
Abstract
We investigated the hepatoprotective effects of the extract of dandelion leaves (EDL) on a murine model of methionine- and choline-deficient (MCD) diet-induced nonalcoholic steatohepatitis (NASH). C57BL/6 mice were fed for 4 weeks with one of the following diets: control diet (Cont), MCD diet (MCD), MCD diet supplemented with EDL at 200 mg/kg body weight·daily (MCD+D200), and MCD diet supplemented with EDL at 500 mg/kg body weight·daily (MCD+D500). Hepatic function was assessed by evaluating the following parameters: liver histology; plasma levels of alanine aminotransferase (ALT), triglyceride (TG), malondialdehyde (MDA), and reduced glutathione (GSH); expression levels of TNF-α and IL-6; and levels of caspase-3 and pJNK/JNK protein. Histopathological evaluations revealed that addition of EDL to the MCD diet dampens the severity of the clinical signs of NASH. Moreover, EDL led to a significant decrease in the serum levels of ALT, hepatic TG, and MDA, and in the expression levels of TNF-α, and IL-6; on the contrary, the levels of reduced GSH increased. At the post-transcriptional level, EDL significantly decreased the activation of procaspase-3 to active caspase-3, and the phosphorylation of JNK. These results suggest that the beneficial effects of EDL on NASH are mainly due to its antioxidant and anti-inflammatory activities.
Introduction
T
The common dandelion (Taraxacum officinale G. H. Weber ex Wiggers, Asteraceae) is widely used as an herbal folk medicine for treating various disorders such as liver disease, gallbladder disorders, digestive complaints, and arthritic and rheumatic diseases. 6 –8 The dandelion is known for its broad spectrum of biological activities, including anti-inflammatory, antioxidative, anticarcinogenic, and antihyperglycemic activities. 9 Recently, dandelion extracts have been shown to effectively reduce oxidative stress and fibrosis in CCL4-induced acute liver damage. 10,11 and to protect against alcohol-induced liver injuries. 12 Interestingly, extracts of dandelion leaves (EDLs) have stronger hydrogen peroxide-scavenging activity than do the root extracts, because of their high polyphenol contents. 9 On the other hand, EDLs inhibit lipopolysaccharide-induced oxidative stress and nitric oxide production via NF-κB modulation in RAW 264.7 cells, 13 and significantly protect against CCL4-induced liver injury in mice, indicating the presence of luteolin (including its glycosidic form) and polyphenol in the leaf extracts. 14 Moreover, the mechanism by which EDL exerts its hepatoprotective effects has not yet been fully uncovered. In this study, we evaluated the efficacy of EDL in preventing steatohepatitis in mice, and investigated the underlying mechanism.
Materials and Methods
Preparation of EDL
Dried, powdered dandelion leaves were obtained from Yanggu Mindlre (
HPLC analysis for luteolin
Luteolin was analyzed by HPLC (Jasco, Tokyo, Japan), equipped with a Model PU980 pump and a model UV975 detector. A YMC ODS-AM (250 mm×4.6 mm inner diameter; particle size, 5 μm) reversed-phase column (Kyoto, Japan). The mobile phase consisted of solvent A (0.1% acetic acid in water) and solvent B (0.1% acetic acid in acetonitrile). The gradient elution was as follows: 0 min, A:B (88:12); 18 min, A:B (78:22); 28 min, A:B (72:28); 35 min, A:B (62:38); 48 min, A:B (52:48); 54 min A:B (32:68); 58 min, A:B (0:100); 60 min, A:B (0:100); and 62 min, A:B (88:12). The column was equilibrated for 15 min before each analysis. Sample and standard solutions of 20 μL each were injected into the HPLC. The flow rate was 1.0 mL/min; temperature, 35°C; and wavelength, 285 nm (Fig. 1).
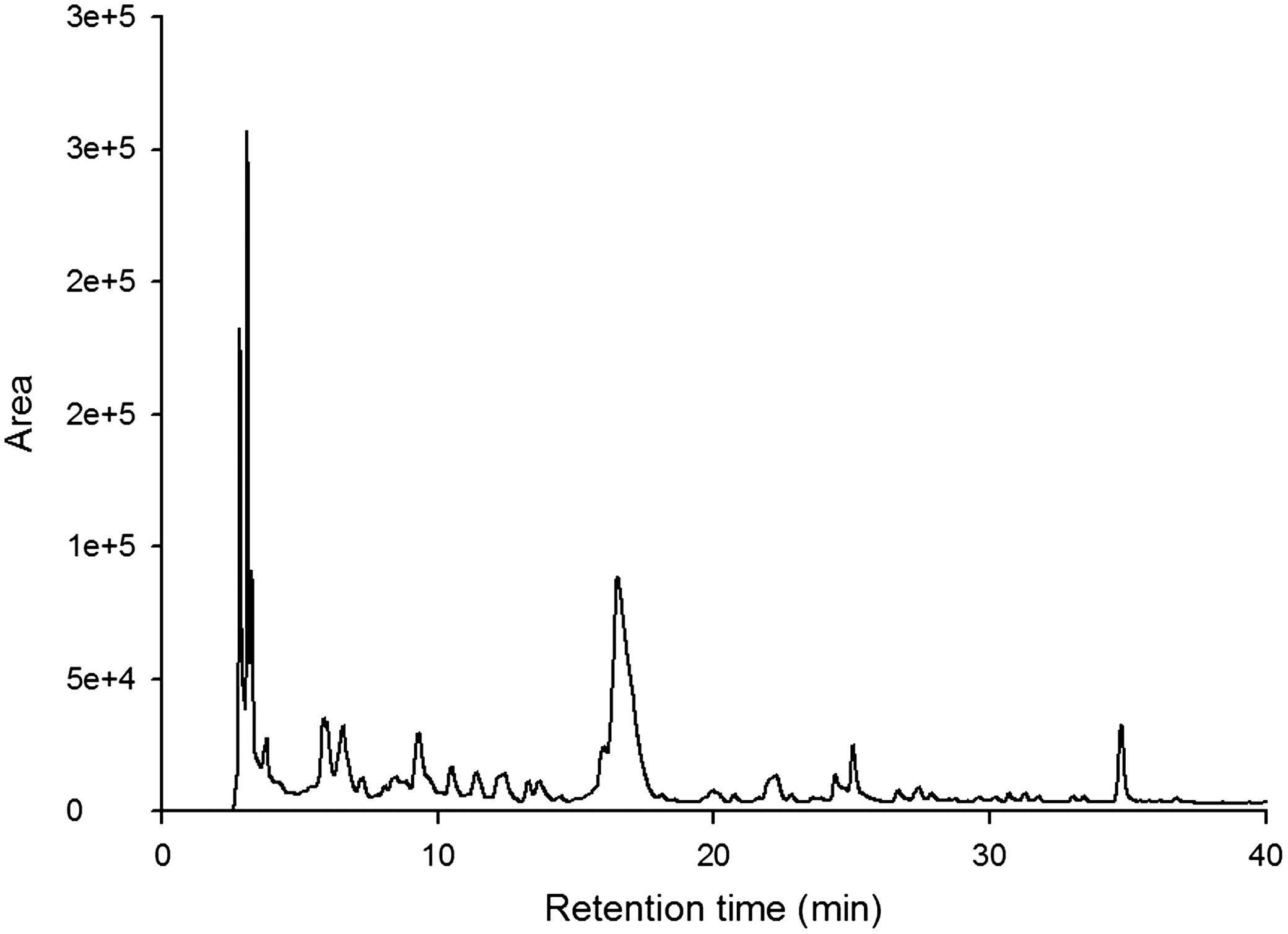
HPLC chromatogram of luteolin in extracts of dandelion leaves (EDLs). The arrowhead indicates luteolin.
Total phenol content
Folin–Ciocalteu reagent was used to determine total phenol content. Sample aliquots (1 mL) were mixed with 2% (w/w) Na2CO3. After 3 min, 0.1 mL Folin–Ciocalteu reagent was added to each reaction mixture. Reactions were incubated in the dark for 60 min; absorbance at 750 nm was recorded by UV spectrophotometry (Jasco) and then converted to total phenol content. The quantification was carried out using a calibration curve of chlorogenic acid.
Animals and diets
Forty 7-week-old male C57BL/6 mice were purchased from Charles River, Korea (Seoul, Korea), and housed in the Korea Food Research Institute (KFRI) at a constant temperature (22°C–26°C), under light/dark cycles of 12 h per day, with free access to water. They were allowed to adapt to their food and environment for 1 week before starting the experiment. Animals were body-weight matched and randomly divided into four groups of eight mice each: control diet (Cont; Dyets, Inc., Bethlehem, PA, USA), methionine- and choline-deficient diet (MCD; Dyets), MCD diet supplemented with EDL at 200 mg/kg body weight·daily (MCD+D200), and MCD diet supplemented with EDL at 500 mg/kg body weight·daily (MCD+D500). All four diet formulations were administered orally for 4 weeks. Body weight was measured weekly. At the day of sacrifice, a laparotomy was performed under ketamine and xylazine anesthesia (intramuscular injection of 100 mg/kg body weight and 5 mg/kg body weight, respectively), and blood samples were collected via abdominal heart puncture for biochemical determinations. Livers were weighed and fixed in 10% formalin for histological analysis or were snap-frozen in liquid nitrogen. Serum and liver samples were then stored at −70°C until required. All animal studies were performed in accordance with the guidelines approved by the Animal Experimentation Ethics Committee of the KFRI.
Histological examinations
Hematoxylin/eosin-stained paraffin-embedded 4-μm-thick liver tissue sections were graded for hepatic steatosis and necroinflammation, as described previously. 15 Hepatic steatosis (percentage of liver cells containing fat) was graded as 0 (0%), 1 (1–25%), 2 (26–50%), 3 (51–75%), or 4 (>75%). Necroinflammation was graded as 0 (no inflammatory foci), 1 (mild), 2 (moderate), and 3 (severe). Two investigators independently evaluated the slides.
Biochemical analyses
Serum alanine aminotransferase (ALT) levels were determined using spectrophotometric assay kits (Sigma, St. Louis, MO, USA). Total hepatic triglycerides (TG) were estimated using the Asan Triglyceride Kit (Asan Pharm, Seoul, Korea) according to the manufacturer's instructions.
Lipid peroxide assay
Liver malondialdehyde (MDA) levels, as an index of lipid peroxidation, were determined by the double-heating method. 16 This method is based on the spectrophotometric measurement of the purple color generated by the reaction of thiobarbituric acid (TBA) with MDA. Each liver tissue was homogenized (10% w/w) in 25 mM Tris–HCl, pH 7.4, and centrifuged at 8,000 g for 15 min. For the MDA assay, 2.5 mL of trichloroacetic acid solution (10%) was added to 0.5 mL of the supernatant, followed by heating in a boiling water bath for 15 min. Two milliliters of the reaction mixture was transferred to a test tube containing 1 mL of TBA solution (0.67%). Each tube was then placed in a boiling water bath for 15 min. After cooling to room temperature, the absorbance was measured at 532 nm.
Glutathione content
Glutathione (GSH) levels in liver tissue specimens were measured using the Glutathione Assay Kit (Cayman, Ann Arbor, MI, USA) according to the manufacturer's instructions.
Real-time PCR analysis
Total RNA was isolated from frozen liver tissues using TRIzol® reagent (Gibco, Grand Island, NY, USA). Total RNA (1 μg) was then reverse-transcribed into cDNA using AccuPower® PCR PreMix (Bioneer, Daejeon, Korea). The amplification reactions were performed on a Roche LightCycler® 480 system (Roche Diagnostics GmbH, Mannheim, Germany) using the following thermal cycling conditions: an initial activation step at 95°C for 3 min, followed by 45 cycles of denaturation for 10 sec at 95°C, annealing for 15 sec at 55°C, and extension for 20 sec at 72°C. β-Actin was used as a housekeeping gene, and the fold change in expression of each target mRNA relative to β-actin was calculated based on the comparative 2−ΔΔct expression method. Primers were as follows: TNF-α: forward, 5′-AGC CCC CAG TCT GTA TCC TT-3′; reverse, 5′-CTC CCT TTG CAG AAC TCA GG-3′; IL-6: forward, 5′-TAG TCC TTC CTA CCC CAA TTT CC-3′; reverse, 5′-TTG GTC CTT AGC CAC TCC TTC-3′; and β-actin: forward, 5′-CGT GCG TGA CAT CAA AGA GAA-3′; reverse 5′-TGG ATG CCA CAG GAT TCC AT-3′.
Western blot analysis
Immunoblotting analysis was performed as described previously. 17 Each frozen liver tissue specimen (10–20 mg) was homogenized in phosphate-buffered saline containing a protease inhibitor cocktail (Calbiochem, San Diego, CA, USA), and the total protein concentration was quantitated. The samples (50 μg of protein per lane) were mixed with a sample buffer, boiled for 10 min, separated by sodium dodecyl sulfate–polyacrylamide (10%) gel electrophoresis under denaturing conditions, and electroblotted onto nitrocellulose membranes.
The membranes were incubated overnight at 4°C with anticaspase-3 or antiphospho-JNK (dilution, 1:1000; Cell Signaling Technology, Beverly, MA, USA). The membranes were then re-blotted with antiactin or anti-JNK (dilution, 1:2000; Sigma, St. Louis, MO, USA) to verify the equal loading of protein in each lane. Each experiment was performed in triplicate.
Statistical analysis
All data are expressed as mean±standard deviation (SD). Analysis of variance was used for comparison between groups. A value of P<.05 was considered statistically significant.
Results
Luteolin and total phenol contents
The luteolin concentration of the EDL water was 4.8±0.01 μg/g, whereas the total phenol content was 8.017±0.13 mg chlorogenic acid equivalents per gram of EDL.
EDL has no effect on MCD-induced changes in body and liver weights
MCD diet is associated with the loss of body weight. 18 Thus, mice fed the MCD diet lost significant body weight compared with mice fed the control diet. However, addition of EDL to the MCD diet prevented further weight loss. The same observations were made with regard to liver weight (Table 1).
Data are mean±SD (n=6 per group).
P<.05, MCD diet vs. control diet.
EDL, extract of dandelion leaves; MCD, methionine- and choline-deficient; MCD+D200, MCD supplemented with 200 mg EDL/kg body weight daily; MCD+D500, MCD supplemented with 500 mg EDL/kg body weight daily; SD, standard deviation.
EDL reduces MCD-induced steatohepatitis in mice
We evaluated the effects of EDL on MCD diet-induced steatohepatitis in vivo. Mice fed the control diet had normal liver histology. On the other hand, mice fed the MCD diet developed steatohepatitis with hepatocyte-ballooning changes, lobular inflammatory cell infiltration, and inflammatory foci. Remarkably, mice fed the MCD diet supplemented with EDL exhibited a dose-dependent decrease in hepatic steatosis and inflammatory infiltration (Fig. 2A). Histological grading of liver sections confirmed that EDL significantly ameliorated hepatic steatosis and necroinflammation (Fig. 2B).

Effect of EDL on methionine- and choline-deficient (MCD) diet-induced steatohepatitis in mice.
EDL reduces MCD-induced high serum ALT levels and hepatic steatosis in mice
To evaluate the effect of EDL on MCD-induced liver injury, we measured serum ALT and liver TG levels in mice. As shown in Figure 3, the MCD diet resulted in a prominent increase in ALT serum levels and TG contents as compared with the control. In contrast, the MCD diet supplemented with EDL yielded dose-dependent inhibition of serum ALT levels and TG accumulation in comparison to the MCD diet alone. The MCD+D500 group showed significant inhibition of serum ALT levels and TG accumulation.
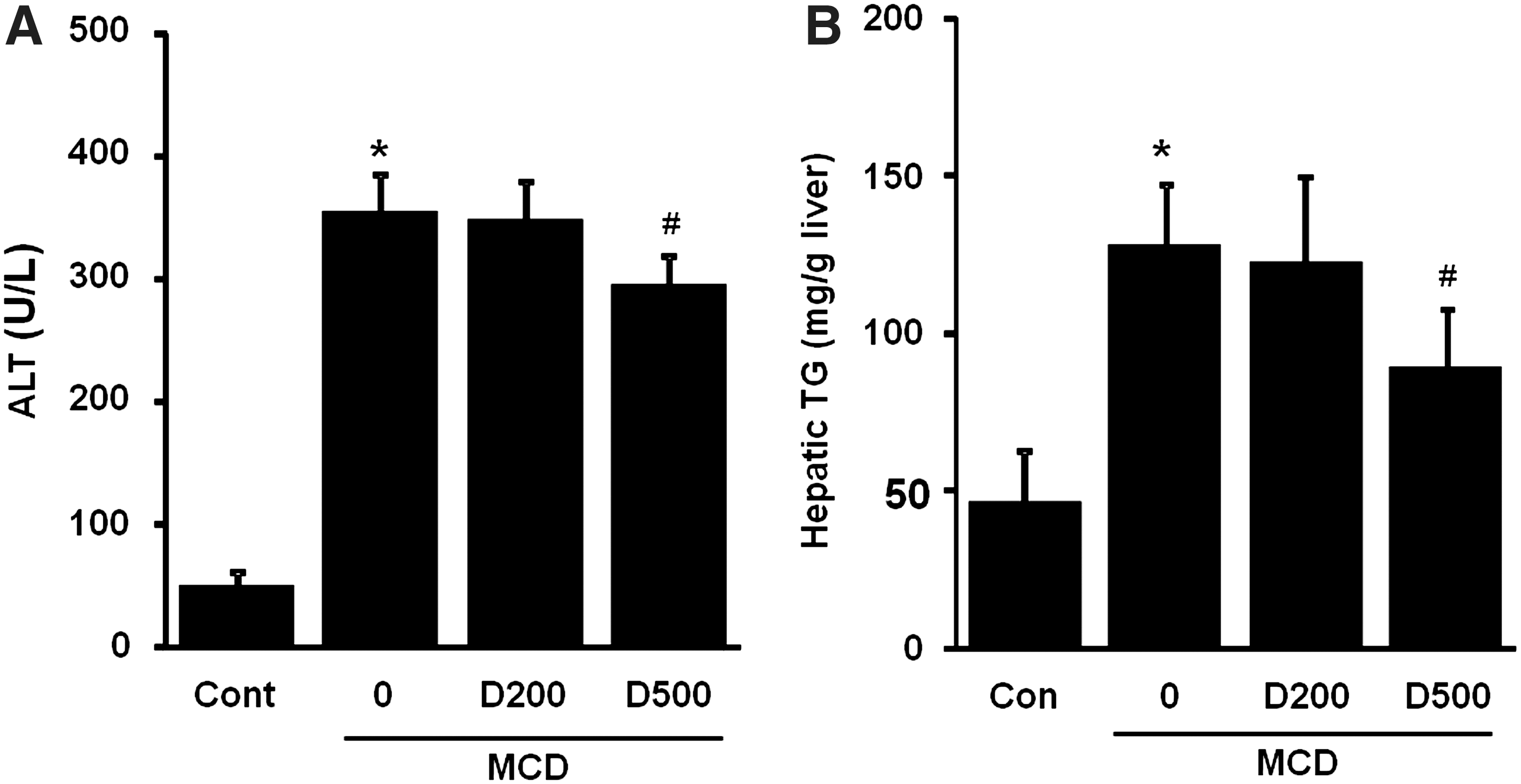
Effect of EDL on MCD-induced serum alanine aminotransferase (ALT) and hepatic steatosis in mice.
EDL reduces MCD-induced hepatic oxidative stress in mice
To explore the effect of EDL on the hepatic oxidative stress status, liver MDA and GSH contents were assessed. Liver MDA level is considered a marker of oxidative stress-induced lipid peroxidation. 19 After 4 weeks of feeding the MCD diet, MDA levels were significantly increased compared to the control diet group. This increase was accompanied by a decreased GSH content. Supplementation with D500, however, significantly lowered MDA levels and increased GSH content in comparison to the MCD diet alone (Fig. 4).
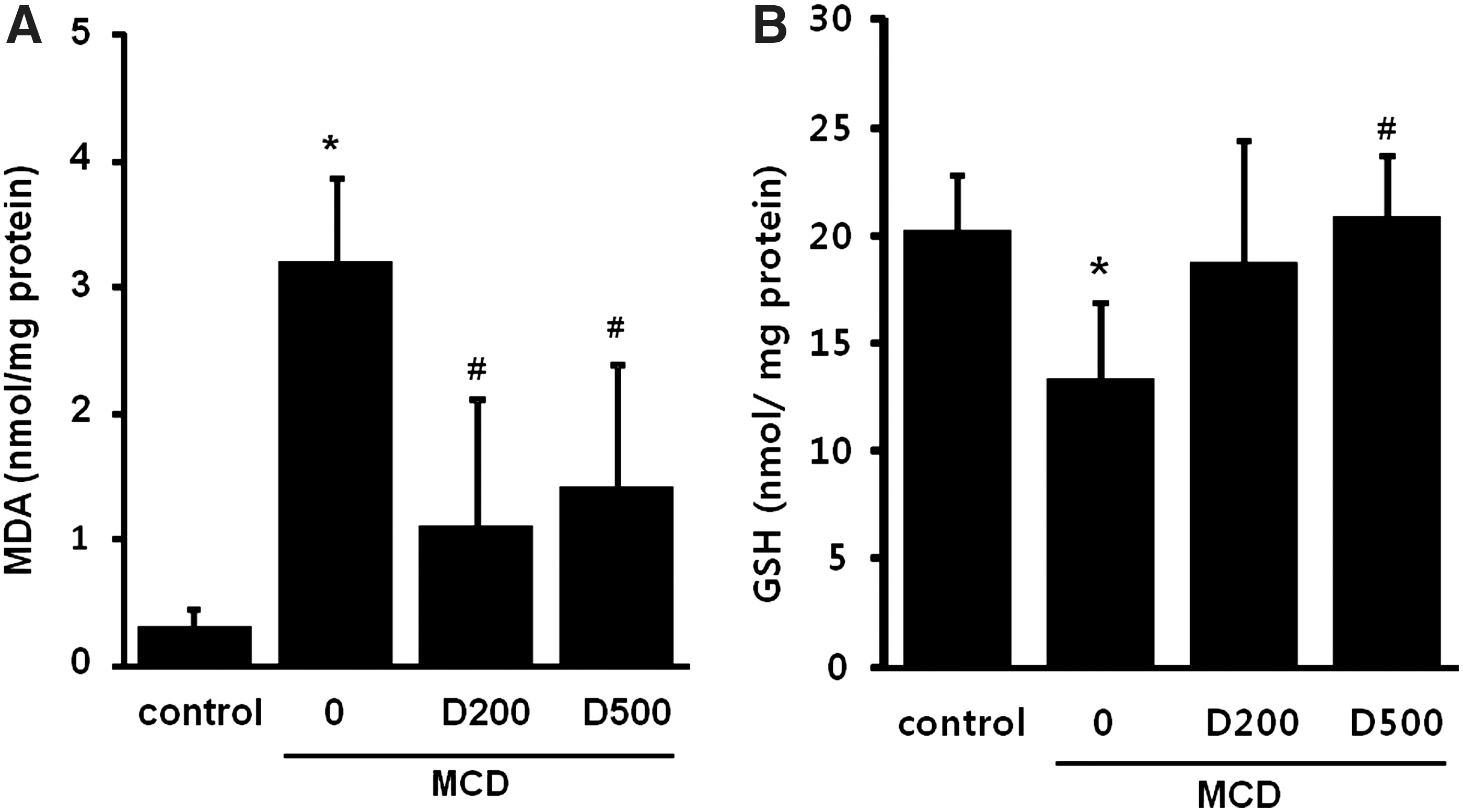
Effect of EDL on MCD-induced hepatic oxidative stress in mice.
EDL reduces MCD-induced TNF-α and IL-6 gene transcription in mice
To investigate the mechanisms underlying the effects of EDL on steatohepatitis, we determined the expression of the proinflammatory factors, TNF-α and IL-6. Mice fed the MCD diet showed a marked elevation in hepatic mRNA expression of TNF-α and IL-6 as compared with mice fed the control diet. Conversely, D500 significantly reduced mRNA expression of TNF-α in mice fed the MCD diet. Mice fed the MCD diet supplemented with EDL showed slightly lower IL-6 mRNA expression than mice fed the MCD diet alone (Fig. 5).

Effect of EDL on MCD-induced TNF-α and IL-6 gene transcription in mice. Expression of the hepatic inflammatory mediators
EDL reduces MCD-induced caspase-3 activation in mice
Procaspase-3 is commonly cleaved to active caspase-3 during caspase-dependent apoptosis. To evaluate the induction of caspase-dependent apoptosis by the MCD diet, we detected caspase-3 activation by immunoblotting. As shown in Figure 6, MCD diet resulted in a significant increase in the levels of active caspase-3 relative to the control diet. In contrast, EDL markedly attenuated MCD diet-induced activation of caspase-3 in a dose-dependent manner.
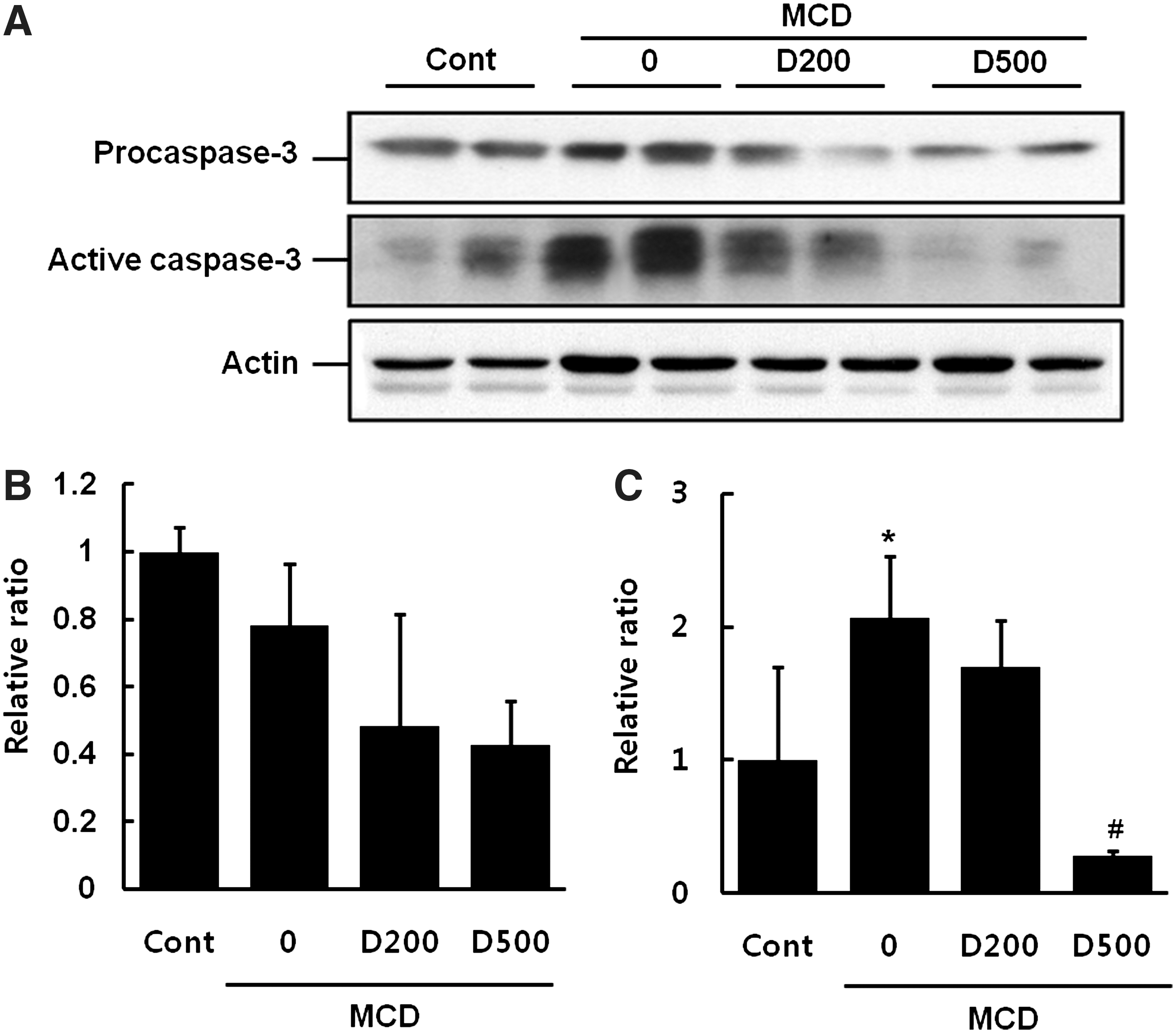
Effect of EDL on MCD-induced caspase-3 levels in mice.
EDL reduces MCD-induced JNK phosphorylation in mice
Many investigators have reported that oxidative stress activates JNK. 5,20 The MCD diet led to significant phosphorylation of JNK compared to the control diet. However, D500 significantly decreased the extent of JNK phosphorylation (Fig. 7).
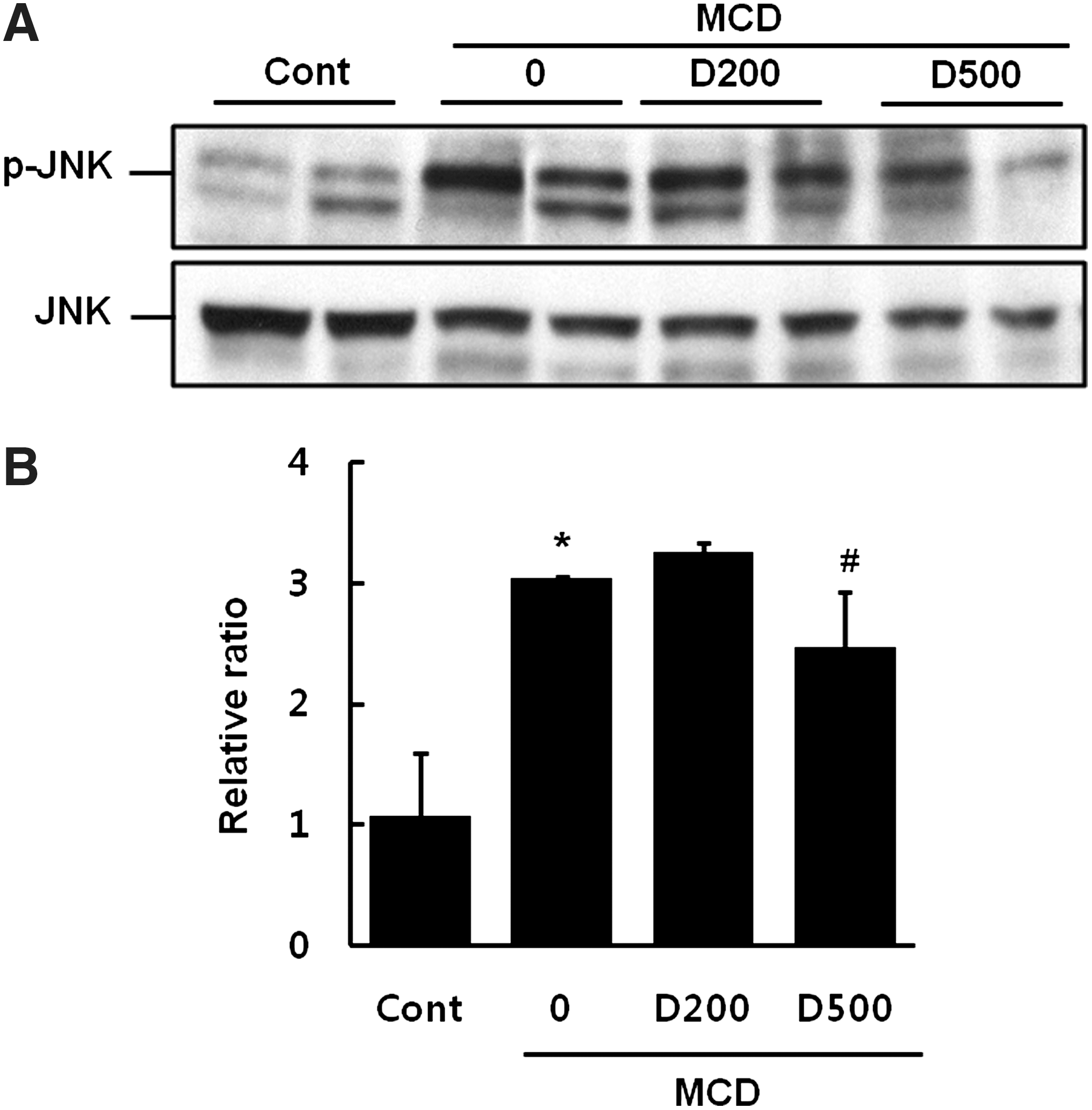
Effect of EDL on MCD-induced JNK phosphorylation in mice.
Discussion
A high-sucrose diet lacking methionine and choline causes hepatic steatosis, hepatocyte injury, inflammation, and ultimately, fibrosis, with pathological similarities to human NASH. 21 Hepatic steatosis is thought to be caused by the suppression of hepatic stearoyl-coenzyme A desaturase-1 mRNA expression, enhanced fatty acid oxidation, and increased energy expenditure, which induces lipid metabolism and loss of body weight, yet not affecting food intake. 18 Our study confirmed that the MCD diet leads to loss of body weight (Table 1) and to histological changes such as hepatic steatosis, cellular inflammatory infiltrate, and hepatocyte necrosis, but does not affect food intake (data not shown). The MCD diet also increased hepatic TG and serum ALT levels. On the other hand, supplementation of MCD diet with EDL markedly alleviated hepatic TG accumulation, high serum ALT levels, hepatic steatosis, and necroinflammation in a dose-dependent manner; in particular, D500 supplementation significantly reduced these factors (Figs. 2 and 3). Thus, EDL protected against liver damage.
GSH-enhancing agents may protect the liver against MCD-induced hepatic oxidative damage. 22 A number of studies have indicated that antioxidants such as polyphenols can prevent or attenuate the extent of lipid peroxidation mainly through free-radical scavenging activity. 23 EDLs contain various bioactive compounds such as luteolin, luteolin-7-O-glucoside, chicoric acid, chlorogenic acid, and total phenol. 8,11 Among these compounds, luteolin, chicoric acid, and the total phenol fraction exhibit remarkable antioxidant activity. 24,25 Our data show that EDL contains luteolin and total phenol. Recently, Park et al. reported that EDL decreased CCL4-induced hepatic MDA formation, possibly due to the presence of luteolin and total polyphenols with antioxidant activity. 13 In the present study, we found that EDL decreased MCD-induced hepatic MDA, a byproduct of free-radical reactions, and attenuated hepatic GSH depletion (Fig. 4). This may have been due to presence of luteolin and polyphenols. Therefore, these findings suggest that EDL may exert antioxidant effects against MCD diet-induced hepatotoxicity.
Apoptosis has been recognized as a prominent feature of liver injuries, and correlates strongly with clinical and histological evidence of disease severity. 26 Regarding this fact, caspases, mainly caspase-3, have been considered as the main causative element in the induction of hepatocyte injuries. 27 We found that MCD elevated caspase-3 levels, whereas EDL treatment significantly suppressed the activation of procaspase-3 to caspase-3 (Fig. 6).
Fatty acids and excessive oxidation lead to the production of radical oxygen species (ROS)-triggered proinflammatory cytokines such as TNF-α and IL-6, which play significant causative roles in the pathogenesis of NASH. 28 TNF-α and IL-6 mediate inflammatory response and apoptosis. 29,30 TNF-α plays a critical role in the evolution of steatohepatitis. 31 IL-6 is another important mediator of inflammatory liver diseases, including obesity-associated fatty liver and cirrhosis. 32,33 In this study, EDL significantly suppressed oxidative stress and dampened the expression of TNF-α and IL-6 (Fig. 5). These results suggest that the anti-inflammatory effects of EDL may be partly related to the inhibition of hepatic lipoperoxide and the mRNA expression of TNF-α and IL-6.
Several studies reported that JNK and MAPK are potently implicated in hepatosteatosis, inflammation, and insulin resistance. 34 –36 The JNK pathway mediates hepatocyte apoptosis and liver injury through oxidative stress, TNF-α, and free fatty acids. 5 Consistent with previous reports, the present study found that MCD led to phosphorylation of JNK, and this activation markedly decreased upon EDL treatment (Fig. 7). Hence, antioxidative EDL might inhibit cellular ROS production and suppress the JNK pathway.
In conclusion, the hepatoprotective effect of EDL might be attributed to its antioxidant activity, which leads to suppression of intrahepatocyte ROS levels, followed by suppression of TNF-α, IL-6, and JNK phosphorylation. EDL holds promise as a therapeutic agent for NASH.
Footnotes
Acknowledgments
This work was supported by the Food Functionality Evaluation Program of the Ministry of Food, Agriculture, Forestry and Fisheries, and by the Korea Food Research Institute, Korea.
Author Disclosure Statement
No competing financial interests exist.