
Abstract
The effect of pomegranate juice (PJ) on hepatic antioxidant enzyme activities, lipid peroxidation, DNA fragmentation (DNAF), and caspase-3 activity in rats both treated and not treated with diethylnitrosamine (DEN) and phenobarbital (PB) was studied. Administration of rats with DEN and PB caused an elevation in the levels of malondialde-hyde (MDA), DNAF, and activities of glutathione reductase (GSR) and caspase-3, while the activities of superoxide dismutase, glutathione S-transferase, total glutathione peroxidase (t-GPx), and glutathione (GSH) level were decreased in hepatocytes compared to the control. Treatment of rats with PJ pre, during, and post DEN and PB administration improved liver function and decreased the levels of MDA, DNAF, t-GPx, GSR, and caspase-3 activities, but the GSH level did not change compared to the D–P group. This indicates that PJ reduced the oxidative stress and apoptosis induced by DEN and PB. Administration of healthy rats with PJ only for a long period induced oxidative stress and apoptosis for hepatocytes.
Introduction
T
Nitrosamines occur in latex products such as balloons and in many foods and other consumables. 3 –5 They are used in the manufacture of some cosmetics, pesticides, and in most rubber products. Diethylnitrosamine (DEN) has been found to be widely distributed in processed meats; tobacco smoke; whisky; smoked, salted, and dried fish; cheese; cured meat; and alcoholic beverages. 6,7 In foods, nitrosamines are produced from nitrites and secondary amines, which often occur in the form of proteins. Their formation can only occur under certain conditions, including strongly acidic conditions such as that of the human stomach. High temperatures, as in frying, can also enhance the formation of nitrosamines. Under acidic conditions, the nitrite forms nitrous acid (HNO2), which is protonated and splits into the nitrosonium cation N≡O+ and water: H2NO2 +=H2O+NO+. The nitrosonium cation then reacts with an amine to produce nitrosamine. These processes lead to significant levels of nitrosamines in many foodstuffs, especially beer, fish, and fish byproducts, and also in meat and cheese products preserved with nitrite pickling salt. There are rules about adding ascorbic acid or related compounds to meat in order to inhibit nitrosamine formation. 8,9 In the human body, nitrosamine can be produced from the metabolism of certain therapeutic drugs or from the reaction of nitrite with amines and amides. 3 DEN causes a variety of hepatocellular injuries, including necrosis, cirrhosis, hypertrophy, and hepatocellular carcinoma. 3 In general N-nitrosamines have been suggested to cause oxidative stress and cellular injury due to involvement of free radicals. 3
Phenobarbital (PB) is a derivative of barbituric acid. It is the most widely used anticonvulsant worldwide and the oldest still commonly used. It is often prescribed as a sedative or antiepileptic drug. 10 Sedation and hypnosis are the principal side effects of PB. Central nervous system effects, such as dizziness, nystagmus, and ataxia, are also common. Kitano et al. 11 reported that PB is a nongenotoxic hepatocarcinogen, and its administration at a high dose (500 ppm) after DEN injection promotes hepatocarcinogenesis in a rat model. This occurs due to its ability to induce the expression of the drug-metabolizing enzymes such as CYP2E1 and CYP1A2 isoenzymes, which are involved in the activation of DEN and PB. 12
A combination of epidemiologic and basic scientific evidence strongly suggests that diet and plant-derived phytochemicals may play an important role in cancer prevention or treatment. 13 Fresh and processed fruits and food products contain high levels of a diverse range of phytochemicals of which polyphenols, including tannins, anthocyanins, and other flavonoids, make up a large proportion. Phytochemical products have been shown to inhibit liver toxicology and cancer cell growth through the modulation of genes that are related to the cell proliferation, cell cycle, apoptosis, signal transduction, oncogenesis, and transcription regulation also possessing both anti-inflammatory and antioxidant effects. 14
Pomegranate (Punica granatum L.) is one of the oldest edible fruits and has been used extensively in the folk medicine of many cultures. Pomegranate fruits are widely consumed fresh and in beverage forms as juice and wines. 15 Commercial pomegranate juice (PJ) shows potent antioxidant, antatherosclerotic, and cardioprotective properties attributed to its high content of polyphenols. 16,17 PJ has been shown previously to exhibit in vitro and in vivo anticarcinogenic properties, such as induction of cell cycle arrest and apoptosis, as well as the inhibition of tumor formation and growth in animals. 13 Pomegranate fruit extracts possess polyphenolic and other compounds with antiproliferative, pro-apoptotic, and anti-inflammatory effects in prostate, lung, and other cancers. 18 Recently, there has been a report on the potent antioxidant activity of the whole fruit extract of pomegranate. 19 In the light of these findings, the aim of the present study was to evaluate (1) the ability of PJ to reduce rat liver injury induced by DEN and PB, and (2) the effect of PJ administration for a long period on healthy rats in order to establish whether PJ has a toxic effect.
Materials and Methods
Chemicals
A caspase-3 assay kit was obtained from BioSource International, Inc. (Camarillo, CA, USA). An AxyPrep DNA gel extraction and purification kit was obtained from Montreal Biotechnologies Inc. (Dorval, PQ, Canada). D, P, cumene-H2O2, 5,5′-dithiobis(2-nitrobenzoic acid), DTNB, reduced glutathione (GSH), oxidized glutathione (GSSG), nicotinamide adenine dinucleotide phosphate (NADPH), standard superoxide dismutase (SOD), thiobarbituric acid (TBA), and tetramethoxypropan (TMP) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Diethylene triaminopentaacetic acid (DTPA) and sodium dodecyle sulfate (SDS) were purchased from Fluka (St. Gallen, Switzerland).
Pomegranate juice
Pomegranate from the family Lythraceae was purchased from the local market. The fruits were peeled mechanically. Then the seeds of the fruit containing the intact juice sacs were manually separated and filtered, and the filtrate was stored at 4°C until used. 20
Characterizations of pomegranate extract
Total phenolic content
The total phenolic content was determined as mg gallic acid equivalent according to the method of Taga et al., 21 using gallic acid as standard.
High-performance liquid chromatography analysis for phenolic compounds in pomegranate juice
Chromatographic separation of 20 μL of PJ were carried out on 150 mm×4.6 mm, 5 μm Eclipse XDB–C18 column (1200S Series; Agilent Technologies, Waldbronn, Germany) at 320 nm and at a flow rate of 0.75 mL/min. The mobile phase was formic acid (1%, v/v in aqueous solution):acetonitrile:2-propanol (70:22:8), pH 2.5.
Animals
Twenty-eight adult male Sprague Dawley rats weighing 100–110 g were obtained from the Experimental Animals Breeding Center of the Holding Company for Biological Products and Vaccines (Helwan, Cairo, Egypt). All rats were examined for health status, and their room was designed to maintain a constant temperature of 25°C, relative humidity of approximately 50%, and a 12 h light/dark photoperiod for two weeks prior to handling. The animals were then housed in stainless steel cages, given a standard diet and water ad libitum throughout the study, and observed daily for abnormal signs. After acclimatization, rats were divided into four groups of seven rats each. Group C was the control group and comprised untreated rats. Group D–P rats were injected subcutaneously (s.c.) with 200 mg of DEN in saline/kg body mass (bm) each week for three successive weeks. 22 At the beginning of the fifth week, the animals were given drinking water containing 500 ppm of PB for 19 weeks 22,23 (the total experimental period is 23 weeks). Group (D–P)–PJ rats were treated orally (using oral gavages) with a daily dose 1 mL PJ/kg bm for 4 weeks. At the beginning of the fifth week, rats were treated with DEN (as described previously) in addition to the PJ treatment. At the beginning of the ninth week, rats were treated with PB with continuous intake of PJ until the 23rd week of the experiment (i.e., rats were treated with PJ pre, during, and post DEN and PB treatment). Group PJ rats were administered with a daily oral dose of 1 mL PJ/kg bm for 23 weeks. At the end of the experimental period, feeding was stopped 12 h prior to killing. Rats were anaesthetized by diethylether, and their livers were removed immediately. Small portions of livers were fixed in 10% formalin for histopathologic examination. The remaining liver tissues were washed with cold saline solution (0.9% NaCl), weighed, divided into two parts, and kept at −80°C until analysis. The first part was used for determination of DNAF, caspase-3, GPx, and GSR activities. The second part was homogenized in five volumes of homogenizing buffer (0.1 M sodium phosphate buffer containing 0.9% NaCl, pH 7.4) using a glass Teflon homogenizer. The liver homogenate was centrifuged at 3200 g for 20 min at 4°C, and the supernatant was divided into five portions and kept at −80°C for subsequent determination of lipid peroxidation, protein content, GSH, glutathione-S-transferase (GST), and SOD activities. All processes were carried out at 4°C.
Unheparinized blood samples were collected, kept for 15 min at room temperature, and the sera then were separated by centrifugation at 1000 g for 20 min and stored at −20°C until used for the determination of alanine aminotransaminase (ALT), aspartate aminotransferase (AST), total bilirubin, total proteins, and albumin.
Biochemical assays
Caspase-3 assay (EC 3.4.22.56)
Caspase-3 activity was determined using a colorimetric kit according to the method of Talanian et al. 24 Liver tissues were homogenized in four volumes of cold cell lysis buffer (50 mM Tris-HCl buffer containing 0.2 M NaCl and 1% Triton X-100, pH 6.8) using a Teflon glass homogenizer. The homogenates were centrifuged at 44,720 g for 3 min at 4°C, and the supernatants were kept at −80°C. The supernatant (50 μL=150 μg protein) was put in a microplate reader, then 50 μL reaction buffer and 5 μL of 4 mM substrate were added, mixed well, and incubated at 37°C in the dark for 2 h. The reaction rate was determined by measuring the absorbance of the produced yellow color at 405 nm against a blank using a microplate reader (Bio-Tek Instruments, Bad Friedrichshall, Germany). Fold increase in caspase-3 activity should be determined by direct comparison to the level of the control.
DNA fragmentation
DNAF was determined in the liver homogenate using agarose gel electrophoresis according to the method of Li et al. 25 Liver tissues were homogenized in (1:5 w/v) 50 mM Tris-HCl buffer containing 20% sucrose and 50 mM EDTA, pH 7.6. DNA was isolated using a DNA purification kit. Then 15 μg/lane DNA was separated by electrophoresis on 1% agarose gel containing 25 μL ethidium bromide at 5 V/1 cm for 2–3 h, and visualized under UV light using a multiband transilluminator from Consort (Turnhout, Belgium).
Lipid peroxidation
Lipid peroxidation was determined calorimetrically by measuring the level of MDA, the end product of lipid peroxidation, according to the method of Ohkawa et al. 26 Fifty microliters of the crude homogenate or homogenizing buffer (blank) were incubated with 100 μL of 8.1% of SDS, 750 μL of 20% acetic acid containing HCl, pH 3.5, 750 μL of 0.8% TBA, and 300 μL of distilled water in boiling water bath for 45 min. After cooling at room temperature, 500 μL of distilled water and 2.5 mL of n-butanol/pyridine mixture (15:1 v/v) were added, mixed well, and centrifuged for 10 min at 1780 g. The absorbance of the pink color was measured at 532 nm and the concentration of MDA was determined as nmol/g liver. Different concentrations of TMP (20–300 nmol) were used as standard and treated in a similar way as the sample.
Reduced glutathione
GSH was determined as described by Ellman. 27 The proteins in 0.5 mL of the liver supernatant were precipitated using 2 mL of 5% trichloroacetic acid (TCA), then centrifuged at 1780 g for 15 min. One milliliter of the deproteinized supernatant was added to 500 μL of Ellman's reagent (0.0198% DTNB in 1% sodium citrate) and 3 mL of 0.2 M phosphate buffer, pH 8. The mixture was incubated at room temperature for 10 min, and then the absorbance of the developed yellow color was read against a blank at 412 nm. GSH is expressed as mg/g tissue.
Total glutathione peroxidase (EC 1.11.1.19)
Liver tissues were homogenized in 0.05 M potassium phosphate buffer (1:4, w/v) containing 1.15% KCl (pH 7.6) using a Teflon glass homogenizer. The homogenate was centrifuged at 3200 g for 20 min at 4°C. The activity of t-GPx was determined as described by Rotruck et al.
28
using cumene hydroperoxide (cumene-OOH) as a substrate. Two hundred microliters of the liver supernatant was added to 200 μL of 0.4 M Tris-HCl buffer, pH 7, and 100 μL of 10 M sodium azide (to inhibit catalase activity). Then 200 μL of 2 mM GSH and 100 μL of 0.2 mM cumene-OOH were added, mixed well, and incubated at 37°C for 10 min along with the control tubes, which contained all the reagents but no enzyme. The reaction was arrested by the addition of 0.4 mL of 10% TCA, and 200 μL of the liver supernatant was added to the control tubes then centrifuged at 1780 g for 15 min. The supernatants were assayed for GSH using Ellman's reagent as mentioned before. The specific activities of t-GPx were calculated as μg/mL/min according to the following equations:
The specific activities of t-GPx in the samples were expressed in μg GSH consumed/min/mg protein.
Glutathione reductase (EC 1.8.1.7)
Liver tissues were homogenized in 0.05 M potassium phosphate buffer, pH 7.2 (1:10, w/v), and centrifuged at 3200 g for 20 min at 4°C. GSR activity was determined in the supernatant according to the method described by Goldberg et al. 29 The reaction mixture contained 1 mL of 0.05 M potassium phosphate buffer containing 15 mM EDTA, pH 7.2, 20 μL liver homogenate, and 10 μL of 65.3 mM GSSG. The mixture was mixed and incubated at 37°C for 5 min, and then 10 μL of 9.6 mM NADPH was added to start the reaction. The decrease in absorbance was read at 340 nm, and the specific activity was expressed as μmol of NADPH oxidized/mg protein/min.
Glutathione-S-transferase (EC 2.5.1.18)
GST was measured spectrophotometrically according to the method described by Habig et al. 30 with modification. Ten microliters of the substrate (1 mM P-nitrobenzyl chloride in 95% ethanol) were mixed well with 1.37 mL of 0.1 M K-phosphate buffer, pH 6.5, and 25 μL of the liver homogenate. The mixture was incubated at 37°C for 5 min, and then 100 μL of 5 mM GSH were added to start the reaction and incubated for 20 min at room temperature. The absorbance was read at 310 nm, and the specific activity was expressed as μmol/mg protein/min.
Superoxide dismutase (EC 1.15.1.1)
Cu–Zn–SOD activity was determined by the indirect method using pyrogallol, as described by Marklund and Marklund. 31 The assay mixture contained 20 μL of the liver homogenate, 1 mL of Tris-HCl buffer containing 1 mM diethylene triaminopenta acetic acid (DTPA), pH 8.2, and 10 μL of 20 mM pyrogallol. The change in the absorbance was read at 420 nm, and the specific activity was determined as IU/mg protein/min.
Liver function tests
Total proteins in serum and liver tissues were determined according to the method of Biuret. 32 Albumin was determined in serum by the method described by Doumas et al. 33 Total bilirubin concentration was determined according to the method of Jendrassik and Grof. 34 Liver AST (EC 2.6.1.1) and ALT (EC 2.6.1.2) activities were assayed according to the method of Reitman and Frankel. 35
Histopathologic study
The liver tissues were fixed, processed, and embedded in paraffin wax. Sections of 5 μm in thickness were cut and stained with hematoxylin and eosin.
Statistical analysis
The data were given as individual values and as means ±standard deviation (SD) for seven animals in each group. Comparisons between the means of various treatment groups were analyzed using a least significant difference (LSD) test. Differences were considered significant at P<.05. All statistical analyses were performed using the statistical software SPSS v11.5 (SPSS, Inc., Chicago, IL, USA).
Results
Total proteins in liver tissues
Values are expressed as mean±standard deviation (SD) for seven rats. Values with different letters are significantly different at P<.05.
C, control group; D–P, diethylnitrosamine–phenobarbital; PJ, pomegranate juice; ALT, alanine aminotransaminase; AST, aspartate aminotransferase.
DNAF in liver tissue
The agarose gel electrophoresis showed very low or undetectable DNA laddering (DNAF) in the hepatocytes of the control. The DNA intact band appears to be condensed near the application point with no DNA smearing suggesting no DNAF. DEN and PB administration resulted in a massive DNAF compared to the C group. Treatment with PJ pre, during, and post DEN and PB administration decreased DNAF compared to the D–P group. DNAF was increased after administration of PJ alone compared to the C group (Fig. 1).
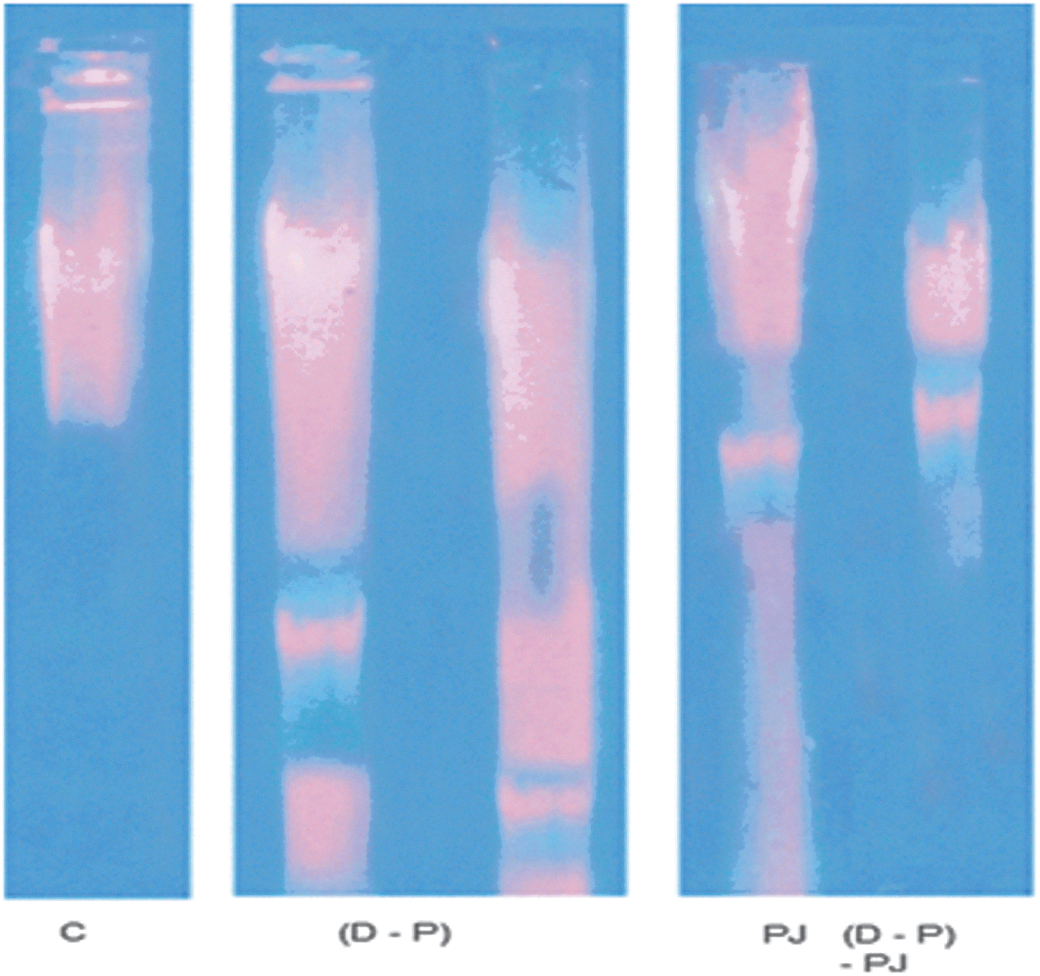
DNA fragmentation in all studied groups. C, control; D–P, diethylnitrosamine-phenobarbital; PJ, pomegranate juice. Color images available online at
Caspase-3 activity
The enzyme activity was increased significantly after DEN and PB administration by about 130% compared to the C group. Treatment with PJ pre, during, and post DEN and PB administration decreased caspase-3 activity significantly by about 65% compared to the D–P group. Administration of healthy rats with PJ alone increased caspase-3 activity significantly by about 91% compared to the C group (Fig. 2A).
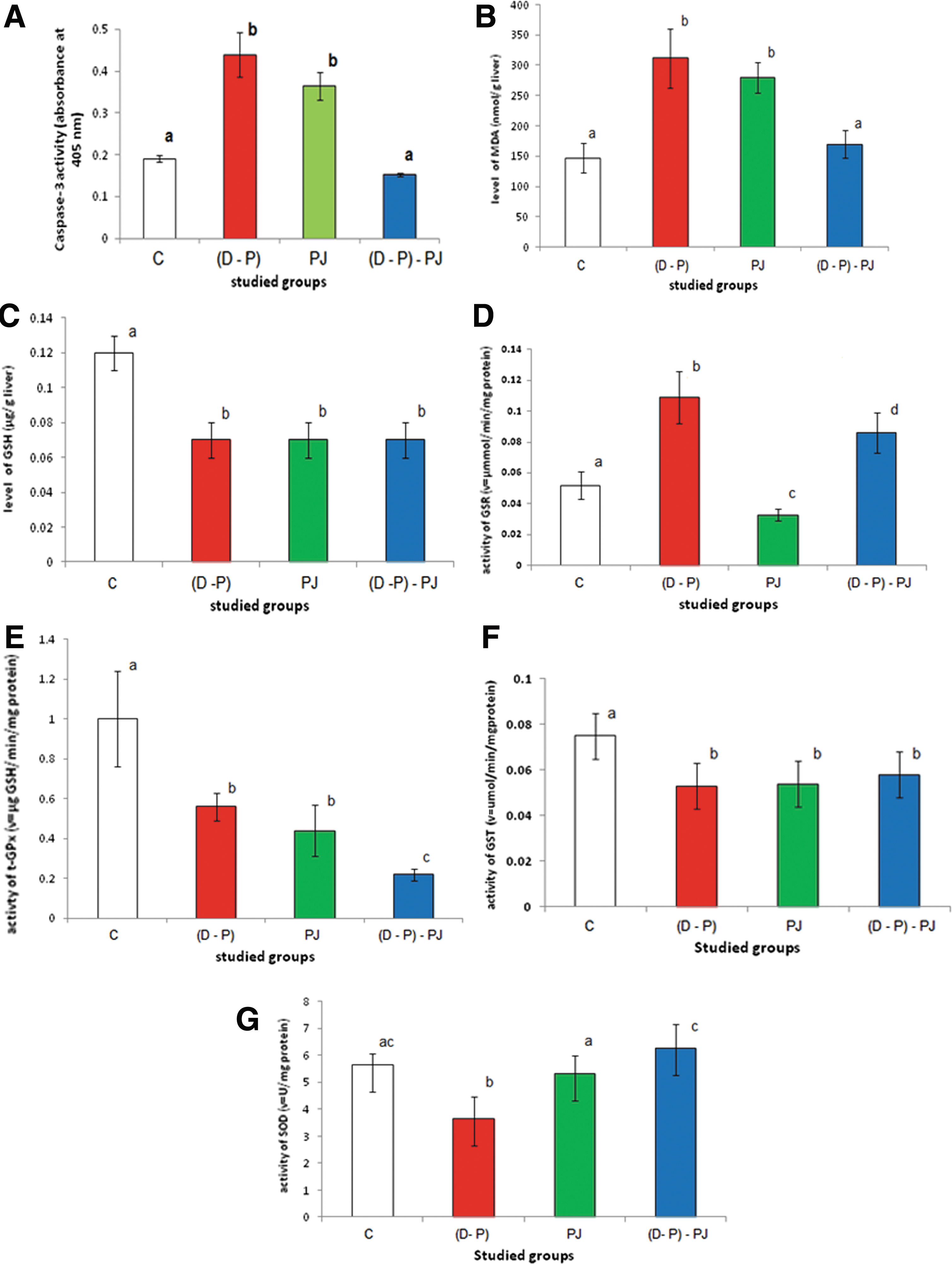
Effect of diethylnitrosamine (DEN) and phenobarbital (PB), PJ extract, and (D–P)–PJ on the activity of:
Malondialde-hyde levels
Administration of DEN and PB caused a significant elevation in MDA levels by about 112% compared to the C group (Fig. 2B). Treatment with PJ pre, during, and post DEN and PB administration caused a significant decline in MDA levels by about 46% compared to the D–P group. MDA levels were increased significantly by about 90% in rats administered with PJ alone compared to the C group.
Glutathione levels
DEN and PB administration decreased GSH levels significantly by about 42% compared to the C group (Fig. 2C). Treatment with PJ pre, during, and post DEN and PB administration showed no change in GSH levels compared to the D–P group. GSH levels were decreased significantly by about 41.67% after administration with PJ compared to the C group.
Glutathione reductase activity
GSR activities were elevated significantly in rats treated with D and P by about 110% compared to the C group (Fig. 2D). Treatment with PJ pre, during, and post DEN and PB administration caused a significant decline in GSR activity by about 21% compared to the D–P group. GSR activity decreased significantly by about 36.54% after administration with PJ compared to the C group.
Total glutathione peroxidase activity
T-GPx activities in rats injected with DEN and PB were decreased significantly by about 44% compared to the C group (Fig. 2E). Treatment with PJ pre, during, and post D–P administration decreased t-GPx activity significantly by 60.71% compared to the D–P group. The activity of t-GPx decreased significantly by about 56% after treatment with PJ compared to the C group.
Glutathione-S-transferase activity
Administration of DEN and PB decreased GST activity significantly by about 29.33% compared to the C group (Fig. 2F). Treatment with PJ pre, during, and post DEN and PB administration showed a nonsignificant elevation in GST activity by about 9.4% compared to the D–P group. GST activity decreased significantly by about 28% after treatment with PJ compared to the C group.
Superoxide dismutase activity
SOD activity decreased significantly (P<.001) after DEN and PB administration by about 35.7% compared to the C group (Fig. 2G). The enzyme activities were increased significantly in rats treated with PJ pre, during, and post DEN and PB administration by about 72% compared to the D–P group. SOD activity decreased nonsignificantly by about 6.2% after treatment with PJ compared to the C group.
Alanine aminotransaminase and aspartate aminotransferase
ALT and AST activities decreased significantly by about 50.4% and 13.7% respectively after DEN and PB administration compared to the C group (Table 1). Treatment with PJ pre, during, and post DEN and PB administration increased ALT activity significantly by about 60.5% with a nonsignificant decline in AST activity of about 3.3% compared to the D–P group. Administration of PJ increased ALT activity significantly by about 28.5% and decreased AST activity significantly by about 8.8% compared to the C group.
Total bilirubin, albumin, and total protein levels in serum
Administration of DEN and PB caused a significant elevation in bilirubin levels by about 1902%, with a significant decline in levels of albumin and total proteins by about 25% and 54% respectively compared to the C group (Table 1). Treatment with PJ pre, during, and post DEN and PB administration decreased bilirubin levels significantly by about 63%, with a significant elevation in the levels of albumin and total proteins by about 31.5% and 131% respectively compared to the D–P group. Administration of PJ decreased the levels of total proteins and bilirubin by about 15.5% and 15.5% respectively, while it increased albumin levels by about 26% compared with the C group.
Histopathologic results
Histopathologic examination of the C group showed normal rat liver with no remarkable pathological changes (Fig. 3A). Administration of DEN and PB showed liver cells with mild dysplastic nuclei, prominent nucleoli, and binucleated cells (Fig. 3B). However, treatment with PJ pre, during, and post D–P administration showed steatotic vacuoles within hepatocytes (Fig. 3C). It also showed ballooning of hepatocytes with no inflammatory or cirrhotic changes (Fig. 3D). Administration of PJ reveals hepatic tissue with slightly less architecture, fibrosis, and a mild collection of lymphocytic cells, congested vessels, and ballooning change with hepatocytes (Fig. 3E).
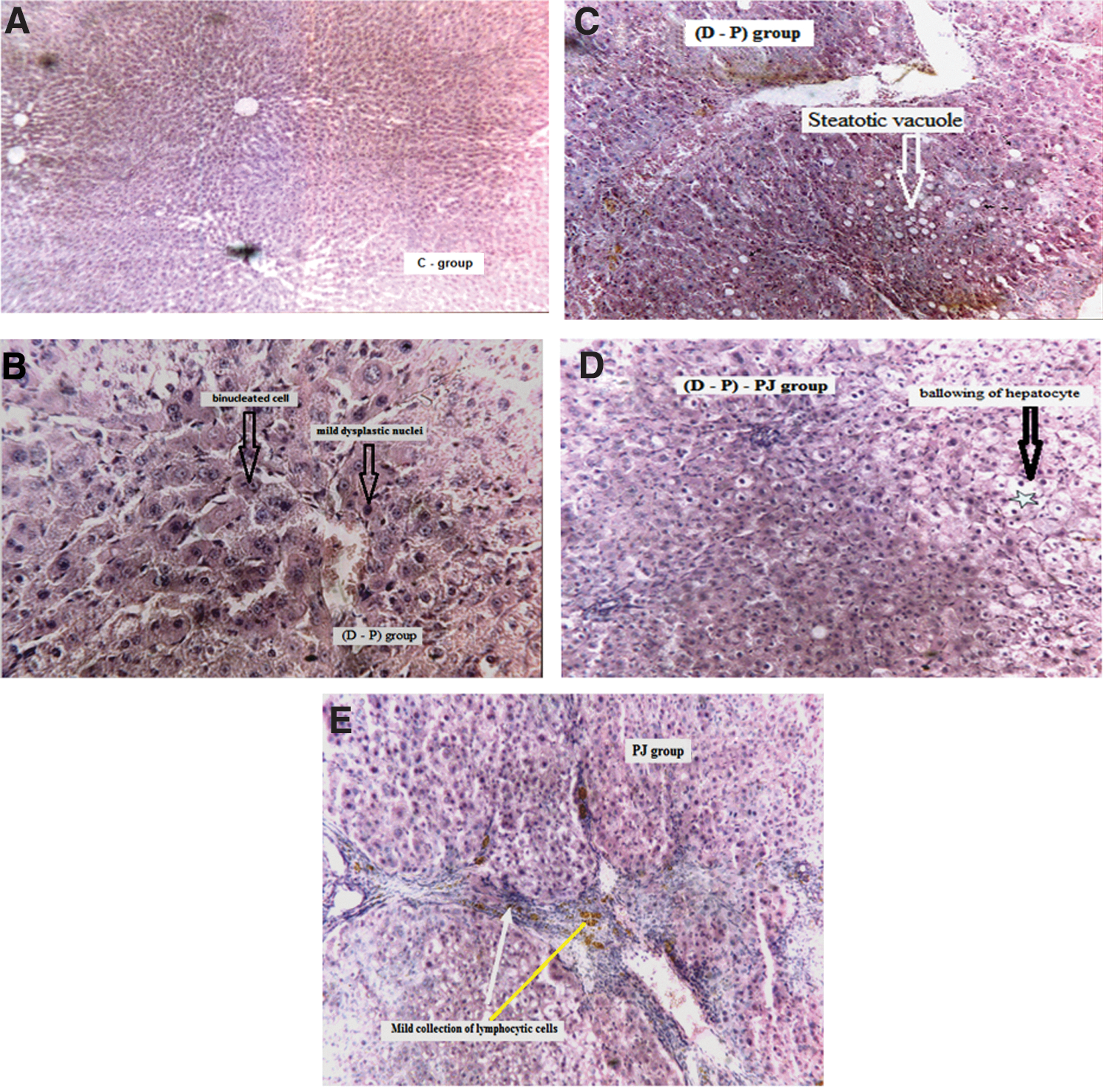
Microscopic examination of hepatic tissues of different studied groups.
Characterization of pomegranate juice
Total phenolic contents in pomegranate juice and its analysis
The results showed that PJ contains 0.43 mg/mL extract in terms of gallic acid equivalents. Figure 4 and Table 2 show high-performance liquid chromatography (HPLC) analysis of some of polyphenolic compounds in PJ and their concentrations.
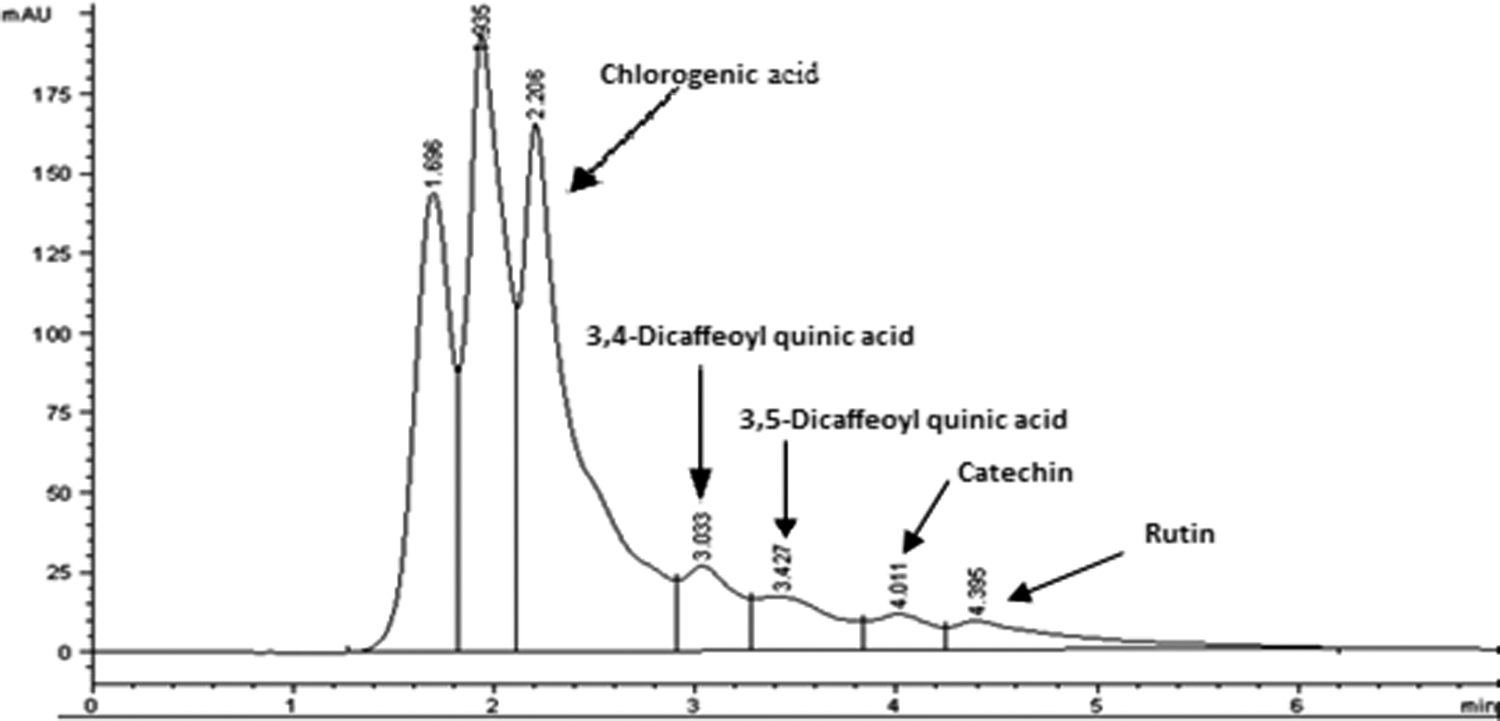
High-performance liquid chromatography analysis of polyphenolic compounds in PJ.
Discussion
Hepatocytes are very active in the metabolism of xenobiotics and drugs. ROS are generated during the detoxification of xenobiotics and drugs, and cause oxidative stress. Oxidative stress has been shown to be linked to liver toxicity and diseases. Hence, it is associated with damage to a wide range of macromolecular species, including lipids, proteins, and nucleic acids. 1 MDA is a major oxidation product of peroxidized polyunsaturated fatty acids, and increased MDA content is an important indicator of lipid peroxidation. 1 The present study has shown a significant elevation in MDA levels with a significant decrease in GSH levels in rat liver after DEN and PB administration. In addition, DEN and PB administration led to elevation in caspase-3 activity and DNAF levels compared to the control group. Formation of lipid peroxides in the crude homogenates resulted in response to the administration of DEN and PB. This may be due to an enhanced generation of O2 •− and H2O2 radicals that accelerated peroxidation of native membrane lipids. Peroxidation of the mitochondrial membrane led to a loss of cell integrity, increase in membrane permeability, and alteration of Ca2+ homeostasis that contribute to cell death due to alteration in the inner membrane potential. 1 In addition, some ROS interact with various tissue compounds leading to dysfunction and injury to the liver and other organs. 1 Elevation in caspase-3 activity and DNAF after DEN and PB administration indicates that DEN and PB caused apoptosis for liver tissues, and apoptosis is an adaptive process of combating excessive damage. Our results illustrate that the endonuclease-dependent fragmentation shows mixed smearing and laddering DNA fragments, indicating that exposure to DEN generates ROS, which trigger DNA damage causing cell death by necrosis and apoptosis. Otherwise, the histopathologic results show that DEN administration caused an elevation in the number of kuppfer cells, mild dysplastic nuclei, prominent nucleoli, and binucleated cells due to the release of mediators by the activated macrophage. DEN stimulates the metabolism in the liver microsomes causing the release of alkylating agents and the generation of superoxide radicals. The alkylating agents bind to both DNA-forming promutagenic adducts that may initiate liver carcinogenesis and RNA, leading to the inhibition of protein synthesis. 36,37 NO radicals, a product of DEN metabolism, are considered likely to be an important endogenous factor in hepatic injury, both by themselves and also in association with other agents. 13 Furthermore, a continuous administration of PB simultaneous with DEN increases the release of ROS. 22
Free radical scavengers such as GPx, GSR, GST, and GSH may protect biological systems from the deleterious effects of free radicals induced by xenobiotics and drugs. GSH acts as a cofactor to GPx 38 and as a nucleophilic scavenger of numerous compounds and their metabolites via enzymatic and chemical mechanisms. 1 The reduction in GSH levels after DEN and PB administration may be related to the reactions through oxidation and or conjugation, leading to the removal of peroxides and aldehydic products of lipid peroxidation. GSH depletion could also contribute to the activation of lipid peroxidation 39 —an observation that agreed with our results. In addition, the decrease in GSH levels after DEN and PB administration may possibly be due to increased demand of the tripeptide for lipid hydroperoxide metabolism by GPx and the interaction of GSH with DEN-derived free radicals in the same manner as reported by Barros et al. 40 about insecticide lindane.
Our results show that administration of DEN and PB caused a decline in SOD activity compared to the control group. SOD is the first line of defense in the body against superoxide radicals, and it is considered the most effective antioxidant. The inhibition of its activity may be related to the action of superoxide radicals, in a free state or after their transformation to H2O2, on oxidation of the cysteine in the enzyme. 1 Another explanation for the inhibition of SOD by DEN may be because of the direct interaction with the enzyme molecules and modification of the post-transcriptional or post-translational steps in the enzyme synthesis. 1 This result is similar to that of Hayakawa and Kuzuya 41 who reported that the inhibition of SOD activity in diabetic patients is due to the deleterious influences of oxidative stress.
GSR plays a key role in cellular defense against oxidative stress by preventing accumulation of GSSG and thus maintaining the redox state. The elevation in GSR activity after D and P administration compared to the control group probably reflects an adaptation to oxidative conditions. GSR was elevated in rat liver after administration with bacillus thuringiensis toxin. 1
GPx provides a second line of defense against hydroperoxides by catalyzing the reduction of H2O2 or organic hydroperoxides (ROOH) in the presence of GSH. 1 The decrease in GPx and GST activities after DEN and PB administration may be related to the ability of DEN to modify the post-transcriptional and post-translational steps in synthesis of these enzymes. This explanation is in agreement with the findings of Sahoo et al. 42 about insecticides.
Our results show that the administration of DEN and PB alone caused a decline in the levels of AST, ALT, total protein, and albumin, with an elevation in total bilirubin levels in serum compared to the control group. This indicates that liver cell death occurs by apoptosis as well as by necrosis. Hepatocytes dying by apoptosis presumably synthesize less AST and ALT as they wither away. Our results are similar to those of Pradeep et al. 13 who reported that DEN-induced hepatotoxicity was judged by elevated serum ALT, AST, and bilirubin levels and a simultaneous fall in their levels after 30 days. Previous studies have demonstrated that hepatocyte necrosis in acute hepatitis, toxic injury, or ischemic injury results in the leakage of enzymes into the blood circulation. 13 However, in chronic liver diseases such as hepatitis C and cirrhosis, serum ALT levels correlate only moderately well with liver inflammation. In hepatitis C, liver cell death occurs by apoptosis as well as by necrosis. This probably explains why at least one third of patients infected with hepatitis C virus have persistently normal serum ALT levels despite the presence of inflammation on liver biopsy. 43 Patients with cirrhosis often have normal or only slightly elevated serum AST and ALT levels. Thus, AST and ALT lack some sensitivity in detecting chronic liver injury.
Several studies have shown that fresh PJ is about 30% of the fruit weight. It contains 85% water, 10% total sugars, 1.5% pectin, ascorbic acid, amino acids, and organic acids. As shown from our results, PJ contains high amounts of total polyphenolic compounds. HPLC analysis showed that some of these polyphenolic compounds are chlorogenic acid, 3,4-dicaffeoyl quinic acid, 3,5-dicaffeoyl quinic acid, catechin, and rutin. Our results agree with the previous studies, which reported that PJ contains from 0.2% to 1% soluble polyphenols. These polyphenols include phenolic acids (gallic acid, ellagic acid, caffeic acid, chlorogenic acid, and ρ-coumaric), flavonoids (catechin, epicatechin, epigallocatechin 3-gallate, and quercetin), anthocyanins
Treatment with PJ pre, during, and post DEN and PB administration showed that there was no change in GSH levels compared with the D–P group, probably due to the oxidation of indoleamines, such as serotonin, tryptamine, and melatonin in PJ by peroxidase in the presence of H2O2 forming semiquinone radicals, which may form GSH adducts. 22,51 In addition, some polyphenols in PJ such as caffeic and chlorogenic acids can form GSH adducts during their metabolism rather than GSSG. 52 Therefore, GSH was consumed without a change in its level compared to the D–P group. These explanations are confirmed by our results about the administration of PJ alone as mentioned below.
Administration of PJ alone in the present study showed a reduction in the levels of total protein, albumin, and bilirubin in serum compared to the control group. This treatment raised ALT while it reduced AST. Furthermore, total protein levels in liver tissues were decreased. The result shows a significant elevation in levels of MDA, DNAF, and caspase-3 activity, with a significant decline in GSH levels after PJ administration compared to the control group. This indicates that the administration of PJ for a long period (23 weeks) induced lipid peroxidation and apoptosis. The prooxidant and apoptotic effects of PJ on the liver tissues resulted from its administration for a long period, which led to the accumulation of polyphenols such as anthocyanin and gallic acid. Previous studies have shown that anthocyanin, punicalagin, and caffeic acid have antiproliferative and proapoptotic effects. 44,53 Long et al. 54 reported that polyphenols such as gallic acid, epigallocatechin gallate, and quercetin each generated H2O2 in a time- and concentration-dependent manner when added to cell culture media. Otherwise, polyphenols can induce lipid peroxidation process and cell damage in the presence of some metal ions in PJ, especially Cu2+ and Fe3+ ions. 44,55 Hence, iron overload is associated with liver damage, characterized by massive iron deposition in hepatic parenchymal cells, leading to fibrosis and eventually to cirrhosis. However, polyphenols act as antioxidants in the absence of these metal ions. 56 The elevation in lipid peroxidation after PJ administration led to a decline in GSH levels compared to the control group, probably due to the formation of GSH adducts through semiquinone and some polyphenols. 51,52 The decrease in GSH levels led to a decrease in GST and t-GPx activities. The decline in the activities of GSR and GST may be due to their inhibition by certain polyphenols in PJ such as tannic acid, quercetin, catechin, and so on. Our results are in accordance with Faria et al. 57 who reported that administration of PJ for 4 weeks led to a decline in GSH, GST, and GSR levels. A nonsignificant decline in SOD activity in the present study compared to the control group may be related to the elevation in ROS levels, particularly superoxide radicals. The elevation in ROS after treatment with PJ caused fibrosis with a mild collection of lymphocytes and congested vessels, since ROS are implicated in the development and pathological progress of hepatic fibrosis by stimulating the collagen synthesis through activating hepatic stellate cell. 38 In conclusion, the present study showed that PJ extract exerts a significant protective effect against DEN and PB induced oxidative stress and apoptosis in hepatocytes by augmenting host antioxidant defense mechanisms. This extract is a promising agent for the prevention of chemical-induced toxicity through enhancing the antioxidative and drug metabolizing enzymes, as well as lowering the extent of lipid peroxidation. Administration of PJ over a long period in healthy rats causes oxidative stress and apoptosis.
Footnotes
Acknowledgment
The authors thank Ahmed Alaa Abdul-Aziz for revising this work.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.