
Abstract
Clostridium difficile is the leading cause of hospital-acquired antibiotic-associated diarrhea worldwide; in addition, the proliferation of antibiotic-resistant C. difficile is becoming a significant problem. Virgin coconut oil (VCO) has been shown previously to have the antimicrobial activity. This study evaluates the lipid components of VCO for the control of C. difficile. VCO and its most active individual fatty acids were tested to evaluate their antimicrobial effect on C. difficile in vitro. The data indicate that exposure to lauric acid (C12) was the most inhibitory to growth (P<.001), as determined by a reduction in colony-forming units per milliliter. Capric acid (C10) and caprylic acid (C8) were inhibitory to growth, but to a lesser degree. VCO did not inhibit the growth of C. difficile; however, growth was inhibited when bacterial cells were exposed to 0.15–1.2% lipolyzed coconut oil. Transmission electron microscopy (TEM) showed the disruption of both the cell membrane and the cytoplasm of cells exposed to 2 mg/mL of lauric acid. Changes in bacterial cell membrane integrity were additionally confirmed for VCO and select fatty acids using Live/Dead staining. This study demonstrates the growth inhibition of C. difficile mediated by medium-chain fatty acids derived from VCO.
Introduction
C
Early studies suggest that medium-chain fatty acids (MCFA), commonly found in tropical oils such as coconut oil, are bactericidal for gram-positive bacteria, more so than gram-negative bacteria, fungi, protozoa, and viruses. 10 –13 These results are consistent with more recent reports of MCFA antimicrobial effects on group A streptococci, group B streptococci, Staphylococcus aureus, Neisseria gonorrhea, Chlamydia trachomatis, and Candida albicans. 14 –16 Further study reveals the sensitivity of ubiquitous species of Candida to virgin coconut oil (VCO) in vitro, 17 as well as antiviral effects of VCO when tested as a monotherapy for human immunodeficiency virus-infected patients in a clinical trial. 18
VCO is composed of greater than 99% triglycerides, with free fatty acids making up less than 0.2%. 19 Saturated fats account for 89% of Garden of Life VCO (used in this study), with polyunsaturated and monounsaturated fats reported as 7% and 4%, respectively, as noted by the manufacturer. Furthermore, lauric acid makes up ∼48% of the saturated fat found in coconut oil, with caprylic and capric accounting for 7% and 6%, respectively. 20 VCO triglycerides can be lipolyzed with lipase and water to form monoglycerides, diglycerides, glycerol, and free fatty acids. 21 Monoglycerides and free fatty acids have been reported to exhibit the antimicrobial activity, 16,22 –24 whereas triglycerides, diglycerides, and glycerol have been previously shown to have a lesser antimicrobial activity. 10
The exact mechanism(s) by which VCO exerts antimicrobial effects is/are still unknown. It has been suggested that VCO must be metabolized to release its component MCFAs, caprylic acid (C8), capric acid (C10), and lauric acid (C12) to exert its antimicrobial effects. 17 Of these metabolites, lauric acid may have the most antimicrobial activity. 10 A proposed mechanism for the antibacterial effects of VCO suggests that membrane lipids are solubilized as the VCO fatty acids integrate into the membrane. 10 Electron microscopy demonstrates cell membrane disruption of bacteria when exposed to VCO fatty acids in following microbes: Clostridium perfringens, C. albicans, C. trachomatis, Streptococcus agalactiae, and S. aureus, with subsequent lysis of bacteria exposed to VCO fatty acids. 12,16,22,25
C. difficile inhabits the small and large intestines in humans where the pH range varies from a high of 8.5 in the duodenum and a range of 6.4–7.5 in the colon. 26 Acidity or alkalinity directly affect the activity of monoglycerides and free fatty acids on microbes; therefore, we studied the lipids that would be most active at the pH range found in these regions of the intestinal tract. 12,27
While previous findings of antimicrobial activity offer insight into potential medical uses of VCO, to date, the susceptibility of C. difficile to VCO, lauric, capric, or caprylic acids, has not been studied. Accordingly, the objective of this study was to determine whether C. difficile is susceptible to the antimicrobial effects of VCO and/or its component fatty acids. Specifically, VCO, lauric (C12), capric (C10), and caprylic acid (C8) were tested against toxin A- and toxin B-positive C. difficile (ATCC 9689). Effects on C. difficile viability in culture, Live/Dead® staining, and electron microscopic evaluation of the cell membrane and cytoplasm following exposure are presented. Other long-chain fatty acids present in VCO (myristic and palmitic) were not studied because they have been previously shown not to be effective against Clostridium sp. at the pH range commonly found in the intestinal tract. 12
Materials And Methods
Culture media
Culture media was prepared using Becton Dickinson Brain Heart Infusion broth (BHIB; ref# 211059; Becton, Dickinson and Company, Sparks, MD, USA) following the manufacturer's directions. Ethyl alcohol (EtOH; Quantum Chemical Co., Newark, NJ, USA) was added to the BHIB so that the final volume was equal to 1% (v/v). Brain Heart Infusion agar plates were prepared following the manufacturer's directions. The pH of the broth cultures was determined using an Accumet pH meter (Fisher Scientific Co., Pittsburgh, PA, USA).
Virgin coconut oil
Pure 100% organic VCO (Garden of Life, West Palm Beach, FL, USA) was digested with porcine lipase (Cat# 096K0747; Sigma-Aldrich, Co. St. Louis, MO, USA) following the method of Dumitriu et al. 28 A reaction mixture containing 1.25 mL of VCO, 1.25 mL of sterile distilled water, 2 mL of phosphate-buffered saline (PBS) pH 7.4 and 200 mg porcine lipase. The mixture was continuously stirred for 30 min at 37°C. After incubation, the mixture was centrifuged (IEC Centra CL5R, rotor model IEC 6555C; Thermo Electron Corporation, Needham Heights, MA, USA) for 10 min at 4000 g to form three layers. Two hundred milliliters of the top clear layer (lipid) was solubilized in 400 mL of dimethyl sulfoxide (DMSO; Sigma-Aldrich) for a concentration of 33.3% (v/v) digested coconut oil. The efficiency of digestion was 23% by titration with 0.1 M KOH, resulting in the following concentrations of free fatty acids (in μM): caprylic acid, 420; capric acid, 940; lauric acid, 4050; myristic acid, 2540; palmitic acid, 1250; oleic acid, 510; and steric acid, 490. The digested VCO was then added to BHIB to provide final concentrations of digested coconut oil as 0.15% (v/v) to 1.2% (v/v) in BHIB.
Fatty acids
Lauric acid (Sigma-Aldrich) was solubilized by diluting 100 mg solid lauric acid in 10 mL DMSO to make a stock solution of 50 mM lauric acid in DMSO. The stock solution was further diluted into BHIB to make final concentrations of 1000, 500, and 250 μM lauric acid in BHIB with 2% DMSO.
Capric acid (Sigma-Aldrich) was treated similarly to lauric acid where 860 mg was placed in 5 mL of DMSO to make a stock solution of 500 mM capric acid in DMSO. The stock was then diluted into the BHIB for final concentrations of 2000, 1000, 500, and 200 μM capric acid in BHIB with 2% DMSO.
A stock solution of caprylic acid (Sigma-Aldrich) was prepared by adding 720 mg to 5 mL of DMSO for a concentration of 500 mM caprylic acid in DMSO. The stock solution was further diluted in BHIB to obtain final concentrations of 10,000, 5000, 2000, and 1000 μM caprylic acid in BHIB with 2% DMSO.
Live/Dead Bacterial Viability Kit (BacLight™)
Overnight cultures of C. difficile were diluted 1:50 into BHIB and grown in the presence or absence of solubilized fatty acids as described above. Cell viability was assessed using SYTO-9™ (Ex/Em 480/500 nm) and propidium iodide (PI; ex/em 490/635 nm), as per the manufacturer's directions (Live/Dead Bacterial Viability Kit, BacLight; Molecular Probes, Inc., Eugene, OR, USA). Briefly, 100 μL aliquots of well-mixed BHIB/1% ethanol inoculated with C. difficile, containing either 2% DMSO only or 2% DMSO plus solubilized fatty acids, were mixed with 30 μL of Live/Dead stain diluted to working concentration as per the manufacturer's instructions. Cell suspensions were vortexed for 5 sec, mounted on a slide, and immediately observed using an Olympus BX-61 microscope (Olympus America, Lake Success, NY, USA) adapted for fluorescence microscopy, using Olympus U-MNB2 and U-MNG2 filter sets for SYTO-9 and PI, respectively. Cells with intact membranes appear green using the U-MNB2 filter set, whereas those cells with compromised membranes appear red using the U-MNG2 filter set.
Fatty acids and C. difficile susceptibility assay
An overnight culture of C. difficile ATCC 9689 (American Type Culture Collection, Manassas, VA, USA) was grown in BHIB. One hundred microliters of bacteria was used to inoculate 9.9 mL of BHIB/1% EtOH containing varying amounts of lauric, capric, or caprylic fatty acids (dissolved in DMSO). The cultures in 15-mL disposable sterile centrifuge tubes (VWR International, Radnor, PA, USA) were incubated at 36°C in an anaerobic chamber (Bactron; Sheldon Manufacturing, Cornelius, OR, USA), being mixed by inversion every 2 min, for a total of 30 min. Following incubation, 100 μL samples were removed from the respective subcultures and serial dilutions were prepared in sterile water blanks and plated to BHI agar (BHIA). The culture plates were incubated overnight in the anaerobic chamber at 36°C and colony counts performed the next day to determine growth inhibition.
Controls for lauric, capric, and caprylic acids included: BHIB alone, BHIB 1% EtOH, and BHIB 1% EtOH+2% DMSO. C. difficile was added to the controls from an overnight culture to insure that no inhibition was caused by the addition of EtOH or DMSO.
Lipolyzed VCO and C. difficile susceptibility assay
The DMSO-solubilized lipase-digested coconut oil was added to the overnight cultures of C. difficile. The mixture was incubated for 30 min at 36°C and mixed by inversion every 2 min before diluting in sterile distilled water and plating onto BHIA. The newly inoculated BHIA was incubated overnight in an anaerobic chamber. Colony counts were performed on plates to determine growth inhibition.
Controls for the digested coconut oil were BHIB, and BHIB with DMSO/lipase/PBS mixture added, resulting in a 4% (v/v) final concentration of DMSO and a final lipase concentration of 6.7×10−7 g/mL. Controls testing for the undigested coconut oil included BHIB, and BHIB containing 4% DMSO.
Transmission electron microscopy
Lauric acid's effect on C. difficile was observed using transmission electron microscopy (TEM) following the methods of Skrivanová et al. 12 Briefly, overnight cultures of C. difficile ATCC 9689 were inoculated with either 2 mg/mL of lauric acid in DMSO or a control of DMSO only. The suspensions were continuously stirred for 30 min at 37°C. After incubation, the 1-mL aliquots of the suspension were centrifuged at 2500 g for 2 min (IEC Centra CL5R, rotor model IEC 6555C; Thermo Electron Corporation); the pellets were fixed with 500 μL of 5% glutaraldehyde (Fisher Scientific Co.) in 0.2 M Na-cacodylate buffer (pH 7.4; Sigma-Aldrich) and 500 μL of a solution of ruthenium red (0.15% in 0.2 M Na-cacodylate buffer; MP Biomedicals, LLC, Solon, OH, USA) for 1 h at room temperature.
The samples were again centrifuged at 2500 g for 2 min, and the pellets washed three times with 0.1 M Na-cacodylate buffer. The samples were fixed with 400 μL of 4% OsO4 (Stevens Metallurgical Company, New York, NY, USA) in 0.2 M Na-cacodylate buffer with 400 μL of ruthenium red (0.15% in 0.2 M Na-cacodylate buffer) for 4 h at 4°C. They were subsequently centrifuged at 2500 g and washed three times with 0.1 M Na-cacodylate buffer.
The fixed cells were suspended in molten 1% agar (Becton, Dickinson and Company), allowed to solidify at 37°C, and cut into 1-mm cubes. The cubes were then dehydrated by placing them in a graded series of ethanol in water. Once dehydrated with 100% ethanol (Quantum Chemical Co.), the samples were passed through propylene oxide (Sigma-Aldrich) by making a 1:1 solution with EtOH, followed by 100% propylene oxide. Agar cubes were placed in a series of dilutions of Embed 812 (Electron Microscopy Sciences, Hatfield, PA, USA) with propylene oxide until 100% of Embed 812 solution was achieved. The treated samples were placed in resin mold trays with 100% Embed 812 and hardened overnight in a 70°C oven (Fisher Isotemp Oven; Fisher Scientific Co.).
Sections of samples embedded in Embed 812 were cut at 100 μm thickness (Leica EMUC6 ultramicrotome; Leica, Vienna, Austria), placed on copper grids (Electron Microscopy Sciences), and stained with filtered (#1 Whatman filter paper; Fisher Scientific Co.) 2% aqueous uranyl acetate (Fisher Scientific Co.) for 15 min, followed by Reynold's lead citrate (Fisher Scientific Co.) for 2 min. The grids were rinsed in distilled water between each step and allowed to air dry. Sections were examined using a Jeol JEM 100-S transmission electron microscopy (Jeol, Peabody, MA, USA) and imaged with Kodak SO-163 electron image film (Electron Microscopy Sciences).
Statistical analysis
Statistical significance of the effects of VCO and select fatty acids on viable colony counts of C. difficile was determined using one-way analysis of variance (ANOVA) or linear regression, as appropriate, using the statistical software package R version 2.14.1 (2011). 29 Replicate data were used to generate arithmetic means and standard deviations for each treatment within an experiment. These group means were first compared using one-way ANOVA. Post hoc tests for significant differences between group means were then calculated using Tukey's honestly significant differences (HSD) method. If the ANOVA or graphs indicated a possible linear relationship, then linear regression, using the ordinary least-squares estimate, was used to model the relationship of the effects of VCO and select fatty acids on viable C. difficile colony counts. Significance was set a priori at P<.05.
Results
C. difficile susceptibility to lipolyzed VCO
VCO that was not predigested with lipase had no effect on the growth of C. difficile when added to overnight cultures (data not shown). VCO was therefore digested with lipase before its addition to C. difficile cultures. Exposure of C. difficile cells to VCO or its constituent fatty acids was evaluated to assess the effect of these lipids on cell growth. Lipolyzed VCO at a concentration of 1.2% inhibited 99.9% of C. difficile growth, while 0.15% VCO inhibited nearly 50% of bacterial growth (Fig. 1). BHIB containing 1.2% VCO had a pH of 6.9. Controls containing DMSO, lipase, and PBS were not significantly different from the BHIB alone in inhibiting the growth of C. difficile. ANOVA results indicated significant differences in group means. Post hoc comparisons of group means demonstrated that all doses of VCO inhibited bacterial growth. The logarithmic growth of C. difficile plotted as the independent variable with respect to the concentration of digested VCO indicates a log-linear relationship and is amenable to simple linear regression. Using a least-squares estimate, it was determined that for every 0.1% increase in the concentration of digested VCO there will be a reduction of ln(7.61) colony counts of C. difficile (P<.0001). The log-linear nature of this relationship means that coconut oil has a diminishing effect on colony counts of C. difficile. For example, the reduction in colony counts between 0.1% and 0.2% concentrations of digested coconut oil is 163,031 colony-forming units (CFU)/mL, whereas the reduction in colony counts between 0.3% and 0.4% concentrations is 35,553 CFU/mL. This log-linear relationship is remarkably strong with a multiple R 2 of 0.97.
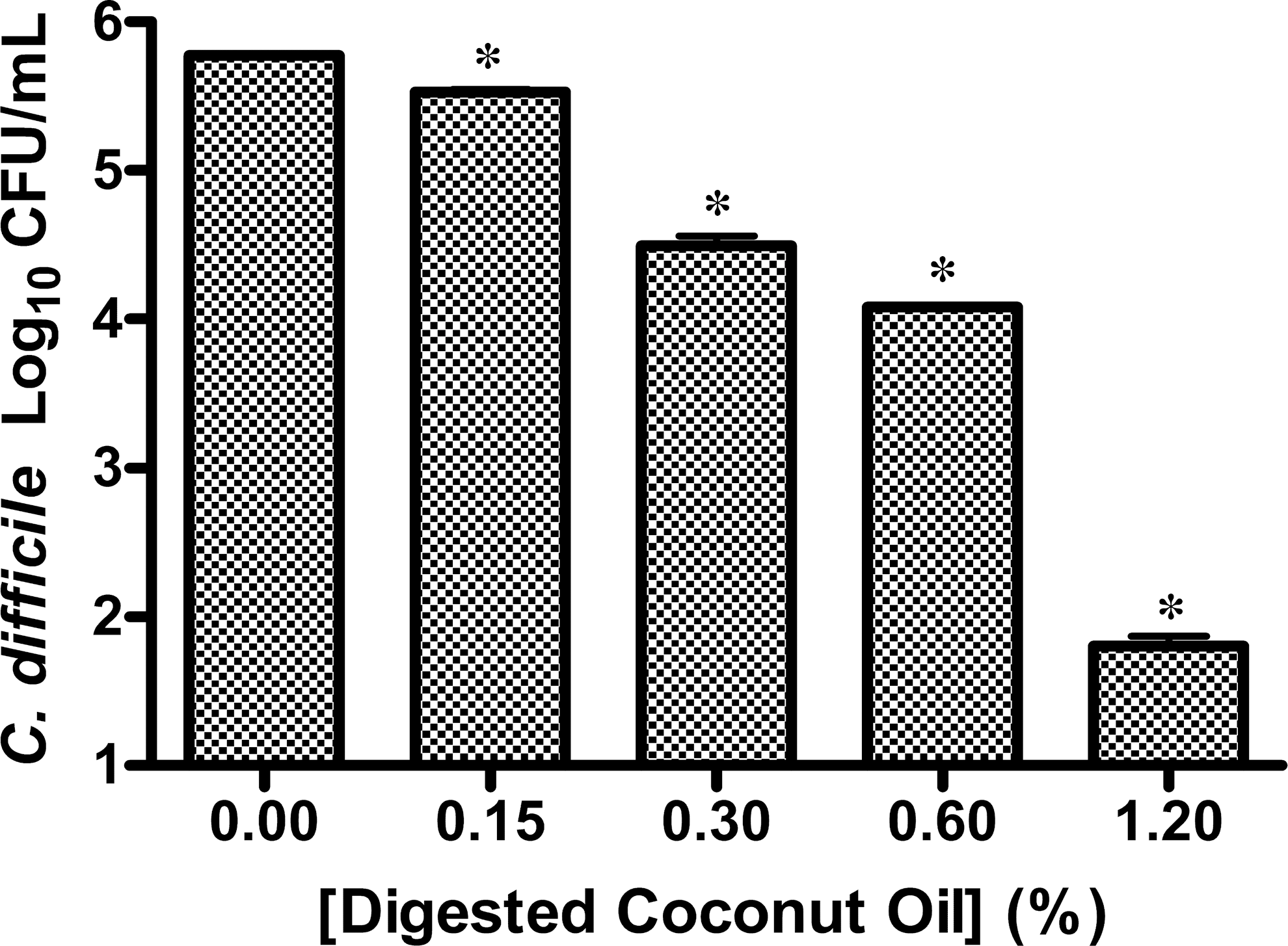
Colony counts of an overnight culture of Clostridium difficile ATCC 9689 exposed to coconut oil digested with porcine lipase for 30 min and plated to Brain Heart Infusion agar (BHIA; n=3). Controls: one with untreated sample, and one with 4% dimethyl sulfoxide (DMSO), lipase, and phosphate-buffered saline buffer. (*P<.0001).
C. difficile susceptibility to fatty acids
Lauric acid
Lauric acid inhibited nearly 100% of bacterial growth at the 1000-μM concentration (Fig. 2). Lauric acid produced strong inhibition even at concentrations as low as 250 μM, reducing growth by 90% compared to controls (Fig. 2). There was no inhibition of C. difficile growth in any of the controls (BHIB, BHIB containing 1% ethanol, BHIB containing 1% ethanol and 2% DMSO). Therefore, only the BHIB containing 2% DMSO and 1% EtOH control result is reported in the figures defining the fatty acid effect on C. difficile growth. The pH of the broth at 1000 μM of lauric acid was 7.0. Data evaluating the effect of lauric acid also demonstrated a strong log-linear relationship to C. difficile, similar to that of digested VCO. The results of simple linear regression show that for every 100 μM increase in the concentration of lauric acid there will be a reduction of ln(0.62) colony counts of C. difficile, which is significant (P<.0001). The log-linear relationship is also remarkably strong, with a multiple R 2 of 0.94.
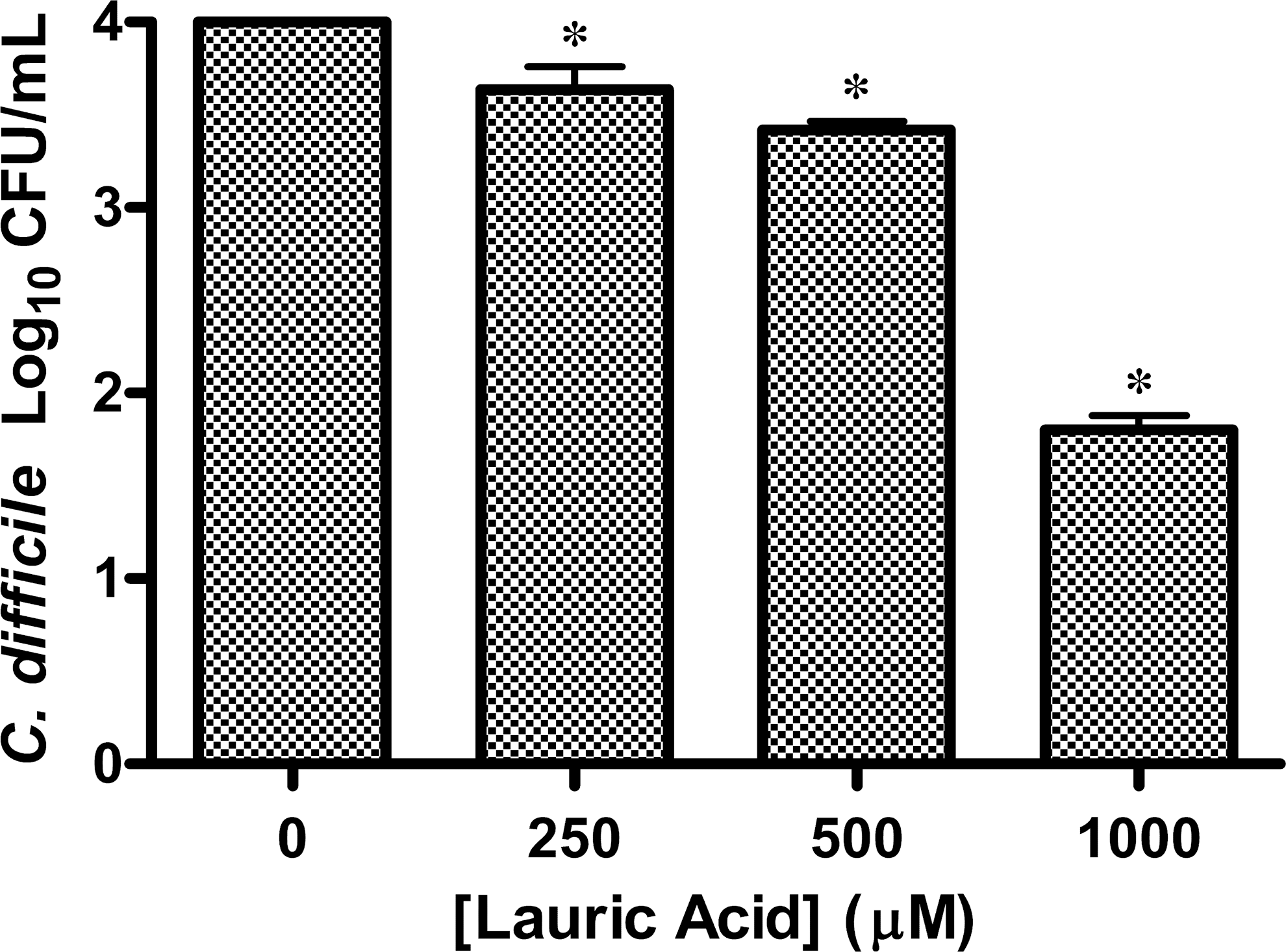
Colony counts of an overnight broth culture of C. difficile ATCC 9689 exposed to lauric acid for 30 min and plated to BHIA (n=3). The control (n=3) contained only Brain Heart Infusion broth (BHIB) with 2% DMSO and 1% ethyl alcohol (EtOH) with no lauric acid. (*P<.0001).
Capric acid
Capric acid (2000 μM) inhibited the growth of bacteria, whereas 1000 μM did not significantly affect the growth of C. difficile (Fig. 3). The pH of broth cultures containing 2000 μM concentration was 6.8. Capric acid at concentrations from 2000 to 200 μM/mL demonstrated a fairly linear relationship with colony counts of C. difficile. Simple linear regression showed that for every 1000 μM increase in the concentration of capric acid, there was a statically significant decrease in 2928 CFU/mL of C. difficile (P<.001). Evaluation by linear regression resulted in an R 2 value of 0.65.
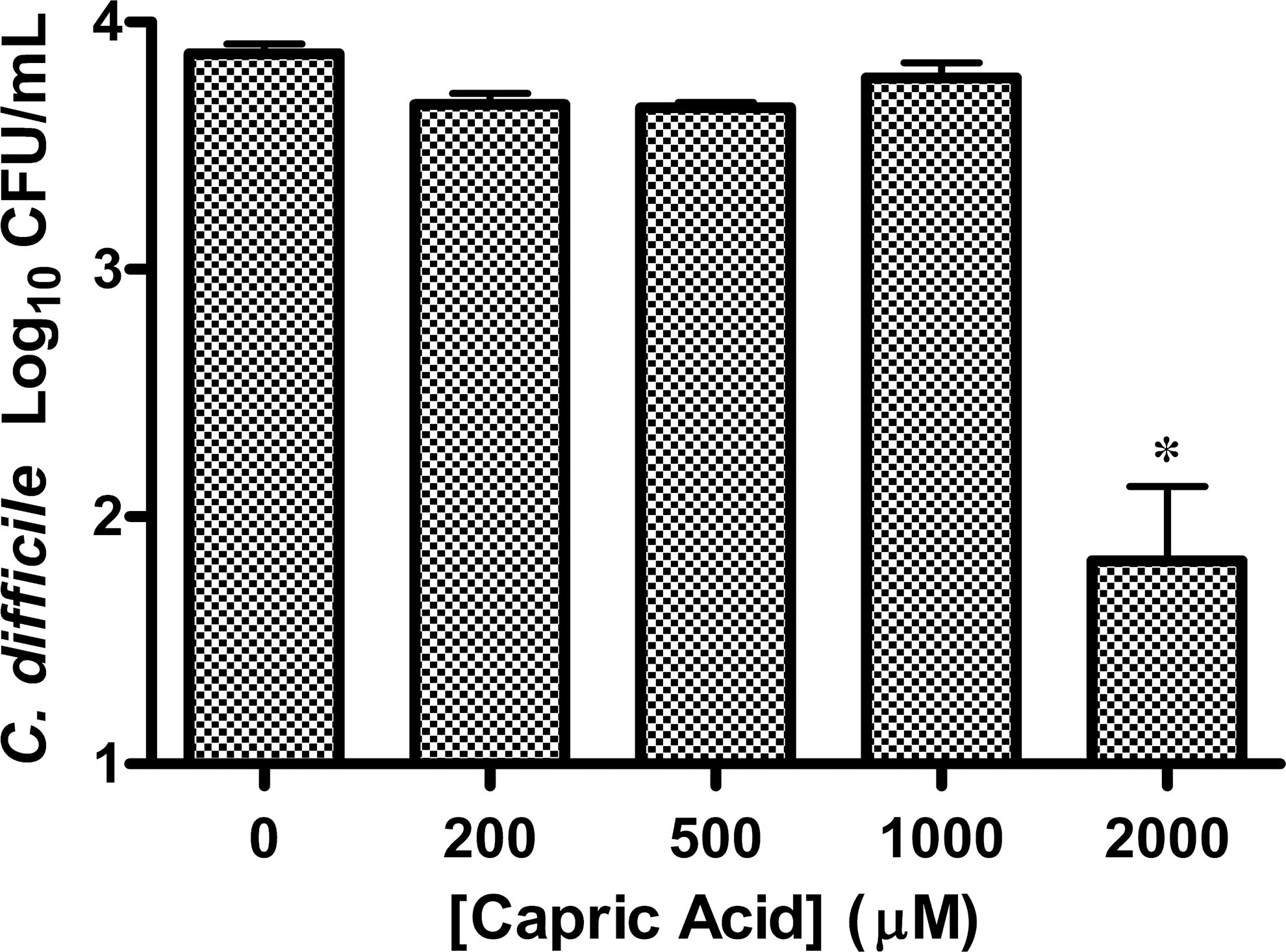
Colony counts of an overnight broth culture of C. difficile ATCC 9689 exposed to capric acid for 30 min and plated to BHIA (n=3). The control (n=3) contained only BHIB with 2% DMSO and 1% EtOH with no capric acid. (R 2=0.65; *P<.001).
Caprylic acid
Caprylic acid inhibited the growth of C. difficile, but its inhibitory effect was blunted when diluted twofold (5000 μM). Further dilution of caprylic acid also did not inhibit growth (Fig. 4). Caprylic acid at concentrations from 10,000 to 1000 μM/mL demonstrated neither a linear or log-linear relationship on the colony counts of C. difficile. The concentrations of caprylic acid were treated as categorical and a one-way ANOVA with multiple comparisons of the concentrations using Tukey's HSD showed that only when the concentration of caprylic acid reached 10,000 μM was there a significant decrease in colony counts of C. difficile (P<.001).
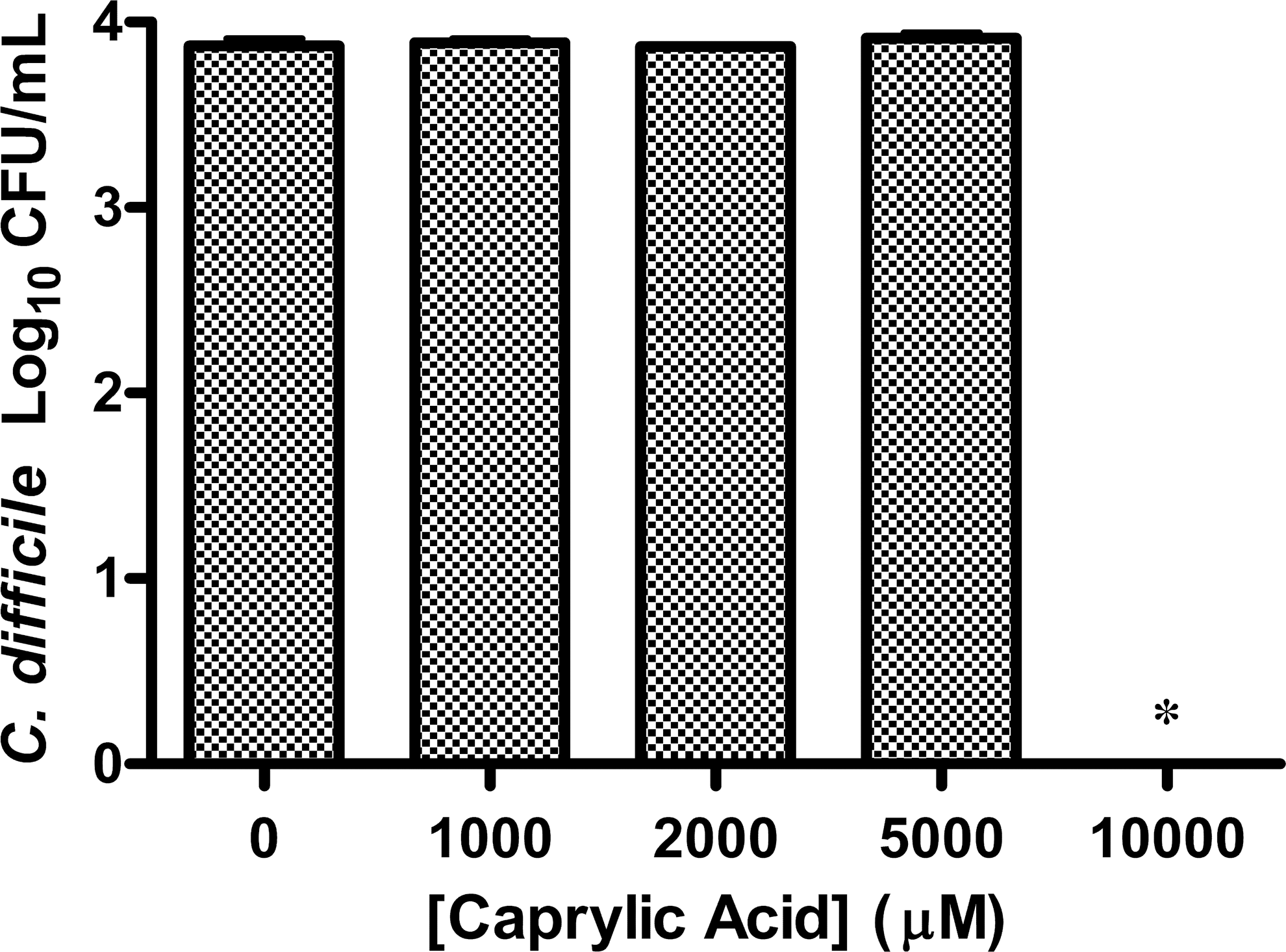
Colony counts of an overnight broth culture of C. difficile ATCC 9689 exposed to caprylic acid for 30 min and plated to BHIA (n=3). The control (n=3) contained only BHIB with 2% DMSO and 1% EtOH with no caprylic acid. (*P<.001).
Transmission electron microscopy
TEM of lauric acid-treated C. difficile demonstrated 100% alteration of cell ultrastructure compared to untreated control TEMs (Fig. 5). The cell membrane of the lauric acid-treated bacteria appeared to be eliminated; the lipid layers were not discernible, whereas the cytoplasm appeared to be disorganized with possible vacuoles, compared to the untreated control.
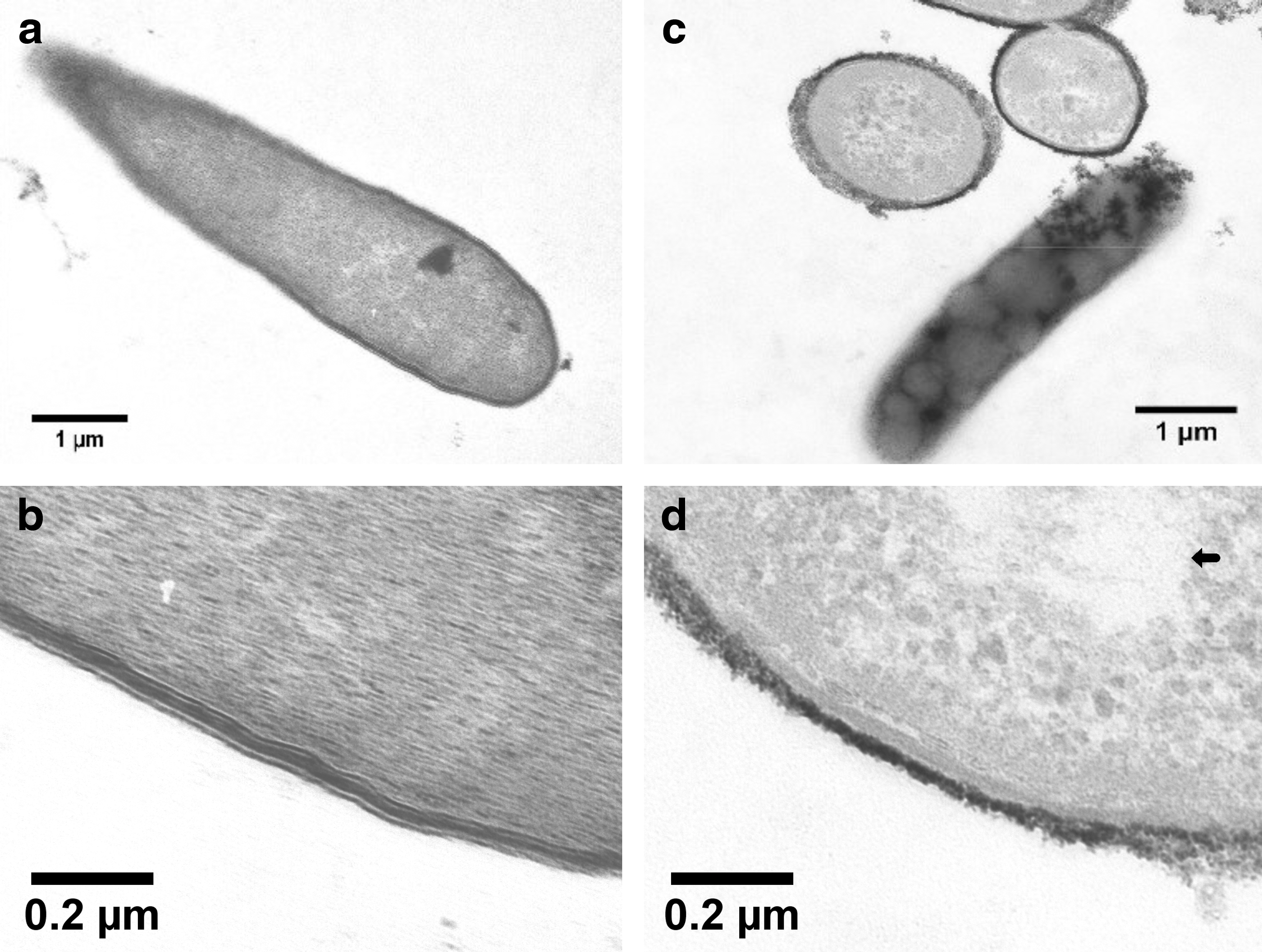
Transmission electron microscopy images of samples of C. difficile treated for 30 min with
BacLight viability testing (Live/Dead staining)
Live/Dead staining performed on bacterial aliquots from controls with DMSO only and treatments with solubilized fatty acids or lipolyzed coconut oil solubilized in DMSO demonstrated a change in membrane permeability as measured by PI uptake by bacteria in samples exposed to fatty acids. Lipolyzed VCO (1.2%), lauric acid (1 mM), capric acid (2 mM), and caprylic acid (10 mM) inhibited the growth of C. difficile by greater than 2 logs and caused nearly all bacteria to convert immediately from green (live) to red (dead). Control treatments (containing DMSO) only caused a few (less than one cell per high power field) to convert from green to red, even after 30 min at room temperature (data not shown).
Discussion
Coconut oil is a natural substance and is used in many tropical countries as the preferred oil for eating and cooking. Coconut oil is not regulated by the FDA and is consumed freely. Animal mucosal cells are not adversely affected by low levels of MCFAs. 16 However, controversy continues to question the safety of consuming of saturated MCFAs such as those contained in coconut oil, in light of the connection between fat consumption and heart disease via increasing cholesterol levels. 30 –32
VCO and several of its constituent fatty acids: lauric, capric, and caprylic acid have been shown in other studies to inhibit the growth of microbes, including enveloped viruses, yeast (e.g., C. albicans), and bacteria (e.g., C. perfringens). 11,12,14 –16,18,23,33 VCO, when consumed, would be digested by lipases of the digestive tract, releasing the individual fatty acids and ultimately inhibiting the growth of microorganisms in vivo. 34 C. difficile lacks lipases to digest coconut oil, 35,36 requiring the oil to be lipolyzed with porcine lipase before evaluating its effect in vitro. Results from this study demonstrate the ability of both the constituent fatty acids and lipolyzed VCO to effectively inhibit the growth of C. difficile in vitro.
The examination of fatty acid and lipolyzed coconut oil treated C. difficile using transmission electron microscopy and Live/Dead staining indicated that the cell membrane is disrupted. We hypothesize that disrupting the cell membrane with these MCFA leads to cell death by changing the permeability to water and other ions.
Our research found that nonlipolyzed VCO resulted in no inhibition of growth of C. difficile (data not shown). Bacterial growth was not inhibited by lipase, PBS, or DMSO. Lipolyzed VCO significantly inhibited the number of colony forming units of C. difficile in vitro at levels as low as 0.15% v/v. These results suggest that human consumption of VCO might inhibit the growth of C. difficile within the gastrointestinal tract.
Lauric acid is the largest component of VCO and, in our study, the most effective inhibitor of C. difficile. The strong similarities in the log-linear relationships of the effects of both lipolyzed coconut oil and lauric acid, together with the fact that lauric acid makes up ∼50% of VCO, suggest that lauric acid may be the primary active ingredient. 37 Capric acid and caprylic acid, which comprise between 5% and 10% of VCO, 37 were also found to inhibit the growth of C. difficile in vitro at concentrations greater than those of lauric acid. Twice as much capric acid (compared to lauric acid) was needed to achieve a 99% inhibition of the growth of C. difficile; similarly, caprylic acid was much less effective than lauric acid, requiring five times as much to achieve the same level of inhibition of growth as lauric acid. Our results suggest that both capric acid and caprylic acid also play a role in VCO's effect on C. difficile bacterial inhibition, but to a much lesser extent than that of lauric acid.
Based on the amount of lauric acid found in breast milk (6.2% of total fat) 38 and the nutritional requirements of infants, an adult would need to consume ∼24 g of fat containing lauric acid, to achieve the same systemic antiviral and antimicrobial inhibition of growth. 18,39 Lauric acid constitutes ∼48–52% of the total fatty acid content of coconut oil, suggesting that ∼52 g of coconut oil per day would be necessary to have systemic antimicrobial activity, similar to that observed in infants consuming breast milk. 18,39 However, the amount of coconut oil required to prevent intestinal colonization of pathogenic microbes may differ in vivo and further research, using an animal model, is warranted.
Efforts are ongoing to identify more effective alternate treatments for CDI. The VCO and lauric acid results outlined in this study provide support for VCO as a potential effective and economical therapy against CDI, especially by financially challenged patients. Since the proposed pathophysiology for the development of C. difficile diarrhea is bacterial overgrowth and alteration of the normal gut microbiota, future studies should include the study of the effect of VCO against cultures of members of the normal microbiota of the gastrointestinal tract and other intestinal pathogens to ensure that ingesting coconut oil will not have harmful effects on the normal GI tract microbiota and will have selective toxicity toward pathogens.
With further study, there may also be potential for coconut oil to be consumed as a prophylactic measure against CDI for those patients requiring long-term hospitalization, extended stays in rehabilitation, or skilled nursing facilities. Such a measure may serve to decrease colonization in patients and subsequently lead to fewer cases of hospital-acquired CDI.
The threat of CDI is a major health concern when caring for patients in health-care facilities. Alternate methods to control C. difficile, such as hand-washing and contact isolation, should continue to be used to prevent its spread. The use of alternative therapeutic agents may reduce health-care providers' dependence on and the growing resistance to metronidazole, as well costs associated with CDI. This in vitro study presents a limited analysis. Thus, a more complete evaluation of VCO and its constituent components should be undertaken to study their impact on C. difficile in vivo.
Footnotes
Acknowledgments
Financial support was provided by the College of Public Health, Kent State University. The authors acknowledge Jeanette Killius, Northeastern Ohio College of Medicine, for assistance with electron microscopy.
Author Disclosure Statement
No competing financial interests exist.