
Abstract
Resveratrol is a polyphenolic compound in red wine that has antioxidant and cardioprotective effects in animal models. Listeria monocytogenes is a pathogen that mainly affects immunocompromised individuals and is initially detected at the cell surface or in phagosomes by toll-like receptor 2. Many antioxidants also exert anti-inflammatory activities; therefore, we evaluated the anti-inflammatory properties of resveratrol by studying the various inflammatory responses induced by heat-killed L. monocytogenes (HKLM). Resveratrol strongly blocked HKLM-induced NADPH oxidase-1 mRNA and reactive oxygen species production by macrophages. Resveratrol also suppressed monocyte chemotactic protein-1 expression, cyclooxygenase-2 expression, prostaglandin production, inducible nitric oxide (NO) synthase expression, and NO production induced by HKLM. We investigated the signaling pathway involved in the resveratrol effect. HKLM stimulated glycogen synthase kinase 3β (GSK3β) and extracellular signal-regulated kinase 1/2 (ERK1/2) phosphorylation. The involvement of GSK3β and ERK1/2 was tested using inhibitors. While the GSK3β inhibitor LiCl potentiated the effect of HKLM, the MEK inhibitor U0126 blocked these responses. Additionally, pretreatment with resveratrol blocked phosphorylation of both kinases induced by HKLM. These results suggest that HKLM is strong inducer of inflammatory mediators, and that the inhibitory effect of resveratrol may be mediated by the GSK3β and ERK1/2 pathways.
Introduction
R
Listeria monocytogenes is a pathogenic food-borne bacterium that causes diseases such as listeriosis. L. monocytogenes resides in the host depending on key virulence factors such as internalin or listeriolysin O. 6,7 However, animals and humans defend against L. monocytogenes through innate receptors. Toll-like receptor 2 (TLR2) is the most well-known innate receptor for defense against L. monocytogenes. 8 Recent reports have shown that TLR4 cooperates in the recognition and regulation of a Listeria infection. 9 TLR2-deficient mice are more susceptible to L. monocytogenes infection than that of wild-type mice; that is, bacterial loads are higher, and macrophage activation is reduced in TLR2 knockout mice. 10 Previous reports have suggested that participation of MyD88, an intracellular adaptor protein in TLR2 signaling, is essential for defense against L. monocytogene. 11 However, TLR2 signaling through MyD88 induces complicated signaling events, leading to increased expression of proinflammatory cytokines, chemokines, and other effector molecules. 12 Recent studies have reported that TLR is related various diseases. TLR2 has been associated with rheumatoid arthritis, type I diabetes, ischemia per reperfusion injury, and atherosclerosis. 13 –16 However, research on various inflammatory mediators induced by heat-killed L. monocytogenes (HKLM) is insufficient, and further study is necessary.
The glycogen synthase kinase 3β (GSK3β) and β-catenin pathways are crucial in the balance between pro- and anti-inflammatory cytokine production. This pathway plays an essential role in inflammation and immune cells. 17,18 In particular, many groups have shown that GSK3β, through TLR signaling, is necessary for inflammation. For example, GSK3β regulates TLR-mediated cytokine production, and inactivation of GSK3β by LPS has a negative effect on production of the proinflammatory cytokine interferon-β. 19
This study focused on the inhibitory mechanism of resveratrol inflammatory mediator production by the TLR2 agonist HKLM. Our results indicated that resveratrol in macrophages had a significant protective role in the HKLM-induced inflammatory response through multiple mechanisms, and that the extracellular signal-regulated kinase 1/2 (ERK1/2) and GSK3β pathways are important for the anti-inflammatory effect.
Materials and Methods
Reagents and cell culture
Cell culture reagents, including fetal bovine serum, were obtained from Life Technologies (Grand Island, NY, USA). Antibodies against phosphor-GSK3β, GSK3β, phosphor-ERK1/2, and ERK1/2 were obtained from Cell Signaling Technology (Beverly, MA, USA), and HKLM was obtained from InvivoGen (San Diego, CA, USA). The inducible nitric oxide synthase (iNOS) antibody was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA), and the cyclooxygenase-2 (COX-2) antibody was obtained from Cayman Chemical (Ann Arbor, MI, USA). An MCP-1 immunoassay kit was obtained from R&D Systems (Minneapolis, MN, USA). Resveratrol, NADPH, lucigenin, and LiCl were obtained from Sigma-Aldrich (St. Louis, MO, USA). Carboxy-H2-DCFDA was obtained from Life Technologies (Carlsbad, CA, USA), and various inhibitors were purchased from Merck KGaA (Darmstadt, Germany). The Raw264.7 macrophage cell line was obtained from the American Type Culture Collection (Manassas, VA, USA).
Measurement of reactive oxygen species generation
Production of reactive oxygen species (ROS), such as hydrogen peroxide, was determined by measuring changes in dichlorofluorescein diacetate (DCF-DA) fluorescence, an oxidation-sensitive fluorescence probe. Briefly, cells were seeded at 5×105 per well in six-well tissue culture plates. On the second day after seeding, cells were loaded with 1 mL fresh DMEM containing 50 μM DCF-DA and incubated at 37°C for 40 min. The change in fluorescence intensity was monitored by flow cytometry using a FACSCalibur apparatus (Becton-Dickinson, Franklin Lakes, NJ, USA)
Measurement of NO generation
Synthesis of NO was determined by assaying culture supernatants for nitrite, the stable reaction product of NO. Briefly, 100 μL of culture supernatant was reacted with 100 μL of Griess reagent (1% sulfanilamide, 0.1% naphthylethylenediamine dihydrochloride, and 2.5% phosphoric acid) at room temperature for 10 min. The optical density of the assay sample was measured spectrophotometrically at 570 nm. A fresh culture medium served as the blank in all experiments. The nitrite concentration was calculated from a standard curve derived from the reaction of NaNO2 under assay conditions.
Enzyme-linked immunosorbent assay
Supernatants from treated macrophages were collected and analyzed for PGE2 or PGI2 using sandwich enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems), according to the manufacturer's instructions.
Western blot analysis
Macrophages were cultured in six-well plates and treated with HKLM in the presence or absence of an inhibitor. Proteins were separated by 8% reducing sodium dodecyl sulfate–polyacrylamide gel electrophoresis and immunoblotted onto nitrocellulose membranes in 20% methanol, 25 mM Tris, and 192 mM glycine. Membranes were then blocked with 5% nonfat dry milk and incubated with the primary antibody for 18 h. The membranes were then washed, incubated for 90 min with the secondary antibody conjugated to horseradish peroxidase, rewashed, and finally developed using an enhanced chemiluminescence system.
Reverse transcription–polymerase chain reaction and real-time polymerase chain reaction
Total RNA was extracted from cells using a total RNA Miniprep kit (Axygen Biosciences, Union City, CA, USA). One microgram of total RNA was used as a template to make first-strand cDNA by oligo-dT priming using the Promega Reverse Transcriptase System (Promega, Madison, WI, USA). Polymerase chain reaction (PCR) was then performed using specific primers for target genes, as well as primers for β-actin as an internal standard. Real-time PCR was performed using a LightCycler 1.5 Thermal Cycler (Roche Diagnostics, Almere, The Netherlands) with SYBR-Green I as the florescent dye, according to the manufacturer's instructions. The cDNA obtained was then amplified using two synthetic primers specific for mouse Nox-1, MCP-1, and β-actin. The fluorescent synthetic gene-specific primer sets used for PCR were (1) Nox1 forward primer, 5′-AAGTGGCTGTACTGGTTGG-3′, and reverse primer, 5′-GTGAGGAAGAGTCGGTAGTT-3′, which amplified 238 bp of mouse Nox1 cDNA; (2) MCP-1 forward primer, 5′-AGAGAGCCAGACGGGAGGAA-3′, and reverse primer, 5′-GTCACACTGGTCACTCCTAC-3′, which amplified 520 bp of mouse MCP-1 cDNA; (3) β-actin forward primer, 5′-AGAGGGAAATCGTGCGTGAC-3′, and reverse primer, 5′-CAATAGTGATGACCTGGCCGT-3′, which amplified 137 bp of mouse β-actin cDNA.
Results
Resveratrol inhibits HKLM-induced Nox-1 expression and ROS generation
ROS are well-known inducers of inflammation, and Nox-1 is an ROS-producing enzyme in macrophages. 20 Stimulation of HKLM, a well-known TLR2 agonist, resulted in a rapid increase in Nox-1 mRNA expression in Raw264.7 cells (Fig. 1A). The effects of resveratrol on Nox-1 mRNA expression were confirmed using a real-time PCR analysis. Similar to Figure 1A, HKLM showed a time-dependent increase in Nox-1 mRNA expression, but pretreatment with resveratrol effectively inhibited the Nox-1 mRNA expression caused by HKLM (Fig. 1B). While HKLM induced ROS production in macrophages, pretreatment with resveratrol partially inhibited the effects of HKLM (Fig. 1C). These results suggest that resveratrol inhibits ROS production by blocking HKLM-mediated Nox-1 expression, but the partial effects suggest the possibility of other types of ROS production mechanisms by HKLM.

Resveratrol suppresses heat-killed Listeria monocytogenes (HKLM)-induced NADPH oxidase-1 (Nox-1) expression and reactive oxygen species generation.
Resveratrol suppresses HKLM-induced MCP-1 expression
MCP-1 is an essential chemokine for initiating inflammation. 21 The effects of HKLM on MCP-1 expression were verified by reverse transcription–PCR. Treatment with HKLM time-dependently elevated MCP-1 mRNA expression (Fig. 2A). Pretreatment with resveratrol decreased the HKLM effects almost down to basal levels (Fig. 2B). Additionally, pretreatment with N-acetyl cysteine (NAC), an ROS scavenger, also inhibited HKLM-induced MCP-1 expression (data not shown). Therefore, we suggest that the inhibition of MCP-1 by resveratrol results from HKLM-induced ROS inhibition.
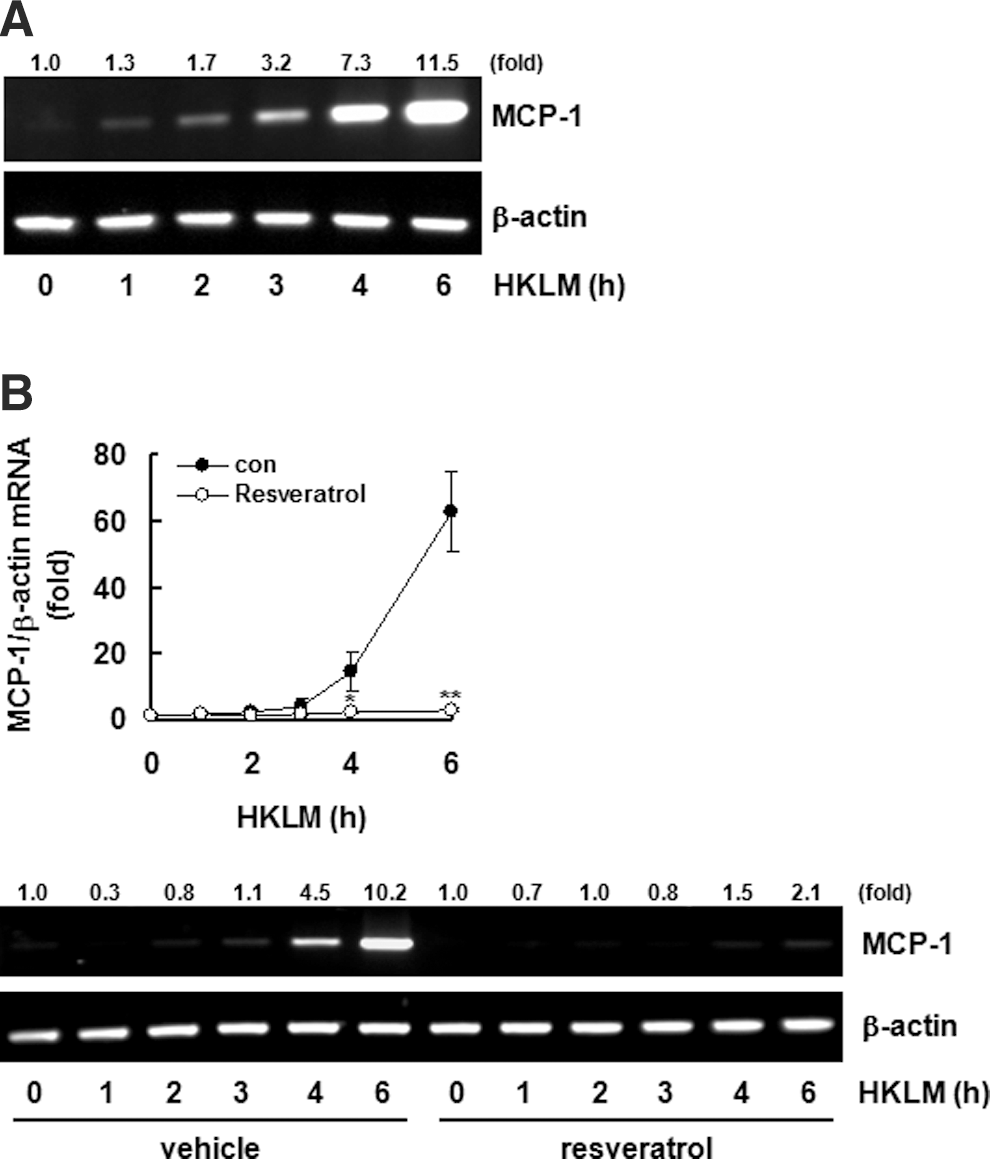
Resveratrol inhibits HKLM-induced monocyte chemotactic protein-1 (MCP-1) expression.
Resveratrol inhibits HKLM-induced COX-2 and iNOS expression and their metabolite production
COX-2 and its metabolites are well-known inflammatory mediators. COX-2 expression by macrophages in the presence of HKLM and the effects of resveratrol were analyzed by Western blot. HKLM treatment significantly induced COX-2 expression in macrophages, whereas pretreatment with resveratrol dose-dependently decreased COX-2 expression (Fig. 3A). Changes in PGE2 and PGI2 production among COX-2 metabolites also demonstrated that resveratrol dose-dependently inhibits HKLM-induced COX-2 metabolite (PGE2 and PGI2) production (Fig. 3B). iNOS expression and NO production, also important inflammatory mediators, were analyzed by the Western blot and Griess reagent analyses. HKLM induced iNOS expression and NO production by macrophages, but pretreatment with resveratrol dose-dependently inhibited HKLM-induced iNOS expression (Fig. 4A) and NO production (Fig. 4B).

Resveratrol suppress cyclooxygenase-2 (COX-2) expression induced by HKLM.
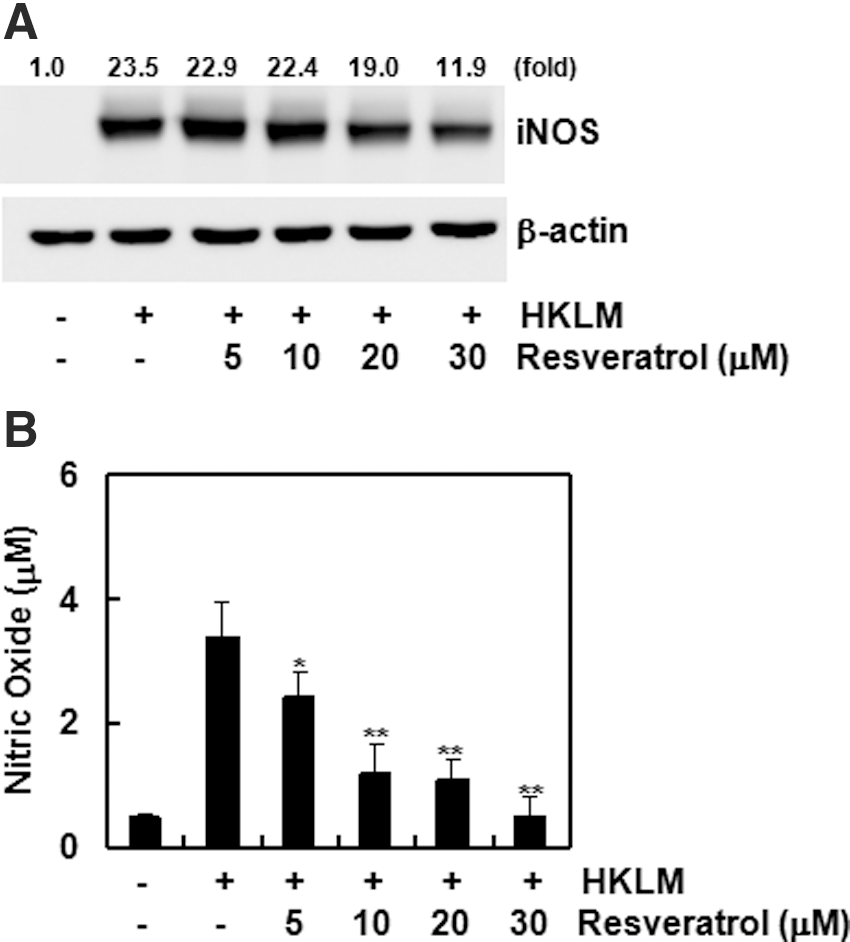
Resveratrol suppresses inducible nitric oxide synthase (iNOS) expression induced by HKLM.
Both ERK1/2 and GSK3β play a role in the resveratrol effect
Resveratrol inhibited production of various inflammatory mediators induced by HKLM. Studies of regulatory signaling proteins have shown a contribution by ERK1/2 and GSK3β as well. HKLM induced phosphorylation of ERK1/2 and GSK3β (Fig. 5A). However, pretreatment with resveratrol inhibited HKLM-induced ERK1/2 and GSK3β phosphorylation (Fig. 5B). Specific inhibitors were used to confirm the direct participation of ERK1/2 and GSK3β. Pretreatment with the ERK1/2 inhibitor U0126 inhibited HKLM-induced MCP-1, iNOS, and COX-2 protein expression (Fig. 5C). However, pretreatment with the GSK3β inhibitor LiCl increased HKLM-induced MCP-1, iNOS, and COX-2 expression (Fig. 5D). These results show that ERK1/2 and GSK3β are involved in HKLM-induced MCP-1, iNOS, and COX-2 expression.
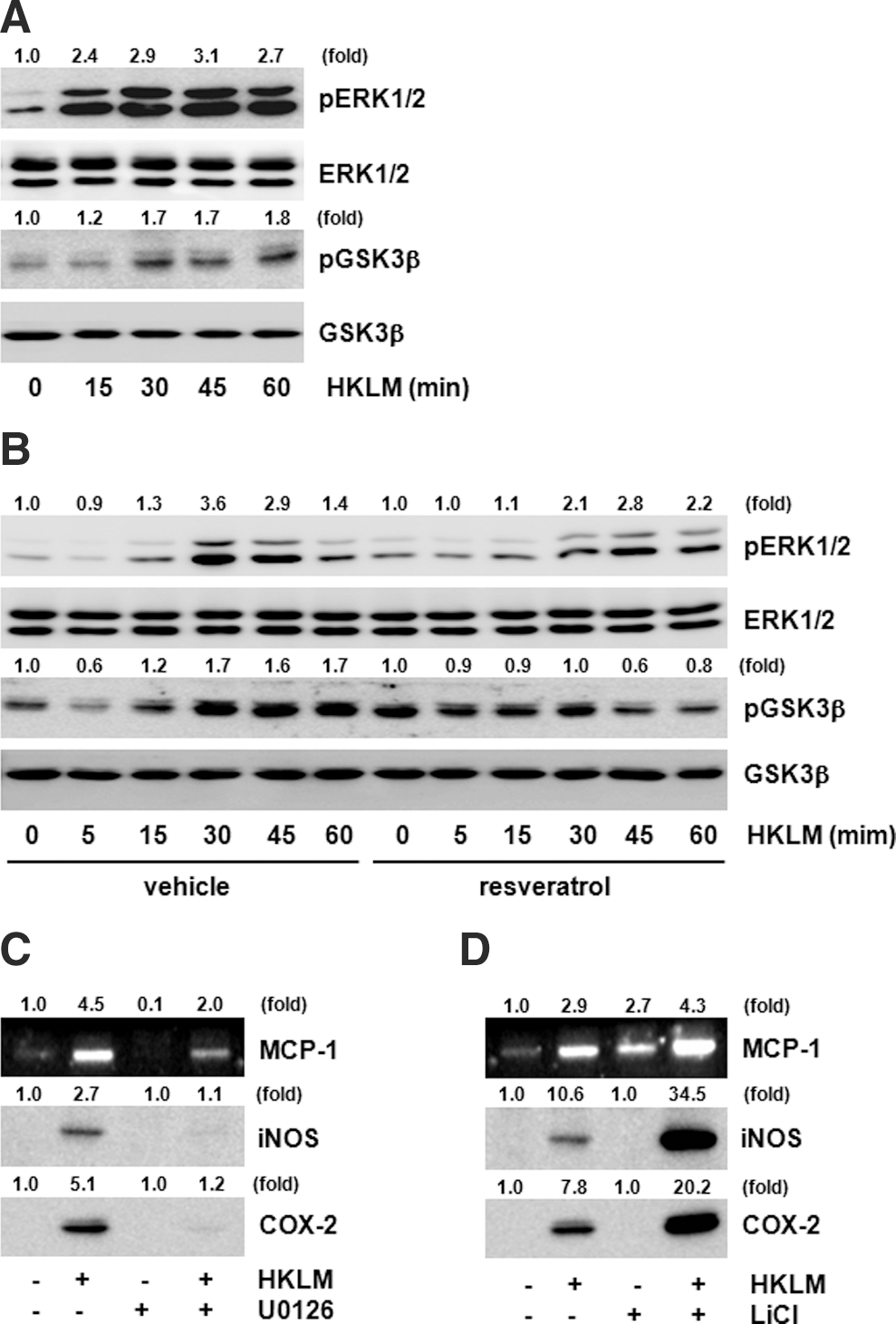
Extracellular signal-regulated kinase 1/2 (ERK1/2) and glycogen synthase kinase 3β (GSK3β) are targets for the effects of resveratrol.
Discussion
We investigated the effects of resveratrol on inflammatory mediator production induced by HKLM, a TLR2 agonist. Resveratrol showed anti-inflammatory effects by inhibiting HKLM-induced Nox-1 expression, ROS production, MCP-1 expression, COX-2 expression, metabolite (PGE2 and PGI2) production, and NO production after iNOS expression. These results suggest that resveratrol prevents the severe inflammatory response by an L. monocytogenes infection by inhibiting various types of mediators.
TLR contributes to infection and septicemia at the early stage by fusing with bacterial ligands. TLR produces various inflammatory mediators such as cytokines, chemokines, PGEs, and NO. 22 Among the many TLRs, TLR2 and TLR4 have been studied the most and are general TLRs that cause inflammation. There are many types of TLR2 ligands, including lipoproteins, and HKLM is known as a TLR2 ligand. 23
HKLM is Gram-positive coccobacillus heat-attenuated L. monocytogenes, which usually grows in clusters in a normal medium, and has a similar morphology to that of Streptococcus. While HKLM usually causes disease in livestock, it infects humans as well, inducing intestinal diseases. HKLM is known as a TLR2 ligand, but research on its inflammatory mechanism is unclear. Our results confirmed that HKLM has a similar mechanism as that of other TLR2 ligands. HKLM induced MCP-1 expression, iNOS expression, NO production, as well as COX-2 and PG expression, resulting in inflammation. Additionally, HKLM significantly induced Nox-1 gene expression, which produces ROS. The HKLM mechanism is very similar to that of Pam3CSK4, a synthetic TLR2 ligand (data not shown). These results might provide a clue as to how L. monocytogenes induces inflammation.
ROS production induced by stimulating TLR2 has been reported previously, 24 but sources ROS within cells are diverse, leading to uncertainty of the ROS source. Recent reports have shown that Mycobacterium stimulates TLR2 receptors, and that TLR2 interacts with Nox-2 to produce ROS. 25 However, our results showed that Nox-1 expression is strongly induced by HKLM, suggesting that Nox-1 plays an essential role in ROS production caused by TLR2. While resveratrol dramatically inhibited Nox-1 expression, ROS production only decreased partially, suggesting the presence of other ROS sources.
Intracellular ROS production induces production of other types of mediators, including cytokines and chemokines, including the chemokine MCP-1. In particular, MCP-1 induces excess activation of macrophages and monocyte migration during initiation, eventually elevating the inflammatory response. 26 –28 HKLM-induced MCP-1 production was an anticipated result, but complete inhibition by resveratrol was a very exciting result. We speculate that inhibition of MCP-1 by resveratrol was probably induced by blocking ROS production. Many reports have shown that MCP-1 production is mediated by ROS. 29 –31 The effect of resveratrol on MCP-1 production was stronger than the most widely used antioxidant NAC. These results show that resveratrol is more effective than NAC in terms of general anti-inflammatory effects.
Recent interest in phenolic compounds derived from grapes has shown that they have a valuable function related to human health, and resveratrol is placed at the center of phenolic compound components. Grape components have cardiac reinforcing, antioxidant, anticancer, antiaging, and antibacterial effects, 32 but the anti-inflammatory effect is the most important. The components of resveratrol have anti-inflammatory effects in rats, mice, and humans. 33 Bralley et al. reported that grape phenolic components inhibit inflammation, edema, and white blood infiltration in mice induced with 12-O-tetra decanoylphorbol 13-acetate. 34 Additionally, this component shows comparable effects to indomethacin, a widely used medication for degenerative joint diseases. 35 These observations demonstrate that resveratrol has clear anti-inflammatory effects. However, the anti-inflammatory mechanism of resveratrol is unknown, and hypotheses are insufficient. Therefore, we investigated whether the anti-inflammatory effects of resveratrol also act on the HKLM system.
The most well-known mechanisms in TLR2-induced reactions are pathways, including the transcription factors nuclear factor-κB and IRF-3 followed by IRAK and TRAF6, which are mediated by MyD88 and Mal. 36 However, recent studies suggest other delivery pathways than such factors. We hypothesized a function for GSK3β. In fact, HKLM increased GSK3β phosphorylation. GSK3β phosphorylation weakens protein activation, and LiCl, a GSK3β inhibitor, elevated HKLM-induced inflammatory gene expression. These results suggest that GSK3β acts as a negative regulator of the HKLM-mediated inflammatory response. GSK3β is inactivated by various pathways.
The Akt pathway is considered important among various pathways. Akt is a typical protein that directly phosphorylates and inactivates GSK3β. 18 We have previously investigated the importance of Akt as an inflammation mediator. Although the importance of Akt was studied for the TLR4-induced inflammatory response, the Akt pathway inhibitor LY294002 also inhibited HKLM-induced inflammation (data not shown). We demonstrated that inactivating GSK3β elevated inflammation mediator production. Resveratrol restored the activation of GSK3β weakened by HKLM. We believe that this result shows that resveratrol regulates inflammatory mediator production by inhibiting the Akt-GSK3β pathway through HKLM. ERK1/2 is also a protein that weakens GSK3β activation. The ERK1/2 inhibitor U0126 inhibited three types of inflammatory gene expression by HKLM. These results suggest the further possibility that ERK1/2 is the main pathway regulated by HKLM. However, further research is necessary.
Our results suggest that HKLM is a strong inducer of inflammatory mediators, and that the inhibitory effect of resveratrol may be mediated by the GSK3β and ERK1/2 pathways.
Footnotes
Acknowledgment
This study was supported by a Korea Science and Engineering Foundation (KOSEF) grant funded by the Korea government (MEST) (2011-0013206 and 2011-0006181).
Author Disclosure Statement
No competing financial interests exist.