
Abstract
In this study, the protective effects of Brussels sprouts extract and its major constituents against oxidative stress-induced damages were investigated in rat pheochromocytoma cells and Institute of Cancer Research mice. The major constituents of Brussels sprouts (3,4′,5,7-tetrahydroxyflavone (kaempferol), indole-3-carbinol, and phenethyl isothiocyanate) were selectively tested. Of these, the flavonoid compound, kaempferol exhibited the highest potency in radical scavenging activity (1,1-diphenyl-2-picryl hydrazyl assay and oxygen radical absorbance capacity assay) and was most protective against oxidative stress in neuronal cell assays (measurement of intracellular oxidative stress levels and cell viability). In mice, after 4 weeks of kaempferol administration, significant protection against amyloid beta (Aβ) peptide-induced neurotoxicity was also observed, as assessed through the passive avoidance test. Taken together, the results suggest that Brussels sprouts could be protective against Aβ-induced neurotoxicity, possibly due to the antioxidative capacity of its major constituent, kaempferol.
A
There has been increasing interest in cruciferous vegetables such as Brussels sprouts owing to their anticarcinogenic properties. Glucosinolates, a class of compounds found in cruciferous vegetables, and their enzymatic degradation products have been acknowledged as potential anticancer agents, through their ability to modulate biotransformation enzymes both in vitro and in vivo. 3,4 However, the protective effects of other similar organic compounds found in Brussels sprouts have not been studied in neuronal models. Thus, the objective of this study was to examine the attenuating effects of Brussels sprouts extract on oxidative stress in a neuronal cell line and mouse model. Our hypothesis was that if Brussels sprouts extract elicits protection against oxidative stress, one of their major phytochemical components, which represents the most potent antioxidative capacity, would be responsible for such a protection and this compound will show similar protective effects against Aβ-induced neuronal deficits in an animal model.
2′,7′-dichlorofluorescein diacetate (DCF-DA), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
The sample vegetable, Brussels sprouts, was obtained from the local grocery stores in Seoul, Republic of Korea. Once obtained, the species of vegetable was confirmed again utilizing photographs of the database, provided by Korea Food and Drug Administration. To prepare the sample extract, stalks and surplus stems were removed and only buds were used. The sample was finely ground and five times the sample weight of ethanol was added. The vegetable was extracted for 24 h and filtered through Whatman ashless filter paper no. 2 (Whatman International Ltd., Maidstone, United Kingdom). Residual samples were collected and extracted two more times. The extracts were pooled and then evaporated under nitrogen for further analyses.
Cell viability was evaluated using the MTT reduction assay. PC12 cells (American Type Culture Collection, Manassas, VA, USA) were pretreated with samples for 48 h. After incubation with or without freshly prepared hydrogen peroxide (3 mM; Sigma) for 2 h, the cell viability was determined by the conventional MTT reduction. 5 The amount of MTT formazan crystal was quantified using a microplate reader (GENois TECAN, Männedorf, Switzerland).
Intracellular oxidative stress levels were evaluated using the DCF-DA assay. PC12 cells were pretreated with samples for 48 h. The cells were treated with or without freshly prepared hydrogen peroxide (100 μM) for 2 h, and then 50 μM of DCF-DA (Sigma) was applied for 50 min. The DCF was quantified by the fluorometer (GENois TECAN). The results were expressed in percent relative to the oxidative stress of the control group set to 100%.
The antioxidant capacity of major compounds of Brussels sprouts (I3C, PEITC, and kaempferol; Sigma) was measured using DPPH and oxygen radical absorbance capacity (ORAC) assay. 6,7
Briefly, sample stock solutions were mixed with DPPH (Sigma) in methanol and incubated at 37°C for 4 h. After the reaction was completed, absorbance was measured at 515 nm, and the antioxidant capacity was calculated based on the slope of the Trolox stock solutions using a spectrophotometer (Genesis 10S; Thermo Scientific, Madison, WI, USA).
On the other hand, AAPH (Sigma) was used as a peroxy radical generator for the ORAC assay. The final reaction mixture was composed of 50 μL sample, 100 μL fluorescein, and 100 μL AAPH. Once the AAPH was added, the plate was shaken automatically for 5 seconds, and then measured every 1 min for 1 h with emission and excitation wavelengths of 485 and 525 nm, respectively, using a microplate fluorescence reader Synergy HT (Bio-Tek Instruments, Inc., Winooski, VT, USA). The reaction temperature was maintained at 37°C. The final ORAC values were calculated by using a quadratic regression equation (y=a+bx+cx2).
A semipurified commercial mouse diet was supplemented with 0.0006%, 0.0012%, and 0.0024% kaempferol (w/w; 6, 12, and 24 ppm, respectively). Institute of Cancer Research (ICR) mice (Daehan Biolink, Chungbuk, Republic of Korea) had free access to food and water for 4 weeks. After 4 weeks, the Aβ 1-42 peptide (410 pmol per mouse; 5 μL injection volume; Sigma) was delivered by means of intracerebroventricular (ICV) injection to induce amnesic symptoms. The negative control group was injected with the nontoxic reverse Aβ 42-1 peptide (Bachem, Bubendorf, Switzerland). One week after the injection of Aβ peptides, the passive avoidance response of ICR mice was assessed as previously described. 8 Further, the level of lipid peroxidation in the brain tissues was measured as described elsewhere. 9 All animal experiments followed the guidelines provided by the Animal Care and Use Committee of Korea University.
The results are presented as mean±standard deviation. The statistical significance of differences among groups was calculated by one-way ANOVA followed by the Scheffe's multiple range test by using SAS (Cary, NC, USA). A P-value <.05 was considered statistically significant.
In the present study, the PC12 cell line was utilized for the in vitro model. The PC12 cell line is originally isolated from the adrenal medulla tumor and it is known that it displays the properties associated with sympathicoblasts; for instance, the major catecholamine, neurotransmitter contents, and the acetylcholine sensitivity of the PC12 cell line are similar compared to that of sympathetic neurons. 10 Therefore, it is one of the most widely utilized cell lines for in vitro neuronal models. 11,12
The cell viability and intracellular oxidative stress were measured using MTT and DCF-DA assays, respectively. Brussels sprouts extracts did not exhibit the significant effects in any concentration, but there was a trend of increase in cell viability with the highest concentration of Brussels sprouts treatment. In contrast, intracellular oxidative stress levels were significantly decreased by Brussels sprouts treatment in all levels. Notably, the level of the DCF formation was inhibited ∼1300% compared to that of positive control at the highest concentration (Fig. 1). These results led us to further investigate the compounds in Brussels sprouts that may be responsible for such protection. According to various studies, Brussels sprouts contains several flavonoids, of which kaempferol is the most abundant compared to other major flavonoids such as luteolin and quercetin. 13,14 Further, there is a class of phytonutrients found in Brussels sprouts and other cruciferous vegetables called glucosinolates, which have been extensively investigated with regard to their anticancer effects. 3,4,15 Thus, we also included two breakdown products of glucosinolates (PEITC and I3C) in this study.
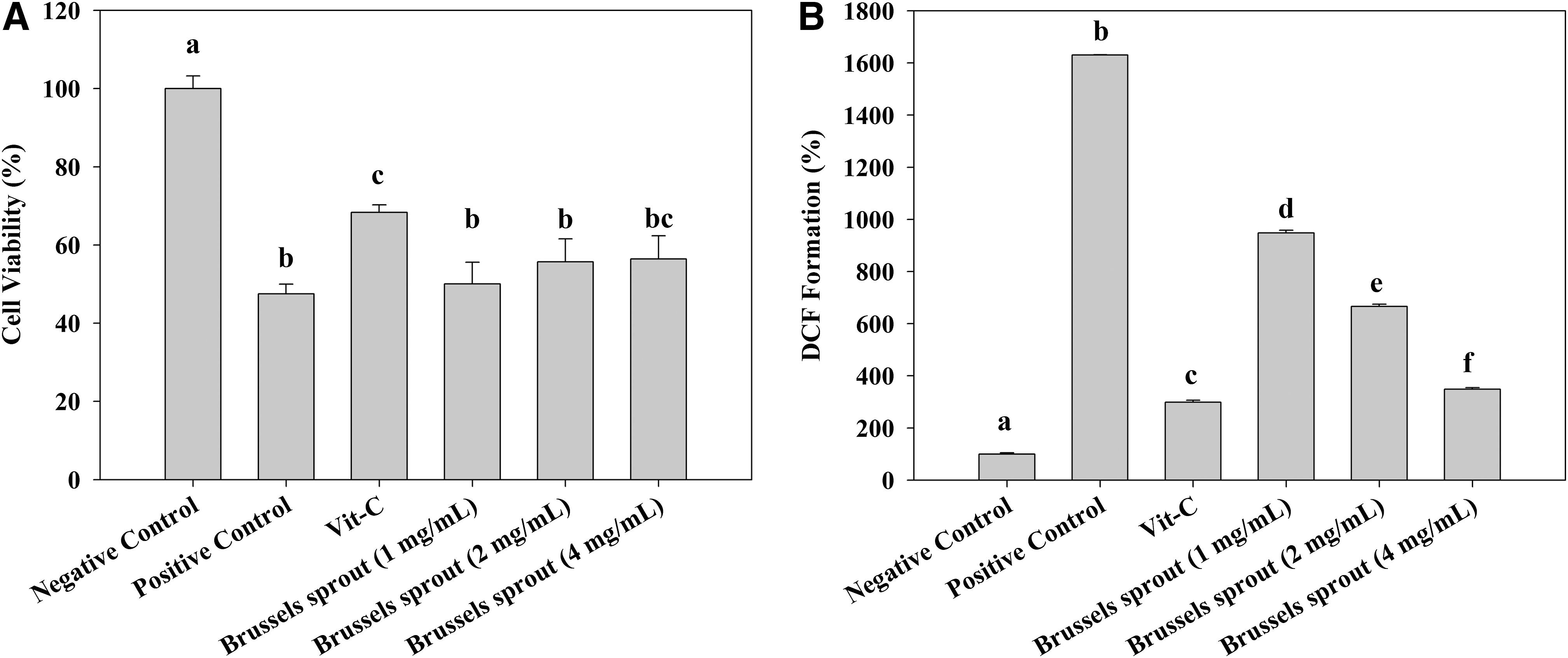
Effects of Brussels sprouts extracts on neuronal cell viability
Three major components (I3C, PEITC, and kaempferol) were selectively tested on PC12 cells and the radical scavenging activity of each compound was determined by DPPH and ORAC assays. In all assays, kaempferol exhibited the most potent activity of the three major components. Kaempferol showed approximately 84% and 42% in cell viability and inhibition of DCF formation compared to positive controls. Similarly, in both DPPH and ORAC assays, kaempferol represented the most potent antioxidative activity (Table 1). Although I3C exhibited relatively high potency in DPPH assay, notable effects on MTT and DCF inhibition were not observed. Similarly, PEITC exhibited negligible potency in all in vitro assays tested. Therefore, the most potent compound among the tested phytonutrients, kaempferol, was supplemented in diets for in vivo behavioral testing.
Data represent the mean±standard deviation (n=3).
Radical scavenging capacity (%)=100–[(positive control–sample group)×100].
The final values were calculated by using a quadratic regression equation between Trolox concentration and the net area under the fluorescence decay curve. Data are expressed as micromole of Trolox equivalents (TE) per gram of sample (μmol TE/g).
Scheffe's multiple-range test indicated a significant difference. Different superscript letters represent statistical differences between groups (P<.05).
DCF, dichlorofluorescein; DPPH, 1,1-diphenyl-2-picrylhydrazyl; ORAC, oxygen radical absorbance capacity; PEITC, phenethylisothiocyanate.
In the passive avoidance test, mice treated with the Aβ 1-42 peptide showed a significant decrease in step-through latency compared to mice treated with the nontoxic reverse Aβ 42-1 peptide (41.7%±7.8% for Aβ 1-42 peptide vs. 100%±12.5% for reverse Aβ 42-1 peptide). Mice fed with kaempferol supplemented at 0.0012% or 0.0024% exhibited a significant improvement in step-through latency time compared to mice lacking kaempferol supplemented food (positive control group). Effects of kaempferol supplementation on oxidative stress in brain tissues were measured as well. In results, supplementation of kaempferol at 0.0012% or 0.0024% showed a significant reduction in the formation of thiobarbituric acid reactive substances (TBARS) in the brain (170.8%±6.8% and 155.5%±8.6%, respectively). No effect was observed in the group that received 0.0006% kaempferol supplementation (Table 2). Given the poor bioavailability of flavonoids, this might be because the ingested kaempferol may not have been delivered to the target tissue. Recently, it was reported that the bioavailability of this compound was approximately 2.7% and 1.9% when orally administrated at doses of 100 and 250 mg/kg, respectively. 16 However, the pharmacodynamic properties of kaempferol have not been investigated in the brain of mice. Thus, further studies are warranted to understand both the bioavailability in the circulatory system and the permeability against the blood–brain barrier.
Data represent the mean±standard deviation (n=10).
Step-through latency was measured up to 300 s.
The negative control group was set to 100%.
Scheffe's multiple-range test indicated a significant difference. Different superscript letters represent statistical differences between groups (P<.05).
TBARS, thiobarbituric acid reactive substances.
A number of studies have reported that oxidative stress and related damage correlate with AD, and abnormal deposition of Aβ is known to increase the intracellular oxidative damage and contribute to an inflammatory response. 17,18 In addition to direct damage from oxidative stress, indirect damage due to byproducts of lipid peroxidation (e.g., 4-hydroxynonenal) is also likely. 19,20 Therefore, administration of antioxidants that block or suppress these detrimental processes of Aβ might protect against or prevent AD. In our study, dietary supplementation with kaempferol at concentrations of 0.0012% or 0.0024% significantly decreased TBARS formation in the brain, indicating the protective effects of kaempferol against Aβ.
To induce learning deficit and memory impairment in mice, the Aβ peptide was delivered by means of ICV injection in the study. The Aβ infusion rodent model is a widely utilized in vivo model in AD research. This model offers some advantages over the use of transgenic animal models (e.g., amyloid precursor protein transgenic model). To be specific, the infusion model allows researchers to determine the dosage and type of Aβ species. Furthermore, it is rather time-consuming to develop plaques and AD-like symptoms in transgenic animal models, whereas Aβ infusion quickly induces deficits in memory and learning ability. 21
In the study, the level of Aβ was determined on the basis of previous experience in the laboratory. Administration of the Aβ peptide successfully decreased the step-through latency of mice and increased the lipid peroxidation levels in the brain, indicating that the dose of Aβ used in the study was appropriate (Table 2). We did not observe significant differences in food intake, body weight, brain tissue weight, and biomarkers of acute toxicity (i.e., aspartate aminotransferase and alanine aminotransferase activities) in any group (data not shown).
One of the possible limitations of the study is the dose required for an effect, as it is difficult to achieve concentrations of active compound that were used in this study by dietary consumption of fresh Brussels sprouts alone. On the basis of the food intake of mice, it is estimated that a human adult (70 kg body weight) should consume approximately 210 mg of kaempferol (0.0024%), requiring the consumption of an unrealistic amount of vegetables as part of the diet. Therefore, it might be more reasonable to administer kaempferol as a supplement.
As aforementioned, cruciferous vegetables have been extensively investigated regarding their modulating activity on biotransformation enzymes. 3,4 In like manner, Brussels sprouts have gained much interest on their effects on carcinogen metabolism rather than antioxidative activity. However, there are some evidence suggesting that this vegetable may elicit anticarcinogenic potency through not only modulation of biotransformation enzymes, but also preventing oxidative DNA damage, which is in agreement with our observation. 22,23 Thus, this study would provide preliminary data to screen candidate components from Brussels sprouts, which are responsible for the protection against oxidative damage of DNA.
In summary, we demonstrated that crude extracts of Brussels sprouts exhibited inhibitory effects against intracellular reactive oxygen species formation in PC12 cells. In subsequent in vitro assays, kaempferol represented the highest potency of selectively chosen phytonutrients. Utilizing an in vivo mouse model, administration of Aβ successfully induced deficits in short-term memory, and these deficits were significantly reversed by dietary intake of kaempferol. Furthermore, lipid peroxidation levels were decreased by this flavonoid, indicating that protective effects of kaempferol might be due, in part, to its antioxidative potency, thereby decreasing Aβ neurotoxicity. Taken together, our results suggest that kaempferol, by protecting against oxidative damage, might exhibit beneficial effects in neurodegenerative diseases such as AD.
Footnotes
Acknowledgment
This study was supported by a Korea University Grant.
Author Disclosure Statement
The authors declare no competing financial interests.