
Abstract
Numerous honeybee products are used in medicine, but the literature furnishes no information concerning the effects of the drone milk (DM), although drone brood, which is similar to DM, was reported to elicit a hormone-like strengthening effect. In certain countries, DM is traditionally used to treat infertility and to promote vitality in both men and women. The aim of this study was to determine the putative estrogen hormone–like effect of raw DM in rats and to identify the effective compounds. Uterotrophic assays revealed that DM increased the relative weight of the immature rat uterus. This effect was confirmed by reverse transcription polymerase chain-reaction and Western blot methods, in which the mRNA and protein expression of the estrogen-dependent peptide complement component C3 was determined. Column chromatography and uterotrophic assays were used to fractionate and check bioactivity, respectively. The active compound after the last fractionation was identified by the nuclear magnetic resonance and mass spectrometry techniques as E-dec-2-enedioic acid, which is very similar to the fatty acids with estrogenic activity that were previously isolated from royal jelly. These results lead us to suppose that E-dec-2-enedioic acid is responsible for the estrogen-like effect of DM. This appears to be the first report on the pharmacological effects of DM and E-dec-2-enedioic acid in mammals.
Introduction
P
Plant sterols are important materials for insects to synthesize their own hormones. From phytosterols, honeybees are able to produce ecdysteroids that regulate molting, metamorphosis, and reproduction. These ecdysteroids are to be found in different organs of the insects and are also synthesized by the honeybee queen ovaries. 5,6 Ecdysteroids such as 20-hydroxyecdysone have been proven to elicit anabolic activity. 7 Additionally, several fatty acids from royal jelly (RJ), including 10-hydroxy-trans-2-decenoic acid and its derivatives, have been found to stimulate the estrogen receptors and to induce hypertrophy of the myometrial epithelium in the rat. 8 RJ has become a natural alternative to hormone replacement therapy in postmenopausal women. 9
It is not widely known that, similarly to the queen, drone honeybees have their own special food. In a similar manner to RJ, drone milk (DM) is secreted by the hypopharyngeal and mandibular glands of worker honeybees (Apis mellifera L.). DM is the main component of drone brood, which also contains larvae and pupae of drones in the comb. DM is separated from drone brood by extraction to eliminate the larvae and pupae during the harvest. Drone brood is traditionally used in China, Russia, and Transylvania (Romania) for climacterium, sexual problems, and rehabilitation and activation of the elderly. The main components of drone brood are proteins, lipids, fatty acids, carbohydrates, and water, and it contains vitamins and minerals, too. 10 DM has a similar use, but without available documentation.
Although RJ and DM are secreted by the same glands of worker honeybees, the latter has not been proven to show hormone-like activity as of yet. Our aim was to investigate the possible estrogenic effect of raw DM and to identify its active compounds by bioactivity-guided fractionation.
Materials and Methods
Collection of DM
DM was prepared during rape blooming and harvested by separation from the drone larvae and pupae in the late spring (first half of May) by a Hungarian private beekeeper, Sándor Polgár, in the acacia season. The raw liquid material was divided into plastic tubes and was stored at −20°C until the beginning of the investigation.
Uterotrophic assays
Animal investigations were carried out with the approval of the Hungarian Ethics Committee for Animal Research (permission number: IV/01758-0/2008). Female Sprague-Dawley rats were obtained from Charles River Laboratories, Budapest, Hungary.
Studies were performed in accordance with the protocol described by Kanno et al. 11 and Song et al. 12 Immature female Sprague-Dawley rats (20–21 days old) were randomly assigned to one of six groups (n=10) and were treated once daily for four consecutive days with the test compounds. No treatment-related clinical signs or abnormal changes were observed in any of the groups. The DM was diluted with distilled water, while the soy extract (Aktiv Meno Capsules, Drogerie Markt, Karlsruhe, Germany) was suspended in 0.5% mucilage methylcellulose (MM; Sigma Aldrich, Budapest, Hungary). Each soy isoflavone capsule contained 25 mg of a powder comprising soy isoflavone, several vitamins (C, E, biotin, and folic acid), and minerals (dicalcium phosphate and zinc oxide). Our preliminary study showed that in the range of 0.05–5 g DM/kg body weight (b.w.) per day, a daily dose of 0.5 g/kg b.w. was the most effective. A submaximal dose for uterotrophic effect of 0.5 g soy isoflavones/kg b.w. per day was previously calculated by de Lima Toccafondo Vieira et al. 4
DM (0.5 g/kg per day) and the soy extract (soy isoflavone, 0.5 g/kg per day) were administered by oral gavage. The positive control 17β-estradiol valerate dissolved in olive oil (3.9 μg/kg b.w. per day, equivalent to 3.0 μg 17β-estradiol/kg b.w. per day; Sigma Aldrich) and the estrogenic antagonist ICI 182.780 (400 μg/kg b.w. per day; Sigma Aldrich) were administered by subcutaneous injection. The control group was treated orally with distilled water. The administration protocol is outlined in Table 1. Table 2 shows the administration protocol of the bioactivity-guided fractions. The control groups were treated with either 0.5% MM or olive oil.
DM, drone milk; p.o., orally; b.w., body weight; s.c., subcutaneously.
MM, mucilage methylcellulose.
One day after the final treatment, the rats were sacrificed by exsanguination under deep isoflurane (Abbott Laboratories, Abbott Park, IL, USA) anesthesia. The uteri were removed, trimmed free of fat and adjacent tissue, and weighed on an analytical scale (HR-202; A&D Instruments, Oxfordshire, United Kingdom). Organ weights were expressed as the relative organ weight (ROW).
Determination of complement component C3 mRNA by real-time reverse transcription polymerase chain-reaction
Mature female rats (160–180 g) were ovariectomized under isoflurane anesthesia 8 days before treatment. The animals were randomly grouped and were treated once daily for 2 days with 17β-estradiol valerate (96.2 μg/kg b.w.), DM (0.5 g/kg b.w.), or vehicle only. One day after the final treatment, the uteri were removed. The uterus samples were stored at 4°C overnight in an RNAlater solution (Life Technologies, Budapest, Hungary); the supernatant was then removed, and the samples were stored at −70°C.
Total RNA was isolated from the uterus tissue by using the TRI Reagent (Molecular Research Center, Inc., Cincinnati, OH, USA). The quality and the quantity of RNA were assessed via the ratio of the absorbances at 260 and 280 nm; all samples exhibited ratios in the range 1.6–2.0. With 1 μg total RNA, cDNA was first synthesized by reverse transcription (RT) using the High-capacity RNA-to-cDNA Kit (Life Technologies). The samples were subjected to 37°C for 60 min, 95°C for 5 min, and 4°C for 5 min. The cDNA was stored at −20°C. The polymerase chain-reaction (PCR) products (C3 Rn00566466_m1) and GAPDH primer (Rn99999916_s1) as an endogenous control were amplified with the TaqMan Gene Expression Master Mix (Life Technologies) and the ABI StepOne Real-Time cycler. For amplification, the following conditions were used: 50°C for 2 min, 95°C for 10 min, and 40 cycles of 95°C for 15 sec and 60°C for 1 min. The fluorescence intensities of the probes were plotted against the PCR cycle numbers. The amplification cycle displaying the first significant increase in the fluorescence signal was defined as the threshold cycle.
Western blot analysis
Mature female rats (160–180 g) were ovariectomized and treated as described previously. Uterus tissues were powdered with Sartorius Mikro Dismembrator U (Sartorius, Göttingen, Germany) and homogenized in a RIPA lysis buffer combined with PMSF solution, sodium orthovanadate solution, and protease inhibitor cocktail solution (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The samples were centrifuged at 10,000 g for 10 min, and the supernatant was then used to measure the total protein. About 20 μg of protein per well was subjected to electrophoresis on 4–12% NuPAGE Bis–Tris Gel in XCell SureLock Mini-Cell Units (Life Technologies). Proteins were transferred from gels to nitrocellulose membranes using the iBlot Gel Transfer System (Life Technologies). Antibody binding was detected with the WesternBreeze Chromogenic Western Blot Immunodetection Kit (Life Technologies). The blots were incubated on a shaker with C3 and β-actin polyclonal antibody (Santa Cruz Biotechnology; 1:200) in the blocking buffer. The optical density of each immunoreactive band was determined with Kodak 1D Image analysis software (Carestream Health, Inc., Rochester, NY, USA). Optical densities were calculated as arbitrary units after a local area background subtraction.
Fractionation of DM
The crude DM was diluted with water (165 mL) and extracted with petroleum ether (5×200 mL). The petroleum ether phases were collected and re-extracted with water (3×10 mL). The combined aqueous phase was applied to an octadecyl-silica column (Chemie Uetikon-C-Gel, Chemie Uetikon, Uetikon am See, Switzerland; 0.06–0.02 μm) and subjected to low-pressure reversed-phase (RP) column chromatography (CC). Water, 20% (I/A), 40–60–80% (I/B) aqueous MeOH (700 mL each), MeOH (I/C; 1000 mL), and dichloromethane (I/D; 1000 mL) were used for gradient elution.
The fractions eluted from the reversed-phase column with 20%, 40%, 60%, and 80% of aqueous MeOH were combined and evaporated to dryness. The residue (2.86 g) was further fractionated through repeated low-pressure RP CC. Elution was carried out with different mixtures of aqueous MeOH, using stepwise gradient elution with 5% or 10% increases of the MeOH content in each step up to 80% and finally with MeOH.
The fractions eluted with 20% (II/A), 35% (II/B), 70% (II/C), or 75% (II/D) aqueous MeOH were further examined by the uterotrophic assay. The most active fraction (II/B) eluted with 35% aqueous MeOH was further purified on a preparative scale by RP (C18) thin-layer chromatography (TLC) using tetrahydrofuran–water 6:4 v/v (TLC System 3). The band at RF=0.41 was scraped off, and the substances were eluted from the sorbent with MeOH. The solution obtained was evaporated to dryness, and the residue (0.06 g) was analyzed by TLC, mass spectrometry [MS], and nuclear magnetic resonance (III/A) analyses.
The whole separation procedure was controlled by RP TLC (TLC Systems 1 and 2). RP TLC was performed on 20 cm×20 cm glass TLC plates coated with RP-18WF254 (E. Merck, Darmstadt, Germany) at room temperature by an ascending technique with a mobile phase: TLC System 1, acetone–water (95:5 v/v); TLC System 2, tetrahydrofuran–water (7:3 v/v); and TLC System 3, tetrahydrofuran–water (6:4 v/v, in the case of preparative-scale TLC). The compounds were detected by the use of a vanillin–sulfuric acid spray reagent.
High-resolution MS and NMR analyses
An LTQ FT Ultra (Thermo Fisher Scientific, Bremen, Germany) spectrometer was used for high-resolution (HR) MS analyses. Ionization was achieved with electrospray ionization (ESI), operated in the positive-ion mode. The ion-transfer capillary temperature was set at 280°C, and the capillary voltage for each measurement was 4.1 kV. In collision-induced dissociation (CID) experiments, helium was used as the collision gas, and normalized collision energy (expressed as a percentage), which is a measure of the amplitude of the resonance excitation radio frequency voltage applied to the endcaps of the linear ion trap, was used to bring about fragmentation. The sodium adduct of the peak molecular ion was fragmented by CID at a normalized collision energy of 50%. The sample was dissolved in MeOH. Data for HRMS were acquired and analyzed with Xcalibur software version 2.0 (Thermo Fisher Scientific).
NMR spectra were recorded in MeOH-d4 (Merck) in a Shigemi sample tube at 298 K, using a Varian 800 MHz NMR spectrometer equipped with an 1H[13C/15N] Triple Resonance 13C Enhanced Salt Tolerant Cold Probe operating at 800 MHz for 1H. Chemical shifts were referenced to the residual solvent resonances (δH=3.31 ppm and δC=49.15 ppm). Pulse sequences in all experiments (1H, 1H-Presat, GHSQCAD, GHMBCAD, and one- and two-dimensional zTOCSY) were taken from the VNMRJ-3.2 software library without any modification.
Statistical analysis
Prism 4.0 Software was used for all statistical analyses for biological determinations (uterotrophic assay, RT-PCR, and Western blot). All data were analyzed by one-way analysis of variance, followed by the Newman–Keuls test, and each value is reported as a mean±standard error of the mean (S.E.M.). Significance was accepted at P<.05.
Results
In vivo estrogenic activity of raw DM
The estrogenic activity of raw DM was examined by means of uterotrophic assays (Fig. 1). The adequacy of the model in our laboratory was proved by treatment with 17β-estradiol valerate (E2). E2 increased the ROW of the uteri (wet uterus weight in mg/100 g b.w.) from the control group value of 42.35±2.85 to 109.60±4.00 mg/100 g b.w. This activity was diminished by the antiestrogenic ICI 182.780 (ICI) to 37.25±2.41 mg/100 g b.w.
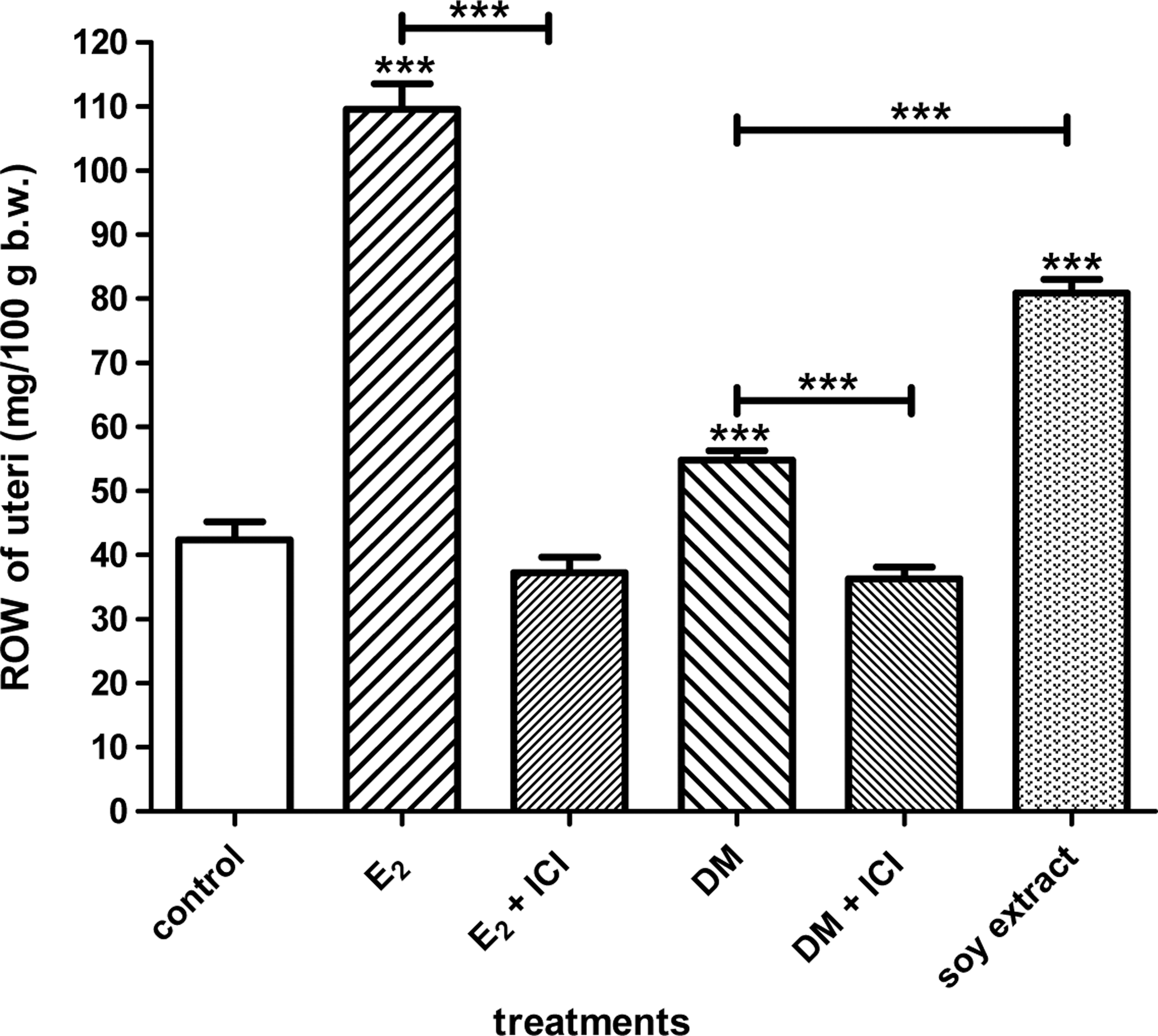
Changes in the relative organ weight (ROW) of the uteri in immature rats (n=10) after treatment with crude drone milk (DM), 17β-estradiol valerate (E2), or soy extract. DM, E2, and the soy extract increased the ROW of the uteri. These effects were ICI 182.780–sensitive. ***P<.001.
Treatment of the animals with raw DM increased the ROW to 54.84±1.41 mg/100 g b.w. Similarly to that of E2, the estrogenic effect of DM was blocked in the presence of ICI 182.780: the ROW was reduced to 36.30±1.77 mg/100 g b.w. The soy extract also increased the ROW (80.90±2.16 mg/100 g b.w.).
By means of a real-time PCR technique, we found that DM almost doubled the relative C3 mRNA expression in the uteri. The effect of positive control E2 was also significant (Fig. 2).
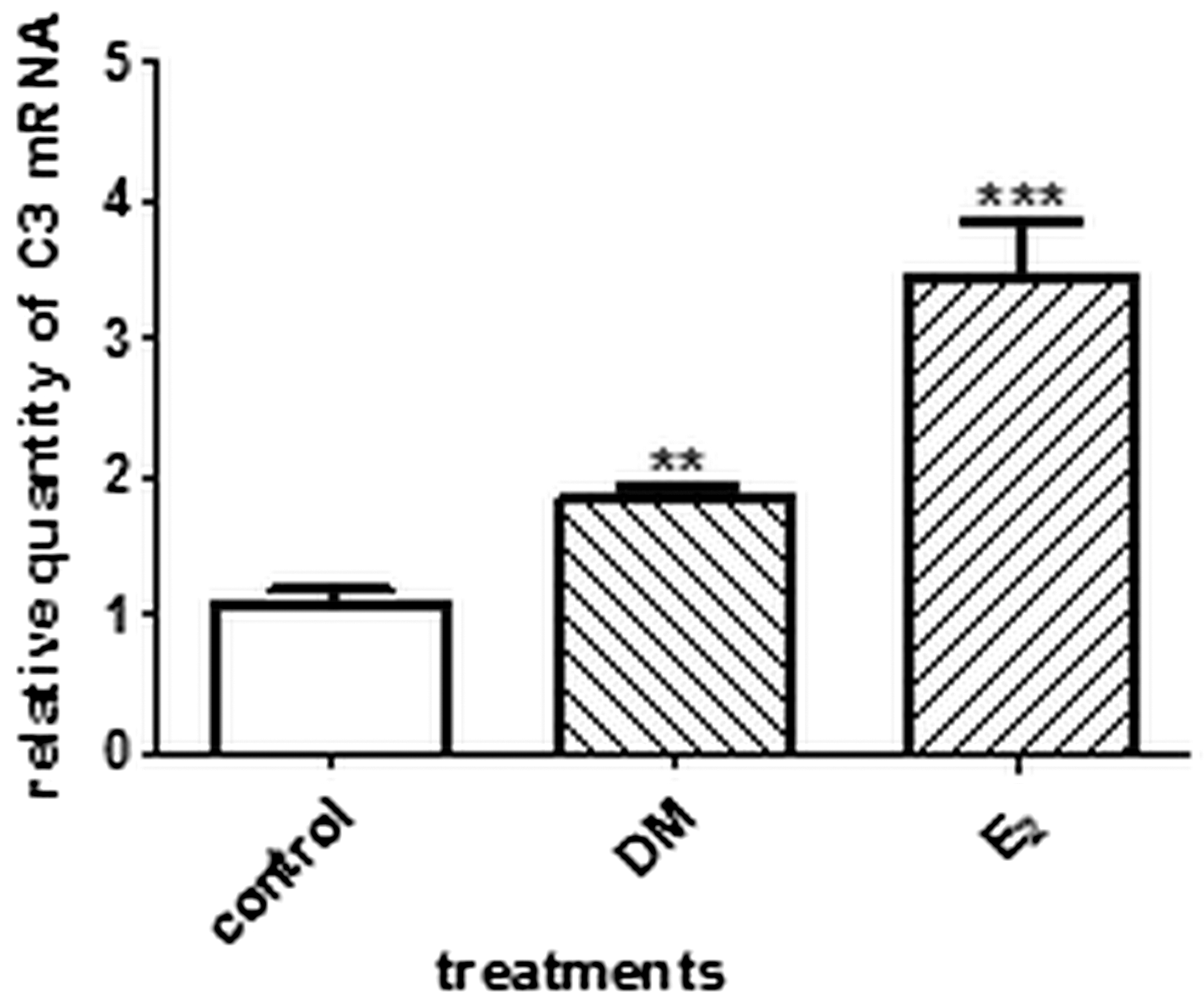
RT-PCR analysis of complement component C3 mRNA expression in the rat uteri (n=6). The crude DM increased the mRNA expression of the estrogen-dependent C3 in rat uterus tissue. **P<.01; ***P<.001.
In Western blot measurement, DM enhanced the level of C3 in the uterine tissue, and E2 was also effective (Fig. 3). The gel photo (Fig. 3B) reveals that the C3 protein is split into alpha- and beta-chains during the Western blot studies; the densities of both chains were therefore evaluated (Fig. 3A).

Western blot analysis of complement component C3 protein in rat uteri (n=6). The crude DM increased the expression of the estrogen-dependent C3 protein in rat uterus tissue
Bioactivity-guided fractionation of DM
The crude DM was diluted with water and extracted with petroleum ether to remove apolar contaminants. The prepurified extract was subjected to repeated RP CC. After the first separation, the fractions with the same compositions were combined and tested by uterotrophic assay (Fig. 4). The fractions were further separated with fraction I/C, but new fractions were ineffective in the uterotrophic assay; therefore, we did not continue the investigation of this product. On the basis of the results, the other two fractions (I/A and I/B) were chosen for further separation. As the main constituent of the more-active fraction (I/A) was also present in I/B, although the material content of the latter was much lower, these two fractions were combined and purified together by CC. The second CC separation, with the stationary phase in a high (e.g., 20-fold) excess, and elution of the constituents with a multistep gradient, proved to permit rather effective purification. The pharmacological experiments led to four fractions being subjected to further examination by the uterotrophic assay (Fig. 5). The most active fraction, II/B, was purified by preparative RP TLC. The fraction obtained (fraction III/A) was subjected to TLC, MS, and NMR analyses.
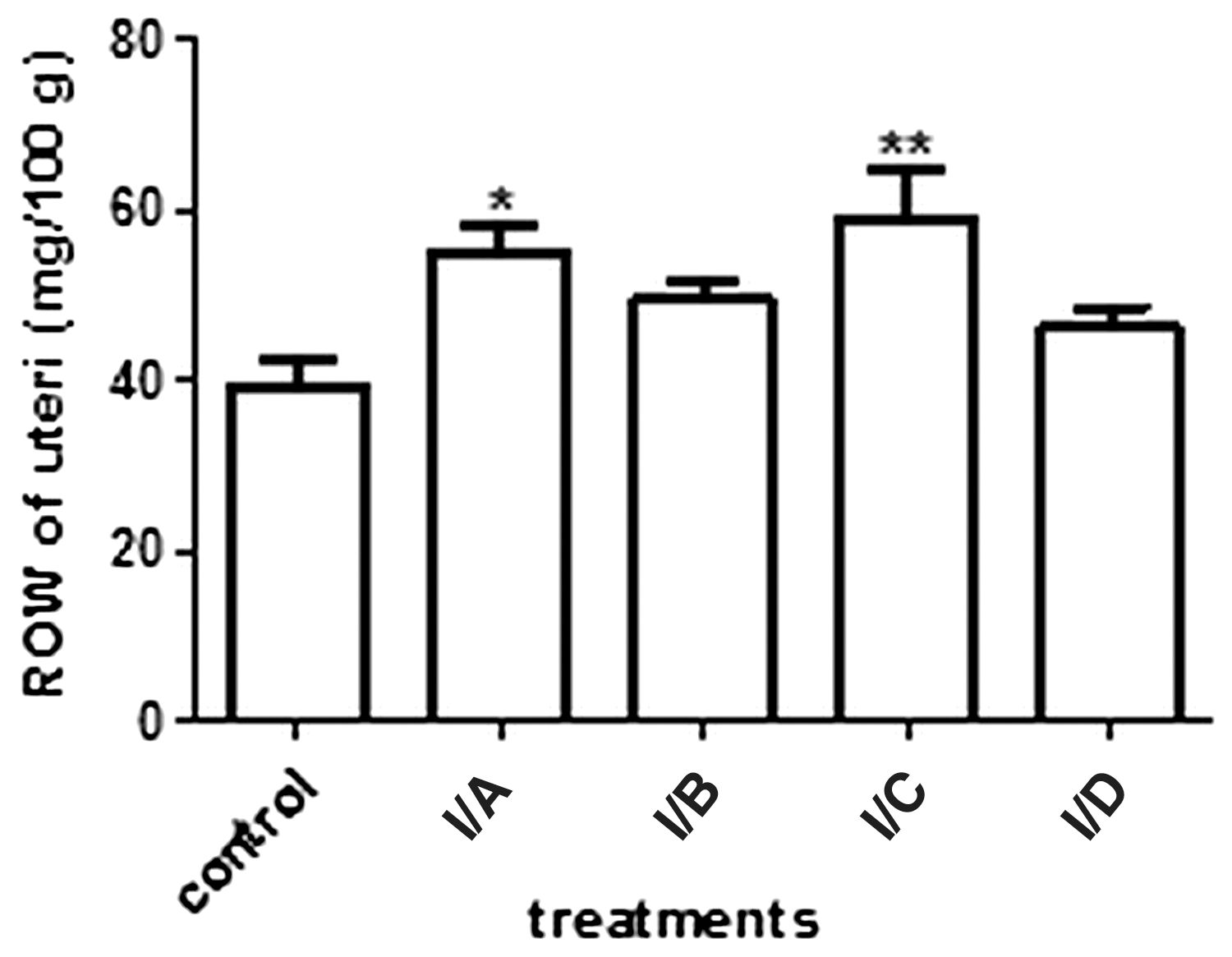
Bioactivity-guided fractionation 1. The 20% aqueous MeOH (I/A) and pure MeOH (I/C) fractions increased the ROW of the uteri (n=10). *P<.05; **P<.01.

Bioactivity-guided fractionation 2. Only fraction II/B increased the ROW of the uteri (n=10). From this fraction, the effective compound, E-dec-2-enedioic acid, was purified. ***P<.001.
This fraction was multicomponent, but one of the main components could be identified by ESI-HR-MS, 1H-Presat (Supplementary Fig. S1) and 2D-homo- (ZTOCSY) (Supplementary Fig. S2), and heteronuclear correlation (GHSQCAD and GHMBCAD) data. In the ESI-HR-MS spectrum, the sodium adduct of the molecular ion was detected at 223 m/z. The accurate mass value (223.09401) of this ion corresponded with an accuracy of 0.3 ppm to an elemental composition C10H16O4Na. Analysis of the 1H NMR spectrum indicated the presence of a trans- double bond, as evidenced by two doublets of triplets at 5.80 (H-2, J=15.5 and 1.5 Hz) and 6.92 ppm (H-3, J=15.5 and 6.9 Hz). The correlations of the methylene and olefinic protons observed in the one- and two-dimensional ZTOCSY spectra permitted the complete 1H NMR assignment of compound
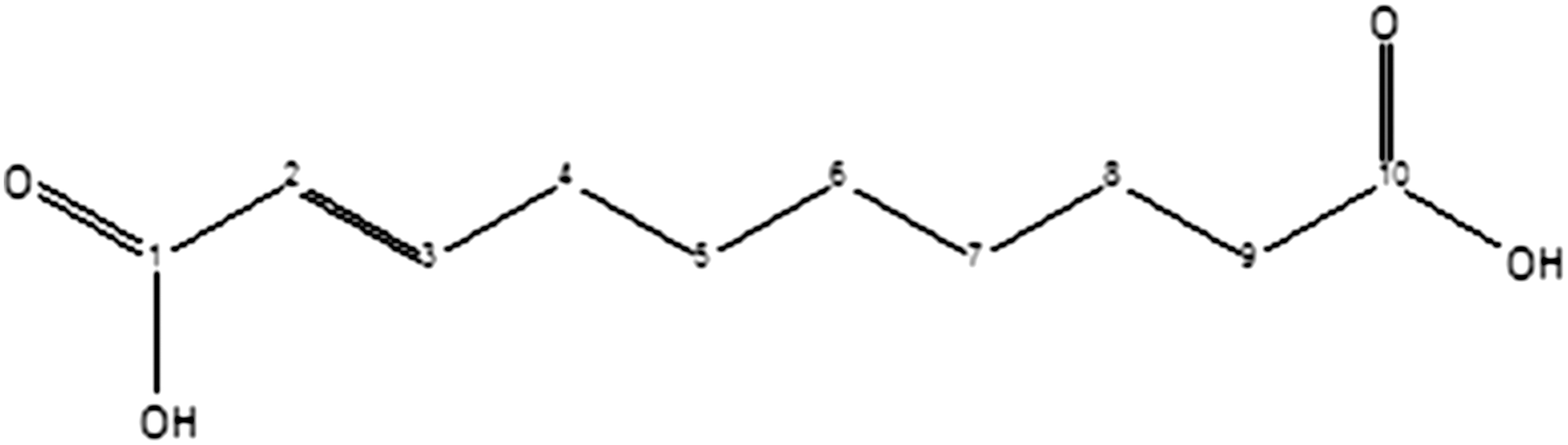
Structure and carbon atom numbering of E-dec-2-enedioic acid (structure
1H
Discussion
DM is a little-known purified extract of drone brood, without larvae and pupae. This natural material is a yellowish, milky, slightly sweet liquid that is an essential food for all forms of drones in the hive. Since drone brood is used to treat sexual problems in folk medicine, we hypothesized that DM may have some hormonal effects, too. Therefore, we searched for both the female hormonal effect and the active component of DM.
The raw DM was able to increase the ROW of the uteri in immature female rats, and this effect was estrogenic antagonist–sensitive, similar to the action of E2. This result clearly indicates that the raw DM has an estrogenic activity. Although the extent of the ROW-increasing effect of DM was moderate when compared with that of the soy extract or E2, its estrogenic activity was still noteworthy, as the DM used was an unpurified material.
Besides the uterotrophic assay, we sought molecular markers with which to confirm the estrogenic effect of DM. C3 expression has been reported to be a molecular marker of estrogenic activity. 13 The raise in the level of the mRNA and protein expression of C3 protein supports the results of the uterotrophic assay. The changes in the uterine level of C3 protein are in harmony with the data gained from an in vivo uterotrophic test.
To identify the active compounds in raw DM, bioactivity-guided fractionations were performed. Through a three-step procedure, we gained a fraction and identified the structure of one main compound. By HRMS and NMR data, the main component in fraction III/A is E-dec-2-enedioic acid, which is very similar to estrogenic fatty acids isolated from RJ. 8 This molecule was earlier found as a major mandibular gland component in the queen and worker dwarf honey bees. 14 Although the biosynthesis of this compound and other similar bee mandibular pheromones and fatty acids is known, 15 the presence of E-dec-2-enedioic acid has not been reported earlier in any honey bee product.
This is the first report on the biological effect of DM containing E-dec-2-enedioic acid in mammalians and the first bioactivity-guided fractionation study of this honey bee product. The estrogenic activity of DM justifies its use in folk medicine and may lead to new possibilities for its use in estrogen-deficient conditions.
Footnotes
Acknowledgments
We thank Ágnes Csiszár (Department of Pharmacodynamics and Biopharmacy) and Ibolya Herke (Institute of Pharmacognosy) of the University of Szeged for their technical assistance in the experiments.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.