Dawley rats and in lean and obese Zucker rats. 33 In the same study, oral EGCG supplementation reduced food intake by 50%–60%, causing a resultant reduction in weight gain. 33 Oral EGCG supplementation (10 g/kg diet) prevented diet-induced increases in body weight in C57BL/6J mice treated for 5 months, and reversed established obesity in Sprague-Dawley rats treated for 4 weeks. 15 However, 10 g EGCG/kg diet did not reduce weight gain in db/db mice unless fed for 9 weeks. Thus, it can be concluded that antiadipogenic effects of EGCG are mainly dose-, but also time and model dependent, and are not established for EGCG intake achievable by even fortified daily green tea consumption.
Hypoadiponectinemia is associated with adiposity and impaired insulin sensitivity. 34 Adiponectin synthesis is stimulated upon ligand binding to the transcription factor PPAR-γ, which is the major target of thiazolidinones. 3 In the present study, hypoadiponectinemia was prevented, irrespective of higher weight gain, only in the RSG-supplemented group, suggesting that GTE has no PPAR-γ ligand activity in db/db mice at the doses employed herein.
Inflammation is a feature common in both obesity and insulin resistance, often accompanied by elevated cytokine secretion from adipose tissue. In the literature, anti-inflammatory effects of RSG are discussed controversially. Cytokines, such as TNF-α and nuclear factor kappa-light-chain enhancer of activated B-cell (NF-κB), induce the expression and release of adhesion molecules (ICAM-1) in different cell types. 35 In chronic inflammatory diseases, sICAM-1 occurs in the circulation and can be used as a biomarker of inflammation. 36 On one hand, RSG treatment has been shown to reduce cytokine expression, 37 but on the other hand, it fails to reduce TNF-α mRNA concentrations in white adipose tissue of db/db mice. 38,39 EGCG, however, has previously been reported to inhibit TNF-α secretion and thus NF-κB function in several cell lines. 40 –42 In the present study, only GTE and EGCG reduced sICAM-1 release in stimulated EAhy926 cells, and only high-dose GTE reduced sICAM release in db/db mice. This might contribute to a hepatoprotective effect. During chronic low-grade inflammation, numerous mediators are released from immune cells and adipocytes that contribute to liver damage and liver disease, associated with hepatic steatosis, necroinflammation, apoptosis, and fibrosis. 43 Despite having no improvement in plasma triacylgylcerol, NEFA, and cholesterol concentrations, mice fed HGTE displayed decreased liver wet weights compared to controls. Although we have not measured hepatic TG content in mice, liver wet weight together with reduced plasma sICAM concentrations is be suggestive of some hepatoprotective effects of GTE.
In summary, oral green tea extract supplementation at nonpharmacological EGCG doses had no adiponectin-inducing or antiadipogenic effects in db/db mice. However, a higher dose of green tea extract, and therefore EGCG, transiently improved glycemia and effectively reduced sICAM-1 release, and thus might exert anti-inflammatory and hepatoprotective effects in the progressive diabetic state.
Footnotes
Acknowledgment
We are grateful to Wiebke Kühl (Institute of Animal Nutrition & Physiology, Christian Albrechts University of Kiel, Germany) for excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist