
Abstract
The current survey investigates the effect of four polysaccharides isolated from fresh leek or alcohol insoluble substances (AIS) of leek on the production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) from phagocytes. The ability of the polysaccharides to activate serum complement was also investigated. Despite the lack of antioxidant activity, the pectic polysaccharides significantly decreased the production of ROS by human neutrophils. Polysaccharides isolated from AIS markedly activated RAW 264.7 macrophages for RNS production in a concentration-dependent manner. The Western blot analysis revealed that this effect was due to the stimulation of the inducible nitric oxide synthase protein expression of macrophages. The polysaccharides extracted from AIS with water showed the ability to fix serum complement, especially through the alternative pathway. It was found that the polysaccharide that has the highest complement-fixing effect is characterized by the highest content of uronic acids and the highest molecular weight.
Introduction
P
Innate immunity serves as an essential first-line of defense against microbial pathogens and foreign substances. Phagocytic cells such as macrophages and neutrophils play a key role in innate immunity because of their ability to recognize, ingest, and destroy pathogens by oxidative and nonoxidative mechanisms. Thus, approaches designed to enhance innate immune mechanisms nonspecifically could increase the defense against microbial infections. 7,8 In response to a variety of stimuli, NADPH oxidase present in neutrophils is activated in a phenomenon described as “the respiratory burst,” characterized by the production of superoxide anion, which gives rise to other forms of reactive oxygen species (ROS). In addition to ROS, nitric oxide (NO) and other reactive nitrogen species (RNS) produced mainly by macrophages are one of the important microbicidal tools in the process of inflammation during the fight against pathogenic microorganisms, bacteria, and tumor cells. 9,10 However, excessive or inappropriate ROS and RNS production by phagocytes is associated with oxidative damage to membrane lipids, DNA, proteins, and lipoproteins, resulting in various autoimmune and inflammatory diseases. Thus, the modulation of inflammation and oxidative stress by natural substances can be beneficial.
The complement cascade is another important part of the innate immune defense. Therefore, an activation of the complement system contributes to inflammatory responses and immunological defense reactions. As reported before, interaction with the complement system of polysaccharides due to fixation could be a good therapeutic strategy for treating inflammatory diseases. 11,12
Leek (along with onion and garlic) is part of the traditional Balkan diet, and its medicinal action has been known since ancient times. So far, there is scarce data about the chemical composition of leek polysaccharides and their biological activity, 13 –15 but it was established that some polysaccharides are promising stimulators of some components of the immune system. That provoked our interest to conduct a more extensive study on the effects of leek pectic polysaccharides obtained through different extraction methods on the production of ROS and RNS by phagocytes.
Taken together, the complement system and phagocytes are both recruited by activation processes. Therefore, the main aim of the present study was to investigate the effects of pectic polysaccharides isolated from leek on ROS and NO production by phagocytes as well as on complement activity.
Materials and Methods
Preparation of leek polysaccharides
Fresh leek was purchased from the local market in Plovdiv, Bulgaria. Polysaccharides 1 and 2 (PS1 and PS2, respectively) were obtained from fresh leek by the following procedure: 500 g of leek was homogenized and mixed with 250 mL 0.5% HCl and 250 mL 0.5% citric acid (for PS1) or 500 mL 0.5% HCl (for PS2). The mixtures were incubated with stirring for 1 h at 80°C and centrifuged (4400 g, 30 min). The supernatants were removed, coagulated with 96% ethanol (1/1.5; v/v), and left at room temperature for 1 h. The coagulated polysaccharides were then separated by centrifugation (4400 g, 15 min). Separated polysaccharides were washed consecutively with 70% ethanol and 96% ethanol and dried at 60°C in a laboratory drier.
Polysaccharides 3 and 4 (PS3 and PS4, respectively) were obtained by the following procedure: fresh leek (1 kg) was cut into small pieces of 8–10 mm and treated with 2500 mL 96% ethanol preheated to 65°C. The obtained mixture was kept for 1 h at 65°C followed by 24 h at room temperature, and then filtered through cheese-cloth, after which, the insoluble material was washed with 96% ethanol. The obtained alcohol insoluble substances (AIS) were dried at 60°C. Fifty grams of dried AIS was treated with 1250 mL water at 80°C for 1 h and then filtered. The filtrate was processed to obtain water extractable pectic PS3. The residue was further extracted with 1000 mL 0.5% HCl for 1 h under the same conditions. The mixture was filtered through cheese-cloth, and the filtrate represents the crude acid extractable pectic PS4. The two obtained aqueous filtrates were coagulated using an equal volume of 96% ethanol and left for an hour. The coagulated crude pectins were separated by filtration, washed with 100 mL 70% acidified ethanol (0.5% HCl), then with 70% ethanol to a neutral pH, and finally with 100 mL 96% ethanol. They were dried at 60°C in a laboratory drier.
Determination of polyuronic content and degree of esterification of pectic polysaccharides
The polyuronic content and degree of esterification were determined by the method of Owens. 16
Determination of molecular weight
Molecular weights of the polysaccharides were assayed through high-performance size exclusion chromatography (HPSEC) on a Waters (Millipore) system as described by Kratchanova et al. 15
Cell culture
The murine peritoneal macrophage cell line RAW 264.7 (American Type Culture Collection) was cultivated in Dulbecco's modified Eagle's medium (PAN) and supplemented with 10% of fetal bovine serum (PAN). Cells were maintained at 37°C, 5% CO2.
Whole blood phagocyte-derived ROS production
Heparinized (50 IU/mL) blood samples were obtained from the cubital vein of healthy human volunteers after overnight fasting. The kinetics of ROS production by blood phagocytes was analyzed by luminol-enhanced chemiluminescence (CL) for a period of 60 min, using a microtiter plate Luminometer LM-01T (Immunotech). For activation of ROS production, opsonized zymosan particles (OZP) or phorbol myristate acetate (PMA; Sigma-Aldrich) were used. The detailed procedure is described elsewhere. 17 Spontaneous CL measurements (without any activator) were included in each assay. The recorded values included the intensity of CL emitted during the time interval studied (integral of the CL).
Measurement of nitrite concentration by Griess reaction
The accumulation of nitrites in a medium was measured spectrophotometrically according to Pekarova et al. 10 Cells were incubated in 12-well plates at 1×106 cells/well for 24 h without/with 0.1 μg/mL lipopolysaccharide (LPS, Escherichia coli serotype 0111.B4; Sigma-Aldrich) and pectic polysaccharides at 37°C, 5% CO2. At the end of the incubation period, culture media were collected from wells and centrifuged at 5000 g, 4°C for 5 min. Then, 150 μL of supernatant was mixed with an equal volume of Griess reagent (Sigma) in a 96-well plate, and the mixture was incubated at room temperature and in the dark for 30 min. The absorbance was measured at 546 nm. Sodium nitrite was used as the standard.
Western blot analysis of inducible nitric oxide synthase expression
After removing the supernatant for the nitrite measurement, the remaining cells were washed with cold phosphate-buffered saline and lysed in the lysis buffer (1% sodium dodecyl sulfate, 10−1 M Tris pH 7.4, 10% glycerol, 10−3 M sodium ortho-vanadate, 10−3 M phenylmethanesulfonyl fluoride). Protein concentrations were determined using BCA™ protein assay (Pierce), with bovine serum albumin as a standard. The equal amounts of protein were then subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis, using 7.5% running gel. The expression of inducible nitric oxide synthase (iNOS) protein was quantified by Western blot analysis, as described previously. 18 Anti–iNOS/NOS Type II mouse monoclonal antibody (BIO-RAD; 1:5000) and ECL™ anti–mouse IgG horseradish peroxidase linked whole antibody (from sheep; Biosciences; 1:2000) were used. The immunoreactive bands were detected using an ECL™ detection reagent kit (Pierce) and exposed to radiographic film (AGFA). Equal loading of proteins was confirmed by determination of β-actin. Relative protein levels were quantified by scanning densitometry using the ImageJ™ program, and the individual band density value was expressed in arbitrary units.
Antioxidant activity determination
The antioxidant activity of the investigated polysaccharides was investigated by total peroxyl radical-trapping antioxidant parameter (TRAP) and oxygen radical absorbance capacity (ORAC) assays. 19,20
Complement fixation assay
The test is based on the colorimetric measurement of hemoglobin released from target erythrocytes after incubation with normal human serum. 21 The detailed procedure is described elsewhere. 15
Statistical analysis
The results are presented as mean from at least three independent experiments±standard error of mean (SEM). Experiments were realized in duplicates or triplicates. Comparisons with the control were performed by analysis of variance (ANOVA), followed by the Newman–Keuls Post hoc test. p values of less than .05 and .01 were considered significant.
Results
Characteristics of leek pectic polysaccharides, isolated by different extraction methods
The yield and characteristics of the isolated pectic polysaccharides from leek are presented in Table 1. It is evident that extraction conditions lead to different characteristics of the extracted pectins. Generally, pectic polysaccharides extracted from dry AIS are distinctive, with a lower degree of esterification when compared to the polysaccharides obtained from fresh leek. Water extracted PS3 has the highest polyuronic content −72.8%, and the highest molecular weight −8.8×105 Da.
HPSEC, high-performance size exclusion chromatography; PS1–PS4, polysaccharides 1–4.
The effect of pectic polysaccharides on ROS production
Phagocytes play a key role in host defense against invading pathogens, and play a crucial role in inflammatory processes. In our study, the effect of pectic polysaccharides on ROS production by whole blood neutrophils was analyzed using luminol-enhanced CL. Typical kinetic curves of spontaneous neutrophil CL and neutrophil CL activated with receptor-bypassing (PMA) and receptor-operating (OZP) stimuli are shown in Figure 1. Spontaneous CL represents the basal production of ROS in nonactivated neutrophils. On the other hand, the activation of neutrophils with both PMA and OZP induced a significant increase in ROS production. The pectic polysaccharides in our study decreased both spontaneous and PMA- or OZP-activated ROS production in a 0.1 mg/mL concentration (Table 2). This effect was expressed even more significantly when a concentration of 0.5 mg/mL was used.

Kinetics of luminol-enhanced spontaneous and activated chemiluminescence (CL) of human whole blood neutrophils. The results are expressed in relative light units (RLU).
The data are expressed as % of respective controls and represent mean±SEM, n=3.
Asterisks indicate significant differences at ** P≤.01 or * P≤.05 levels when compared to respective control values (cells incubated without lipopolysaccharide [LPS]).
PMA, phorbol myristate acetate; OZP, opsonized zymosan particles; CL, chemiluminescence; SEM, standard error of the mean.
The effect of pectic polysaccharides on nitrite production and iNOS expression
In our previous experiments, 18 the highest concentration of nitrites in a medium was observed in control RAW 264.7 cells after their 1-day stimulation with LPS, and this schedule was chosen for the experiments. Figure 2 demonstrates that while PS1 and PS2, in both concentrations, do not activate resting macrophages (not treated with LPS) to produce nitrites, PS3 at the concentration of 0.1 mg/mL markedly activated macrophages for nitrite production. This effect became significant (p<.01) when 0.5 mg/mL PS3 was used. PS4 was even more effective activating macrophages very significantly (p<.01) in a 0.1 mg/mL concentration, with a further activation increase in a 0.5 mg/mL concentration. Interestingly, none of the tested pectic polysaccharides influenced the production of nitrites in macrophages treated with LPS (data not shown). The possibility that the changes in nitrite concentration induced with PS3 and PS4 were associated with changes in iNOS protein expression was determined using Western blot analysis (Fig. 3). In comparison with the iNOS protein level in the control macrophages without LPS treatment, iNOS protein expression was stimulated with PS3 and PS4 in a manner very similar to the increase in nitrite concentrations in the cultured media shown in Figure 2. The expression of iNOS protein was not changed in LPS-treated macrophages, which also corresponds with our data on NO production.
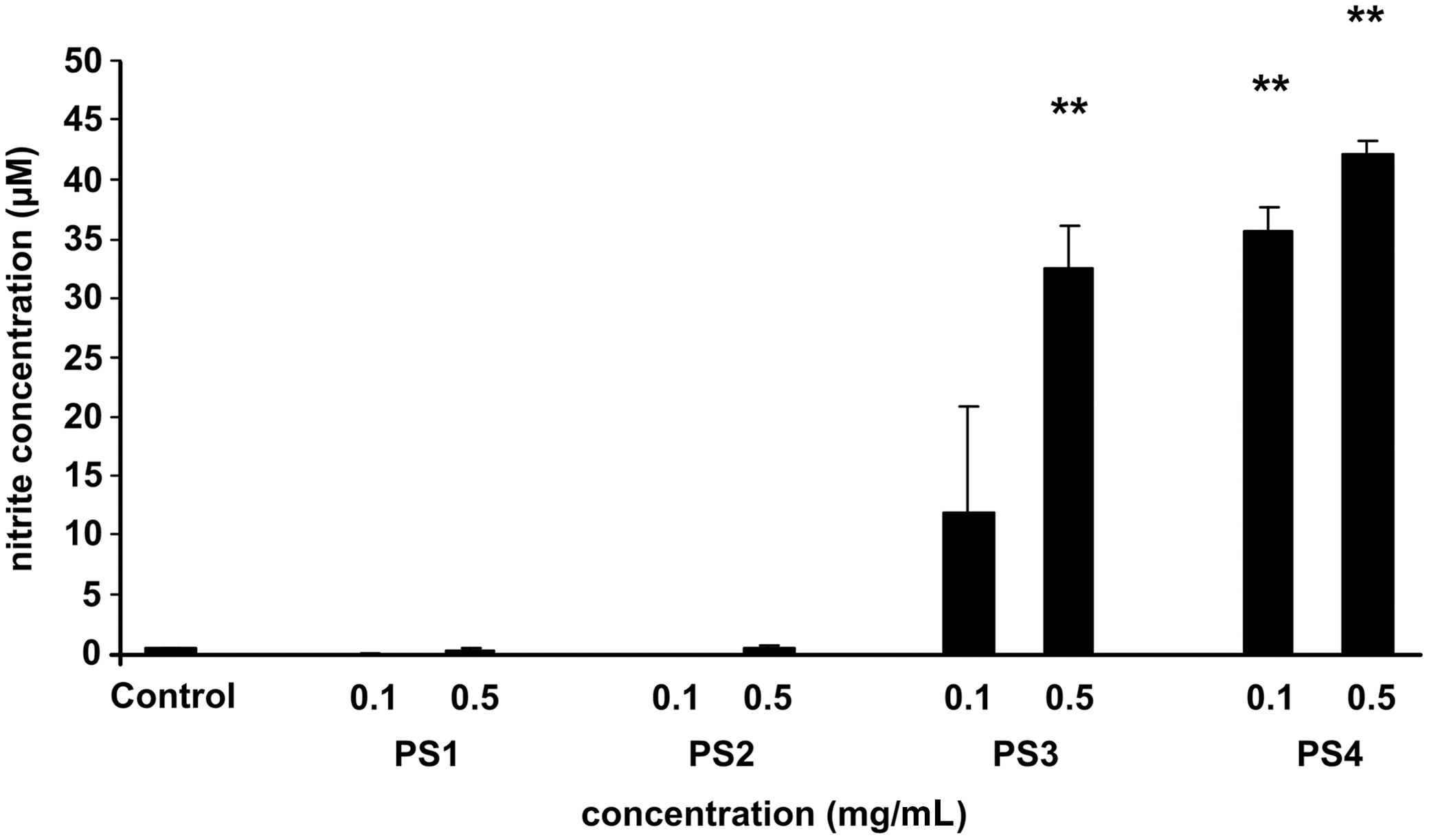
The effect of pectic polysaccharides on NO production by nonstimulated murine RAW 264.7 macrophages. Data are expressed as mean±SEM from three independent experiments. **Indicates significance at level P<.01 when compared with a relevant control. PS1–4, polysaccharides 1–4; NO, nitric oxide; SEM, standard error of the mean.
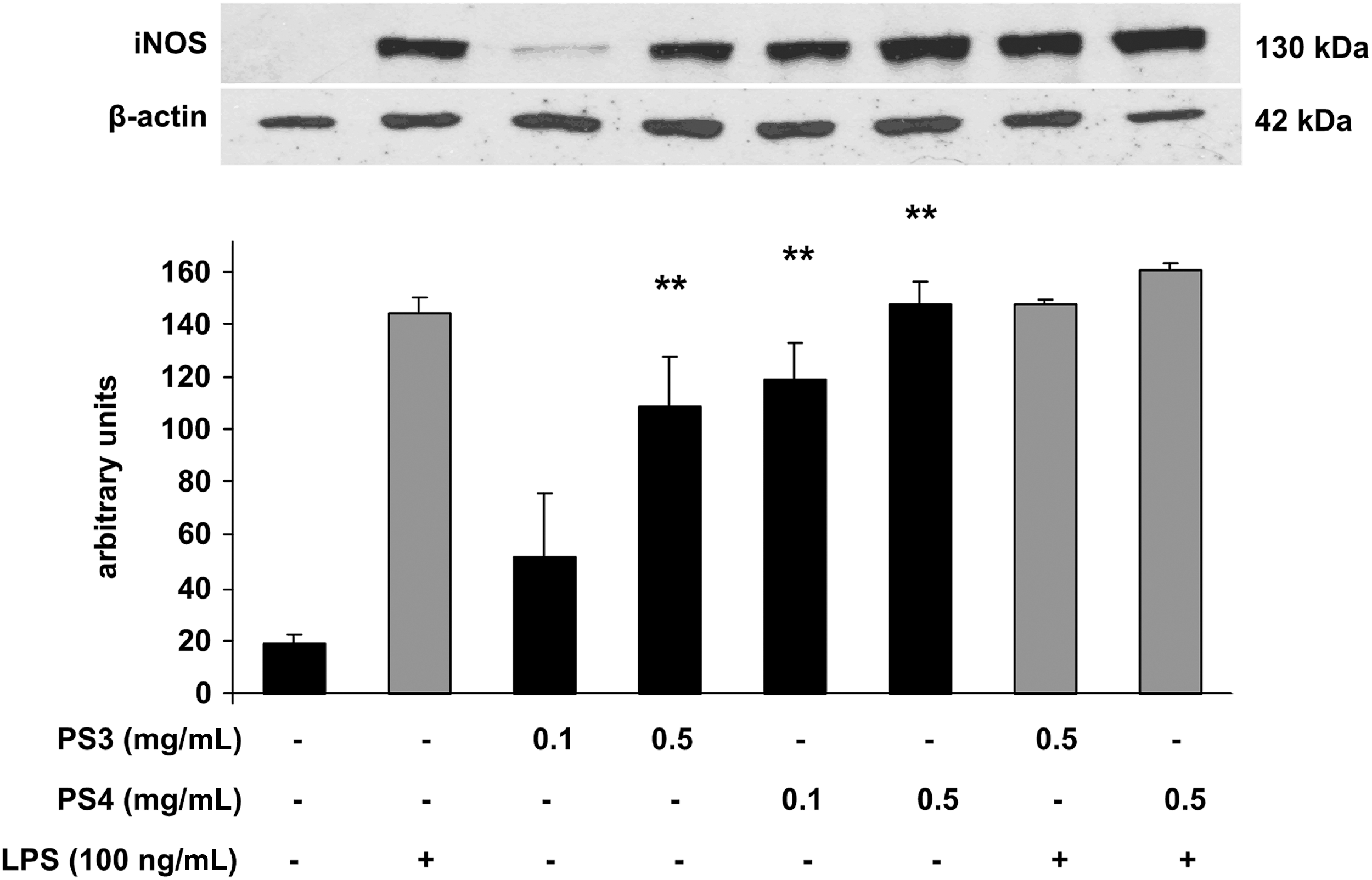
Densitometric analysis and representative Western blot of iNOS protein expression in LPS-stimulated RAW 264.7 cells treated with pectic polysaccharides. The data represent mean±SEM from three independent experiments. Asterisks indicate a significant difference (**P<.01) when compared to the relevant control value (cells incubated without LPS). iNOS, inducible NO synthase; LPS, lipopolysaccharide.
Scavenging properties against the peroxyl radical or NO
The results of some authors (e.g., Kaur et al. 22 ) suggest that polysaccharides isolated from plants may play important roles in free radical scavenging. Therefore, we analyzed whether the antioxidant properties of polysaccharides against ROS and NO can interfere with changes in phagocyte-derived ROS and NO production. All assays used (TRAP, ORAC, and direct amperometrical analysis) confirmed that none of the tested pectic polysaccharides exerted significant scavenging properties against the peroxyl radical or NO (data not shown).
The effect of pectic polysaccharides on the complement system
The polysaccharides were tested for their ability to fix complement. As shown in Table 3, PS1, PS2, and PS4 showed a weak complement-fixing effect, whereas the addition of pectic PS3 resulted in dose-dependent erythrocyte hemolysis, indicating that this fraction exhibited potent complement-fixing activity. The effect was more profound via the alternative pathway of complement activation. Therefore, the fixation of complement by this polysaccharide can be proposed as a potential therapeutic strategy for treating inflammatory diseases.
The data represent mean±SEM from three independent experiments.
Values marked with different superscript letters within individual columns are significantly different (P≤.01).
Discussion
One of the most promising recent alternatives to classical medical treatment is the use of immunomodulators for enhancing host defense responses. In recent decades, polysaccharides, including pectic substances isolated from plant sources, have attracted a great deal of attention because of their broad spectrum of therapeutic properties and relatively low toxicity. 4,5 While our understanding of the mechanism of action of these substances is still developing, it appears that one of the primary mechanisms involves modulation of the phagocyte and the complement activity. 23 In our study, we investigated the effects of pectic polysaccharides isolated from leek on ROS and NO production by neutrophils and macrophages, as well as on the complement activity.
Only limited information exists on the macrophage immunomodulatory activity of pectic polysaccharides; and, to our knowledge, there are no reports regarding the effects of leek polysaccharides on neutrophils. However, in contrast with studies by other authors, where the stimulation effects of some polysaccharides on neutrophils is reported, 8,24 –26 the pectic polysaccharides in our study decreased both spontaneous and PMA- or OZP-activated ROS production in a concentration-dependent manner. The results of some authors 22 suggest that polysaccharides isolated from plants may play an important role in ROS scavenging. Schepetkin et al. 27 found that Opuntia polyacantha polysaccharide fractions exhibited ROS scavenging activity, with the high molecular weight fractions being the most active. In our study, both TRAP and ORAC assays confirmed that none of the tested pectic polysaccharides exerted antioxidant properties against peroxyl radicals, proving that the decrease in neutrophil-derived CL is not caused by the antioxidant properties of the tested polysaccharides.
It is well established that a wide range of plant polysaccharides exhibit beneficial pharmacological effects via their ability to modulate macrophage function, including the production of NO by iNOS. 23 The macrophage-modulating activity of polysaccharides toward NO production has been reported for polysaccharides isolated from different sources, 8,25,26,28 but very little is known about the structural characteristics of the polysaccharides that determine this activity. Schepetkin et al. 27 proposed that the macrophage-immunomodulatory activity of Opuntia polysaccharides is positively correlated with the average molecular weight of the polysaccharides. The authors suggested that polysaccharides may activate macrophages via receptor(s) or other surface structures, although the nature of these surface targets is currently unknown. Contrary to these observations, in our study, polysaccharide PS4 exhibited the most potent macrophage-modulatory properties, by activating macrophages to produce NO, even when a concentration of 0.1 mg/mL was used (Fig. 2). HPSEC analysis showed that this acid-extractable polysaccharide is heterogeneous, with an average molecular weight of 3.2×105 Da, which is the lowest, compared to other polysaccharides isolated from leek. The homogeneous PS3 with the highest molecular weight among leek pectic polysaccharides (8.8×105 Da) also activated macrophages for NO production, although its activity in a concentration of 0.1 mg/mL was lower compared with PS4. At a concentration of 0.5 mg/mL, there was no significant difference in the macrophage-modulatory activity of the two polysaccharides tested. Our results indicate that a high molecular weight is not the only prerequisite for macrophage-immunomodulatory activity. There are probably some structural characteristics that also determine this activity, but further studies are necessary to understand the relationship between the structure and biological activity.
While interpreting results from an in vitro study, one has to keep in mind that the absorption of plant polysaccharides into the bloodstream after oral administration is not well understood and is a disputable issue. Nevertheless, the metabolic activity of neutrophils can be modulated by polysaccharides directly in the intestinal lumen, especially during inflammatory conditions. One of the challenges of the immune system is to ensure, on one hand, a balance between the protection of barrier surfaces from pathogens and, on the other, the establishment of a beneficial relationship with commensal bacteria. 29 In mucosal tissues, the inflammatory response is central to effective host defense against invading pathogens. The mucosal immune system must be tightly regulated to prevent abnormal responses to innocuous environmental antigens and commensal organisms, which could result in an allergy or chronic inflammatory diseases. Many intestinal inflammatory illnesses are marked by neutrophil transepithelial migration and arrival in the lumen, forming crypt abscesses. Here neutrophils not only directly defend the surface, but also release biologically active mediators and stimulate a secretory flush. 30,31 Neutrophil migration across the intestinal epithelium and their presence in the lumen enable pectins to interact directly with the receptors on the surface of neutrophils.
The absorption of plant polysaccharides into the bloodstream after oral administration is another possibility for how pectins can influence the components of the immune system, including neutrophils, monocytes/macrophages, and the complement system. The majority of the intestine is covered with a single layer of columnar epithelial cells, which make up the absorptive surface for water and nutrient absorption. 30 While the intestinal barrier is permeable to digested nutrients as well as fluids, it is, in general, impermeable to macromolecules, particular antigens, and most microorganisms. However, the immune system needs direct contact with antigens or pathogens to generate specific immune responses. For this purpose, the intestine provides specialized epithelial cells, called M cells, which are responsible for the uptake of antigens and microorganisms. The most noticeable feature of M cells is their active transport (by a phagocytic mode of transport that has yet to be fully characterized) of a wide variety of inert material from the gut lumen toward the follicles, from where particles can migrate to the blood via the mesentery nodes and the thoracic lymph duct. 32 Because polysaccharides also behave as hydrated nanoparticles in an aqueous solution, it is postulated that plant polysaccharides become incorporated into the lymphoid follicles of Peyer's patches and isolated lymphoid follicles by a mechanism similar to other nanoparticles, resulting in their dissemination to systemic circulation through the mesenteric lymph nodes and the thoracic duct. Some experimental results suggest that oral administration of certain immunostimulating polysaccharides affects the gastric mucosal immune system through Peyer's patch cells. Otsuka et al. 33 reported that peptide–mannan complexes with a molecular weight of 60,000–95,000, when orally administered to rats, reached the lymph and the blood stream. Mitsuhashi et al. 34 found that α-glucan isolated from the liquid-cultured mycelium of Tricholoma matsutakwith, with a molecular weight over 2.0×106 daltons administered as a 14 C-labeled derivative, was absorbed in the digestive system and remained in the blood stream even 168 h after administration. The peptide–mannan complex, as well as α-glucan, was observed to accumulate in the mesenteric lymph nodes and Peyer's patches as well as in the liver, spleen, and kidneys. Yunoki et al. 35 reported the accumulation of the polysaccharides derived from the basidiomycetes Coriolus versicolor in the liver of mice after oral administration, whereas Sakurai et al. 36 observed the same for the pectic polysaccharide bupleuran 2IIc, from Bupleurum falcatum, which was also accumulated in Peyer's patches. Otsuka et al. 33 indicated that peptide–mannan appeared in aortic blood more than 2 h after it was detected in portal blood, suggesting that the peptide–mannan, which was absorbed in the digestive system was initially incorporated by the phagocytic system of the liver, while the excess of peptide–mannan overflowed into the systemic blood stream. It is assumed that the formation of the immune complexes of the pectic polysaccharide-reacting natural IgA antibody with the active pectic polysaccharide in the intestinal fluid may not only participate in effective incorporation of the active pectic polysaccharides into Peyer's patches, but may also activate complement components that result in certain modulations of the intestinal immune system. 37
The complement can also meet pectins directly in the epithelium or even in the intestinal lumen. In general, the complement system plays an important role as a primary defense against bacterial and viral infections, and appears to be intrinsically associated with several immune reactions, such as the chemotactic attraction of leukocytes, immune adherence, the modulation of antibody production, and increased local vascular permeability. 37,38 According to the study of Kiyohara et al., 37 the pectic polysaccharide-reacting natural and secretory IgA antibody exists in the mucosal sites of the human intestine. Dimeric IgA has been assumed to contribute to activating the alternative complement pathway, producing some biologically active complement fragments, and complement components such as C3 and C4, which are known to be produced by the epithelial cells of the intestinal tract. 39,40 It is also speculated that the formation of immune complexes of the pectic polysaccharide-reacting natural IgA antibody with the active pectic polysaccharides in the intestinal fluid may activate complement components in the fluid, resulting in certain modulations of the intestinal immune system.
As reported by Gasque, 41 complement component proteins can be activated through three cascade pathways: the classical pathway, the alternative pathway, and the antibody-independent lectin pathway. In general, the alternative and lectin pathways contribute to the early natural defense mechanism of the host before production of the antibody. It is known that some plant polysaccharides are good modulators of the immune system, and their activity depends on several factors, such as raw material, extraction conditions, chemical composition, and the structure of the polysaccharide. However, it is disputable which part of the pectic macromolecule is responsible for their immunomodulating properties. Most polysaccharides with immunomodulating properties have been isolated from hot-water extracts of medicinal herbs, 5 and some authors suggest that a high molecular weight and a higher content of galacturonic acids are prerequisites for a higher complement-fixing ability. 3 Our results confirm these observations, and show that PS3 obtained after water extraction of AIS reveals the highest complement-fixing effect and is characterized by a high content of uronic acids and a high molecular weight.
Conclusion
It is evident from the performed study that leek polysaccharides modulate the immune system via different mechanisms. Polysaccharides PS3 and PS4 isolated from AIS exhibited potent macrophage-activating properties resulting in increased generation of NO via the induction of inducible NO synthase. Pectin PS3 obtained after water extraction of AIS revealed the highest complement-fixing effect. These findings make leek a valuable source of biologically active polysaccharides, which could also be considered for further medicinal applications. It was shown that molecular weight is not the only thing that determines the macrophage immunomodulatory activity of leek polysaccharides, and further studies are required to investigate the structural features in relation to the biological activity.
Footnotes
Acknowledgments
This study was supported by grants IF-02-66/2007 (Bulgarian Innovation Fund) and 524/08/1753 (Czech Science Foundation).
Author Disclosure Statement
No competing financial interests exist. All authors report no conflicts of interest.