
Abstract
Tucuma (Astrocaryum aculeatum) is an Amazonian fruit that presents high levels of carotenoids and other bioactive compounds such as quercetin. The extracts of tucuma peel and pulp present strong antioxidant activity which illustrate an elevated concentration that causes cytotoxic effects in human peripheral blood mononuclear cells (PBMCs). This study performed additional investigations to analyze the potential genotoxic effects of the tucuma extracts on PBMCs. The genotoxicity was evaluated by DNA fragmentation, Comet assay, and chromosomal instability G-band assays. The acute tucuma extract treatment showed genoprotective effects against DNA denaturation when compared with untreated PBMC cells. However, in the experiments with 24 and 72 h treatments to tucuma treatments, we observed low genotoxicity through a concentration of 100 μg/mL, some genotoxic effects related to intermediary concentrations (100–500 μg/mL), and more pronounced genotoxic effects on higher tucuma extract concentrations. After 24 h of treatment, the reactive oxygen species were similar among treatments and PBMC control groups. However, the caspase-1 activity related to the apoptosis and pyroptosis process increased significantly in higher tucuma concentrations. In summary, tucuma extracts, despite their higher antioxidant content and antioxidant activity, would present PBMCs genotoxic effects that are dependent on concentration and time exposition. These results need to be considered in future in vitro and in vivo studies of tucuma effects.
Introduction
T
Among the compounds present in vegetables, the carotenoids have attracted significant attention due to their characteristic biological activities, including anti-allergic, anti-cancer, and anti-obesity effects. 5 Some Amazonian fruits can be considered ideal natural carotenoid sources for consumption or supplementation because of the concomitant presence of high amounts of oil and carotenoids in these fruits that improve the oral bioavailability of these compounds. 6
Tucuma (Astrocaryum aculeatum), an Amazonian palm fruit, is one of the traditional components of the regional breakfast. Due to its high popularity, the tucuma can be sold in the Manaus local markets as well as on downtown streets. 7 In Brazil, the common names of this fruit are tucuma or tucum, whereas in other Latin American regions, these species are named wara awara, awarra, and chontilla. 7
Fruit centesimal composition analysis showed that the mesocarp contains 412.73±2.12 kcal, 44.9±0.30 wet content, 10.9 fibers±0.1, 3.5±0.07 g proteins, 8.5±0.6 carbohydrates, and 40.5±0.5 fats per 100 g pulp. The oil extract from the tucuma pulp presents 74.4% unsaturated and 25.6% saturated fatty acids that are rich in Omega 3, 6, and 9 fatty acids. Micronutrient analysis showed that tucuma presents one of the highest concentrations of provitamin A “beta-carotene” with 52 mg per 100 g of pulp. This concentration is approximately eight times higher than that which is found in carrots (6.6 mg/100 g pulp). 8
The tucuma carotenoid's evaluation described 24 carotenoids, of which 21 were chemically identified. The all-trans-β-carotene was found to be the main carotenoid, representing 75% of the total carotenoid content in tucuma followed by 13-cis-β-carotene, all-trans-α-carotene, and all-trans-β-cryptoxanthin, with each representing from 2.0% to 2.8% of the total carotenoid content. The other 19 carotenoids represent 15% of the total content. 8
Previous studies defined other bioactive compounds in tucuma pulp as catechins (79±5 mg/100 g sample dry weight), quercetin (2.96±0.05 mg/100 g sample dry weight) a general antioxidant capacity as well as antiinflammatory activity. 9,10
Within this context, the tucuma extracts from peel and pulp could represent a potential effect on human health. However, it will be important to determine the range of concentrations that do not present toxic effects on cells. For this reason, we performed a complementary investigation to evaluate whether tucuma extracts present some genotoxic effect using human peripheral blood mononuclear cells (PBMCs), such as an in vitro experimental model.
Materials And Methods
Tucuma extract production and chemical characterization
Tucuma peel and pulp extracts were obtained from a composite sample representing a mixture of progenies from a native forest near Manaus City (Amazonas State, Brazil), located in the Amazonian region (3.08°S, 60.01°W). The pulp and peel were manually removed and kept frozen at −18°C until extraction procedures were performed 1 week later, producing 800 g peel and 400 g pulp. The ethanolic tucuma extract was prepared from tucuma pulp and peel samples that were titrated and placed separately into sealed amber glass jars containing an absolute ethanol solution, at a ratio of 1:5. The extraction was performed over 4 days.
The homogenate was filtered through Whatman No. 1 paper and then collected. The ethanol was removed using a rotary evaporator at reduced pressure, 25°C at 2 g. After this procedure, the pulp and peel extracts were lyophilized and stored at −20°C until they were to be used. We obtained 3359 g of peel and 6091 g of dried tucuma pulp extract.
Chemical characterization of tucuma extracts was performed in triplicate. The total phenolic content (TPC) was evaluated using Folin–Ciocalteu's method. 11 The TPC samples were calculated as a gallic acid equivalent from the calibration curve of gallic acid standard solutions (5, 10, 15, 20, 25, and 30 μg/mL) of 0.2% aqueous gallic acid that followed the same method. The equation obtained for the calibration curve of gallic acid was y=18.185x−0.0266 (R 2=0.9783). The results were expressed as mg gallic acid equivalent (mg GAE)/g dry plant extract materials.
The tannin content was measured using the method described by Morrison et al. 12 with some modifications. Samples in concentrations of 0.25 mg/mL, 5 mL of solution A (1 g vanillin in 100 mL of methanol), and solution B (8 mL HCl in 100 mL of methanol) were used for the experiment. The samples were read at 500 nm in a spectrophotometer. The total tannin content was expressed in milligram equivalents of catechin per gram of each fraction. The equation obtained for the calibration curve of catechin in the range of 0.001–0.025 mg/mL was y=0.00015x+0.005 (r=0.9989).
The alkaloid content was determined using the method described by Sreevidja and Mehrotra, 13 where Dragendorff's reagent precipitates alkaloids in plant materials. The samples were read at 435 nm in the spectrophotometer. The equation obtained for the calibration curve of bismuth nitrate pentahydrate solution in the range of 0.01–0.09 mg/mL was y=2.2783x+0.0361 (r=0.9997).
The quantification of β-carotene and quercetin investigation was performed using reverse-phase chromatographic analyses (HPLC). The HPLC analyses were carried out under gradient conditions using the C18 column (4.6 mm×250 mm) packed with 5 μm diameter particles; the mobile phase was water containing 2% acetic acid (A) and methanol (B), and the composition gradient was 5% of B for 2 min, then changed to obtain 25%, 40%, 50%, 60%, 70%, and 100% B at 10, 20, 30, 40, 50, and 80 min, respectively, following the method described by Laghari et al., 14 with slight modifications. The lyophilized tucuma peel and pulp extracts were analyzed and dissolved in ethanol at a concentration of 3 mg/mL. Identification of the compounds was performed by comparing their retention time and UV absorption spectrum with commercial standards. The flow rate was 0.8 mL/min, injection volume was 40 μL, and the wavelength was 365 nm for quercetin and 450 nm for β-carotene. All the samples and the mobile phase were filtered through a 0.45 μm membrane filter (Millipore, Billerica, MA, USA) and then degassed by an ultrasonic bath before use. Stock solutions of standard references were prepared in the HPLC mobile phase at a concentration range of 0.020–0.200 mg/mL for β-carotene and quercetin. The chromatography peaks were confirmed by comparing its retention time with those of reference standards and by DAD spectra (200 to 600 nm). The calibration curve for quercetin is y=13,495x−1092.6 (r=0.9999) and β-carotene: y=13,681x−1518.7 (r=0.9999).
Tucuma extract treatments
To perform the experimental assays, the extracts were first diluted in 40% ethanol in order to improve their solubilization. The solution was then diluted in a cellular medium culture to obtain <0.5% ethanol concentration in all tucuma concentrations tested here. The ethanol was also added in all control treatments to eliminate the potential influence of this molecule in the results obtained. The concentration of tucuma extracts tested in this study was 5, 10, 100, 500, and 1000 μg/mL.
Human PBMC culture conditions
We used PBMC cells from three healthy human donors to test the potential in vitro genotoxicity of tucuma extracts as previously described in Montagner et al. 15 Briefly, peripheral blood samples were collected after a 12 h overnight fasting period by venipuncture using top Vacutainer (BD Diagnostics, Plymouth, United Kingdom) tubes with heparin. One aliquot of each sample was used for cell culture; the other aliquots were used for biochemical and DNA damage analysis. Blood specimens (5 mL) were routinely centrifuged within 1 h of collection for 15 min at 2500 g; leucocyte samples were used to obtain PBMCs through a gradient centrifugation. The PBMCs were immediately transferred to culture media containing RPMI 1640 (Gibco, Life Technologies, Grand Island, NY, USA) with 10% fetal calf serum (FCS) and 1% penicillin/streptomycin. The cells were maintained in a suspension culture at 37°C in a 5% humidified CO2 atmosphere.
Genotoxicity assays
We performed five general protocols to test whether the tucuma extracts present in vitro genotoxic effects on human PBMCs.
DNA picogreen denaturation assay
The DNA denaturation assay was applied by a protocol previously described by Batel et al. 16 The method is based on the ability of the specific fluorochrome dye (PicoGreen®) to make a very stable complex with dsDNA in alkaline conditions instead of ssDNA, proteins, SDS, and urea. This selectivity characteristic is used to follow DNA denaturation with decreasing fluorimetric signal intensity proportionate to increasing ssDNA and mononucleotide content. Initially, we evaluated the potential acute effect of tucuma extracts on DNA denaturation.
Two tests were performed: acute effect of tucuma extracts on DNA damage and after 24 h of PBMC treatment to tucuma extracts. In the acute effect experiment, a kinetic of DNA denaturation (15, 30 and 60 min) was evaluated. The fluorescence was measured at an excitation of 485 nm and an emission of 520 nm recorded at room temperature (SpectraMax M2/M2e Multi-mode Plate Reader; Molecular Devices Corporation, Sunnyvale, CA, USA). Expressions of results to DNA strand-break calculations were made in relation to control values at each time in denaturation after correction for blank readings. Blank values were measured in wells containing only the samples. Results were expressed as a percentage of dsDNA calculated for each treatment in relation to the untreated control samples.
Alkaline DNA comet and chromosomal instability by cytogenetic G-band assays
In the second protocol, we tested the potential genotoxicity of tucuma extracts. In this experiment, we evaluated the effect of tucuma extracts at different concentrations on DNA damage using alkaline DNA Comet and chromosomal instability by cytogenetic G-band assays.
The Comet assay was used to test the effect of tucuma extracts on PBMCs cellular DNA damage. The assay was performed as described by Singh et al. 17 in accordance with general guidelines. Overall, 100 cells (50 cells from each of the two replicate slides) were selected and analyzed. Cells were visually scored according to tail length and received scores from 0 (no migration) to 4 (maximal migration). Therefore, the damage index for cells ranged from 0 (all cells with no migration) to 400 (all cells with maximal migration). The slides were analyzed under blind conditions by at least two different researchers.
Karyotype analysis can provide clues to significant genes involved in the genesis and growth of several cancers. Therefore, we tested whether the tucuma extracts induced changes in mitotic index (MI) and chromosomal instability evaluated by the G-band cytogenetic carried out by optical microscopy analysis, that is, the most commonly used standardized method to detect chromosomal abnormalities. 18 The nuclei found at the centerline of the slide glass were observed, counted, and classified by considering the occurrence of intact nuclei (in interphase or prophase), intact metaphases, and three indicators of chromosomal instability: broken chromosomes, karyiopyknosis, and karyyorrhexis/karyolysis. The karyopyknosis is characterized by a cytological condition caused by shrinkage of the nucleus of a cell with the condensation of the chromatin into structure-less masses. The karyyorrhexis and karyolysis are the destructive fragmentation of the nucleus of a dying cell by which its chromatin is distributed irregularly throughout the cytoplasm that occurs as a result of either programmed cell death or necrosis. The MI that indicates the cell proliferation level was also evaluated to each of the treated and untreated PBMC samples. The MI observed was represented by the percentage of metaphases divided by the total nuclei observed in each treatment. Two replicates of each treatment were performed and analyzed.
Reactive oxygen species assay
We evaluated whether the tucuma extracts in different concentrations present effects on cellular oxidative metabolism, altering the intracellular reactive oxygen species (ROS) production that could be related to genotoxic effects. Therefore, ROS production was determined in all PBMC treatments with 24 h of tucuma extract treatments using the non-fluorescent cell permeating compound 2′-7′-dichlorofluorescein diacetate (DCFDA) assay. In this technique, the DCFDA is hydrolyzed by intracellular esterases to DCFH, which is trapped within the cell. This non-fluorescent molecule is then oxidized to fluorescent DCF by cellular oxidants. After the designated treatment time, the cells were treated with DCFDA (10 μM) for 60 min at 37°C. In the assay, 1×105 a cell to each sample was used to measured ROS levels.
The fluorescence was measured at an excitation of 488 nm and an emission of 525 nm (SpectraMax M2/M2e Multi-mode Plate Reader; Molecular Devices). All tests were performed in triplicate for each of the samples tested. The results were expressed as % of non-treated control cells where the absorbance observed in the treatments was divided by the average absorbance of control group. This value was multiplied by 100 to give the percentage of ROS in relation to the control group. Values above 100% indicate increased levels of ROS, while those below 100% denote lower levels of ROS. 19
Caspase-1 activity assay
The potential programmed cell death induction was analyzed from the evaluation of caspase-1 activity 20 that is involved in pyroptosis and also apoptosis processes of PBMCs treatment to tucuma extracts at different concentrations. The PBMCs were incubated under the conditions described earlier for 24 h. The analyses were performed using the Quantikine Human Caspase-1 Immunoassay to measure caspase-1 in cell culture supernates using a microplate with 96 wells, according to the manufacturer. Briefly, all reagents and working standards were prepared, and the excess microplate strips were removed. The assay diluent RD1W was added (50 μL) to each well. Furthermore, 100 μL of the standard control for our sample was added per well, covered with an adhesive strip, and incubated for 1.5 h at room temperature. Each well was aspirated and washed twice, for a total of three washes. Caspase-1 antiserum was added to each well and covered, given a new adhesive strip, and incubated for 30 min at room temperature. The aspiration/wash step was repeated, and the caspase-1 conjugate (100 μL) was added to each well and incubated for 30 min at room temperature. The aspiration/wash step was repeated, and 200 μL of substrate solution was added to each well and incubated for 20 min at room temperature. Finally, the 50 μL stop solution was added to each well, and the optical density was determined within 30 min using a microplate reader set to 450 nm. Experiments were performed in triplicate.
Statistical analysis
Data are presented as mean±standard error of the mean. Statistical differences between pulp and peel bioactive compound concentrations were compared by Student's t-test. The comparison among genotoxic effects of tucuma extracts at different concentrations was performed by analyzing the one-way variance followed by a Tukey post hoc test. Chromosomal instability frequency among treatments was compared by chi-square test. All statistical analyses were performed where all P-values were two tailed, and P<.05 was considered statistically significant.
Results
The TPC value was 941.8±0.07 mg/GAEg in peel and 872.1±0.06 mg/GAEg in pulp extracts. The chemical characterization of other bioactive compounds also showed higher concentrations in peel when compared with pulp extracts (Fig. 1).
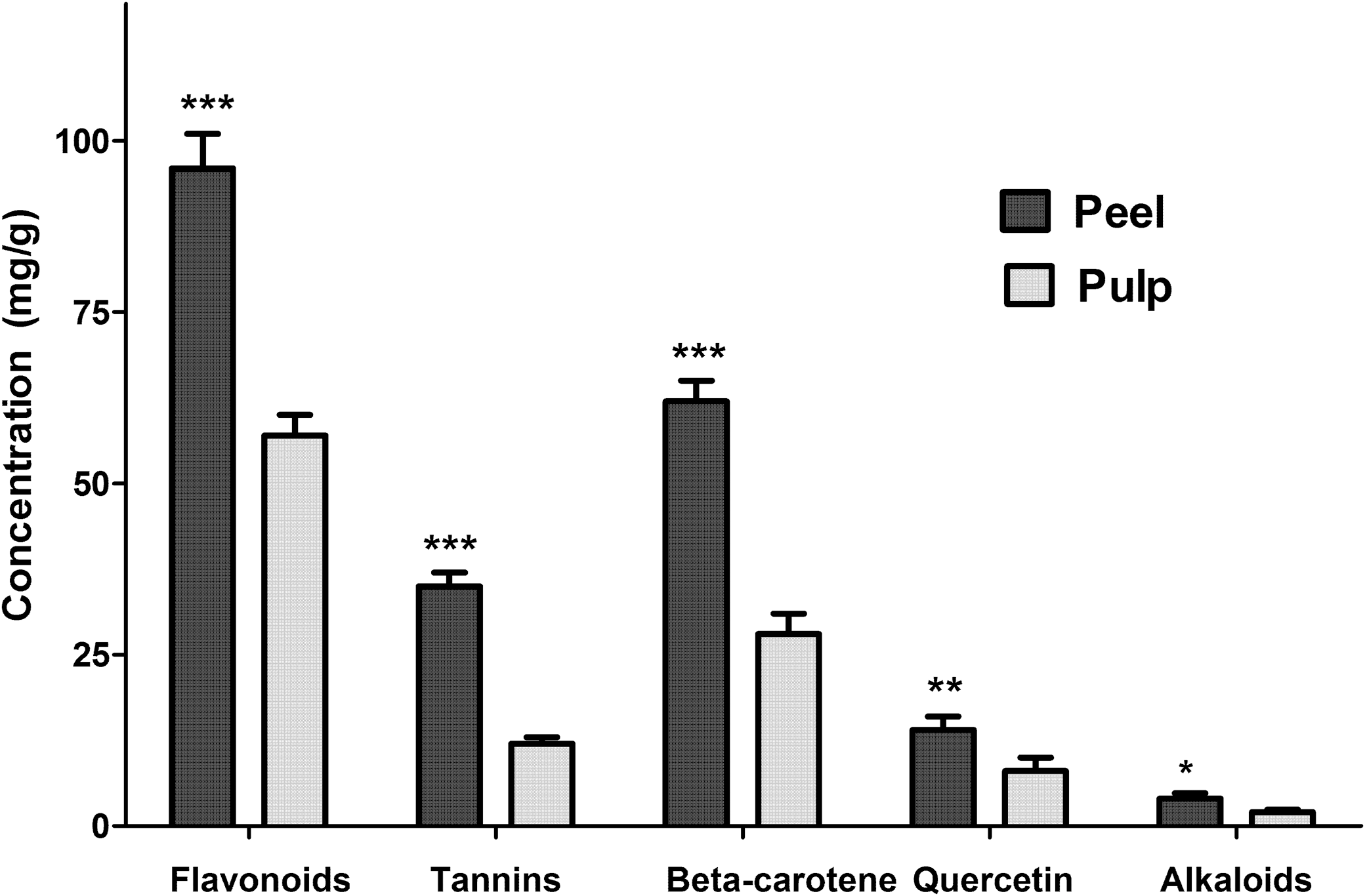
Bioactive compounds detected by spectrophotometry (total flavonoids, tannins, and alkaloids) and HPLC (β-carotene and quercetin) present in tucuma peel and pulp extracts compared by Student's t-test (*P<.05; **P<.01; ***P<.001). Graphic bars represent mean±standard error.
The first experiment evaluated the acute effect of tucuma extracts on PBMC DNA fragmentation, and the results are presented in Figure 2. The PBMCs treated with tucuma peel extracts presented a significant increase in DNA fluorescence when compared with an untreated group in 15, 30, and 60 min of treatment, indicating a genoprotective effect. However, the PBMC treated with tucuma pulp extract presented similar DNA denaturation observed in the control group except at a concentration of 10 μg/mL. In this concentration, the PBMCs fluorescence level was higher than the control group, indicating a genoprotective effect.

DNA denaturation of human peripheral blood mononuclear cells (PBMCs) after treatments with tucuma peel ), and expressed as μg/mL. Percentages of dsDNA were calculated in relation to the untreated control, and each of the symbols represents means±standard error of the mean (SEM) (n=8 wells) obtained in experiments. (*) Significant differences (P<.05) versus control (♦) evaluated by Measure Repeated analysis of variance followed by post hoc Tukey test.
The second experiment evaluated the potential genotoxic effect on PBMC exposed during 24 h to different concentrations of tucuma extracts. All PBMCs exposed to tucuma peel extracts presented higher DNA degradation than the control group (P=.0001). However, in PBMC exposed to tucuma pulp extracts, 51.4% of dsDNA degradation occurred in only 1000 μg/mL of pulp extract when compared with the control group, indicating some genotoxicity effect (P=.0001).
The results of genotoxicity induced by tucuma extracts using comet assay under alkaline conditions are described in Table 1. The damage index was higher in all concentrations of both extracts when compared with the PBMC control group (P=.0001).
Index damage: Σ (1,2,3,4 comet class)/100.
0, nucleus without DNA damage.
The potential effect of tucuma extract on chromosome instability was also evaluated, and the results are presented in Table 2 and Figure 3. The MI was influenced by the presence of tucuma extracts. In the lowest concentrations of both extracts, the MI was similar to the control group. However, from the 100 μg/mL concentration, the mitosis decreased dramatically (≤1%) in the peel (P=.001) and in the pulp (P=.001).
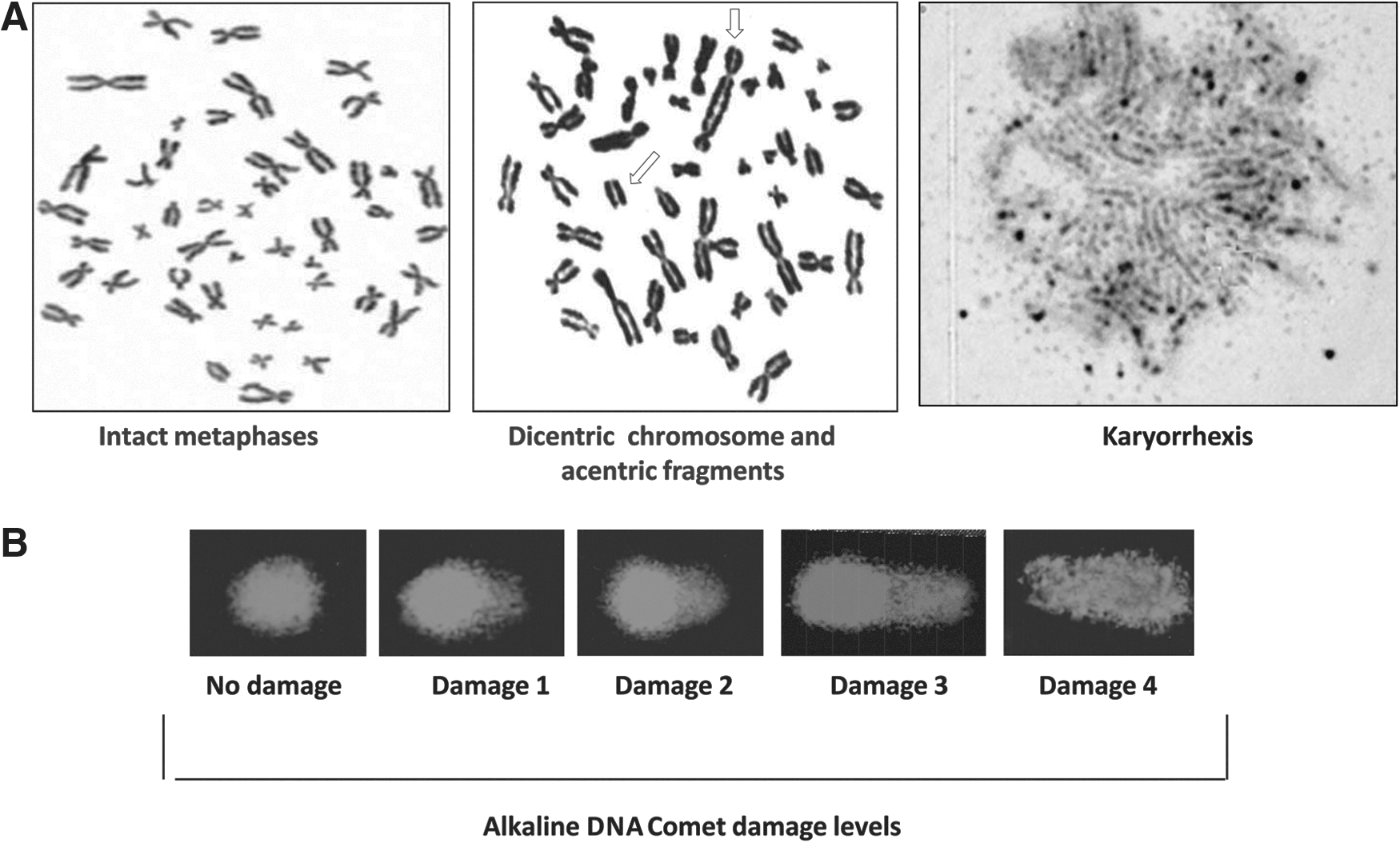
Microphotographs of genotoxicity assays:
n, number of nuclei counted in each treatment present in central region of glass slide.
Percentage of cells in the metaphase phase divided by total nuclei.
Metaphases with chromosomal damage.
The chromosome instability was not detected until 100 μg/mL of both extract's broken chromosomes could no longer be observed. The karyiopyknosis frequency was also lower until it reached a 10 μg/mL concentration in both extracts (<1.1%). However, karyiopyknosis increased in a concentration-dependent way from a 100 μg/mL concentration in both extracts. The Karyorrhexis/karyolysis was observed only in the higher concentrations (500 and 1000 μg/mL) of tucuma extracts.
We found significant differences in the MI among the cells treated with different tucuma extract concentrations.
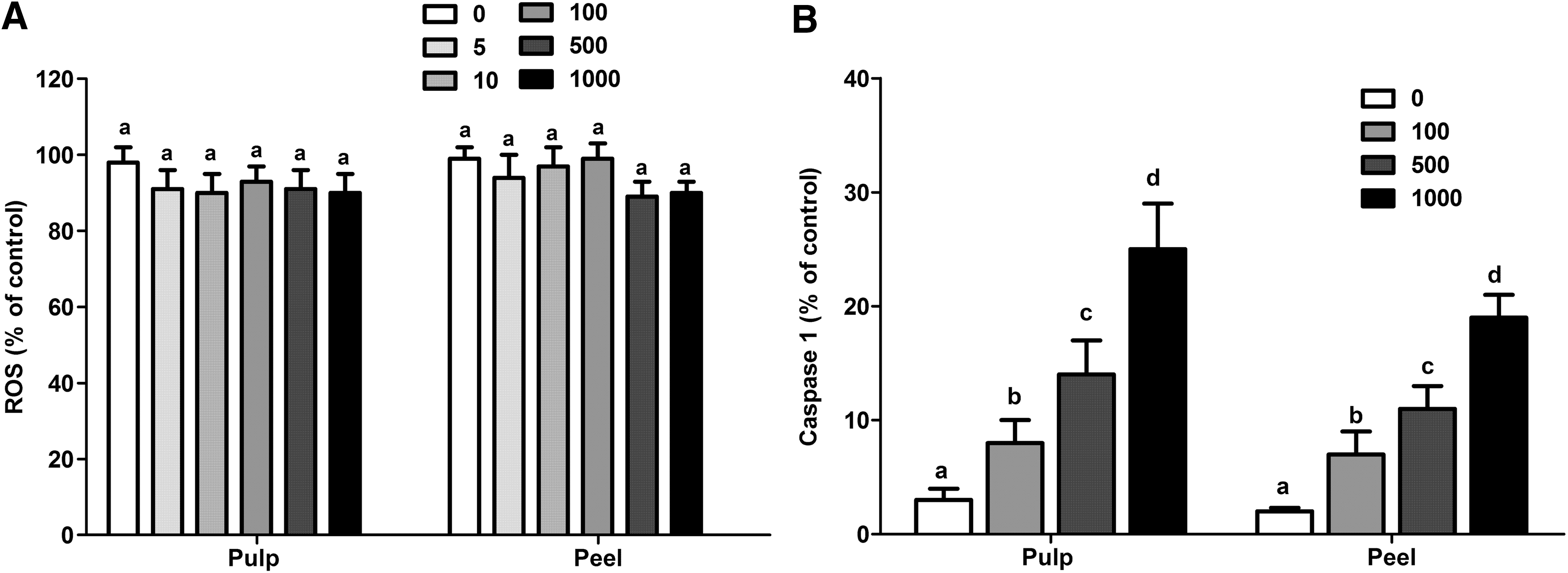
The ROS production for PBMCs culture exposed during 24 h to different concentrations of tucuma peel and pulp extracts were also examined. Despite the 500 and 1000 μg/mL concentrations, in both extracts, the trend to decrease 23–25% the ROS production was found. When compared with the control group (Fig. 4A), these differences were not statistically significant (peel extract P=.167; pulp extract P=.232).
Reactive oxygen species (ROS) levels evaluated by fluorimetric DCFH-DA assay
The caspase-1 activity was also evaluated among the different tucuma extract concentrations to observe the potential triggering of an apoptosis event, mainly pyroptosis in PBMC cells exposed to >100 μg/mL tucuma extract. As viewed in Figure 4B, the caspase-1 activity presented an increase in activity concentration dependence in both extracts (P=.0001). However, the peel extracts presented higher caspase-1 activation in 500 and 1000 μg/mL concentrations than the pulp extracts (P=.0001).
Discussion
The study described the potential genotoxicity effect of pulp and peel obtained from the tucuma Amazonian fruit that is very rich in carotenoids and other bioactive compounds. In the first experiment that evaluated the potential effect of tucuma extracts on DNA denaturation, a genoprotective action was observed mainly in tucuma peel extract. These results are most likely related to the high concentration of antioxidant compounds, mainly β-carotene, which is found in tucuma fruit. 8,9
However, with the other results obtained from PBMC treatments in which DNA fragmentation was analyzed, DNA cell damage evaluated by Comet assay and analysis of chromosomal instability suggested some genotoxic effect of tucuma extract in >500 μg/mg concentrations. Since we did not find differences in the ROS production among the control group and PBMC treated with tucuma extracts, we can hypothesize that the antioxidant compounds present in the extracts were metabolized by the cells in the first hours. In these terms, since the tucuma presented other molecules (tannins and alkaloids) for which we did not know their specificity, we could not discard the possibility that some could present toxic effects on PBMC cells. In addition, several investigations have shown the antioxidant, pro-oxidant, or both effects. 21
Why do such ambiguous results exist with regard to carotenoids? Authors such as Krinsky and Johnson 22 consider that the reason for this discrepancy may be attributed to the use of different methodologies (1) in dissolving the carotenoid to be evaluated, (2) in initiating oxidant stress, (3) in the presence of other antioxidants, (4) in the type of animal used for in vivo studies, and (5) in the evaluation technique used to determine the efficacy of the various carotenoids. Our results corroborate this suggestion, as we found different effects considering PBMC acute or chronic treatment of tucuma extracts.
Other important results are related to differences in the genotoxic response depending on tucuma concentrations. From our results, we observed that the lower tucuma extract concentrations (from 5 to 100 μg/mL) presented more positive effects than higher concentrations. The occurrence of U-shaped dose-response relationships (often termed hormesis) of several herbal products has been documented in numerous biological, toxicological, and pharmacological investigations. Many of the endpoints studied are of considerable significance to public health. 23
The potential differential effect of tucuma extract dose dependence is reinforced by results related to caspase-1 activity that increased in PBMC cultures treated with higher tucuma extract concentrations (≥500 μg/mL).
Cell death can occur via apoptosis, necrosis, and pyroptosis, distinct processes that present morphologic changes, the presence of DNA fragmentation, chromatin condensation, cellular lysis, and the release of cytosolic contents to the extracellular space. 24,25 The process is distinct from apoptosis and necrosis, although the pyroptosis is an inflammatory and a more rapid event than apoptosis. 24
The caspase-1 activation results in apoptosis via caspase-7 and is also a key molecule in the pyroptosis process. The higher caspase-1 activity in PBMC extracts treated with higher tucuma extracts indicated that these extracts are able to trigger cellular programmed death, specifically apoptosis and pyroptosis.
These results suggest that tucuma extracts could have some effect on cancer biology due to the properties which induce caspase-1 activation. This suggestion is corroborated in studies about the effect of retinoids as all-trans-retinoic acid (ATRA) that are present in tucuma extracts in cancer cells. Caspase activation is fundamental for retinoid-induced apoptosis, including caspase-1. 24 In promyelocytic leukemia cells, the growth inhibition, differentiation, and apoptosis are directly influenced by retinoids via activation of caspase, in particular, caspase-1. A previous study showed that bioactive compounds present in tucuma extracts in high doses suppress the growth of some cell lines such as cervical carcinoma by activation of caspase-1. 26
Therefore, caspase-1 can operate at the cell-autonomous level to eliminate malignant precursors through programmed cell death. On the other hand, in healthy cells, the activation of inflamed pathways may stimulate the production of trophic factors for cancer cells and their stroma. 27 Our results described here indicate that higher tucuma extract concentrations could not present beneficial effects on PBMCs, including routes related to inflammation and cell-programmed pathways.
Conversely, lower tucuma extract concentrations are safe and potentially present beneficial effects on human physiology that could contribute in the prevention of diseases.
Phytochemicals have great potential in disease prevention, mainly cancer, because of their safety, low cost, and oral bioavailability. There are numerous reports of cancer chemopreventive activity of dietary botanicals, including cruciferous vegetables such as cabbage and broccoli, as well as Allium vegetables such as garlic and onions, green tea, citrus fruits, soybeans, tomatoes, berries, and ginger, as well as medicinal plants. Several lead compounds, such as genistein (from soybeans), lycopene (from tomatoes), brassinin (from cruciferous vegetables), sulforaphane (from asparagus), indole-3-carbinol (from broccoli), and resveratrol (from grapes and peanuts), are in preclinical or clinical trials for cancer chemoprevention. 27
When compared with traditional vegetables such as carrots, tucuma extracts present higher carotenoids concentrations, including RAs that are a class of compounds structurally related to vitamin A. 2 These compounds presented antioxidant activity as well as other biological effects that regulate a wide range of biological processes, including embryonic development, differentiation, proliferation, and apoptosis. The RAs also participate in important physiological functions, primarily in the brain and the reproductive system. 28 Investigations have also observed that RAs can interfere with these events on several levels related to carcinogenesis. This is the case of ATRA molecules that provide essential support to diverse biological systems and physiological processes. 29
Differentiation therapy with ATRA has marked a major advance and become the first drug of choice in the treatment of cancers, such as acute promyelocytic leukemia. ATRA is being increasingly included in anti-tumor therapeutical schemes for the treatment of various tumoral diseases such as Kaposi's sarcoma, head and neck squamous cell carcinoma, ovarian carcinoma, bladder cancer, and neuroblastoma. The ATRA also presents antiangiogenic effects in several systems, inhibiting proliferation in vascular smooth muscle cells (VSMCs) and anti-inflammatory in rheumatoid arthritis. 30
Tucuma extracts also present other important polyphenols such as quercetin. Quercetin is the main flavonoid present in the human diet. Flavonoids are compounds occurring in plants that present important antioxidative properties.
Several therapeutic proprieties of flavonoids, mainly of quercetin, have been studied in the last decade, such as the antioxidant and anticarcinogenic potential and their protective effect to the renal, cardiovascular, and hepatic systems. Most flavonoids have some potentially positive effects on the cardiovascular system, although the final pharmacological targets differ markedly among various flavonoids. 31 Biochemical and genetic studies on cellular and animal models showed that quercetin presents a broad range of biological properties. Multiple lines of experimental evidence suggest a positive association between quercetin intake and improved outcomes of inflammatory cardiovascular risk. Quercetin can be considered the prototype of a naturally occurring chemopreventive agent because of its key roles in triggering the “hallmarks of cancer.” 32 In this context, tucuma extracts could affect the modulation of biological process related to cancer prevention and therapy.
However, our results also support the fact that the beneficial tucuma effects are dependent on the concentration and time of treatment, as these variables were related to an increase in the genotoxic effects observed in PBMCs. The genotoxic concentration-dependent effects have been previously described for other plants used by humans. For example, Wnuk et al. 33 described that yerba-mate (Ilex paranaguensis) extracts increased in the level of apoptotic and necrotic of lymphocytes in a dose-depending manner.
Finally, we need to consider some methodological limitations related to the study presented here. We only used in vitro protocols to analyze the potential genotoxic effect of tucuma extracts. Therefore, these results cannot be directly transferred to in vivo models, as with these models, we need to consider other intervenient variables such as bioactive compounds and bioavailability as well as the absorption and biological metabolism. For example, despite β-carotene being one of the major carotenoids present in our diet and in human blood and tissues, its bioavailability is dependent on a vegetable source. Other factors can be important in its bioavailability, such as cooking, chopping, and the presence of dietary fat. 34 Since the tucuma fruit is very rich in oil, this condition potentially increases the bioavailability of compounds such as carotenoids. 33 Therefore, complementary investigations using experimental models need to be performed to confirm the results described here.
Conclusion
In summary, tucuma extracts, despite their higher antioxidant content and antioxidant activity, would be present in the genotoxic effects of PBMCs that are dependent on concentration and time exposition. These results need to be considered in future in vitro and in vivo studies of tucuma effects.
Footnotes
Acknowledgments
This work was performed with financial support (grants and fellowships) from Brazilian research agencies: FAPEAM, FAPERGS, and CNPq. The authors thank Aline Augusti Boligon, Margareth Linde Athayde, and the Biogenomic Lab research team for technical support.
Author Disclosure Statement
No competing financial interests exist.