
Abstract
A novel polysaccharide named Angelica sinensis polysaccharide (ASP) was obtained from the powdered and defatted roots of A. sinensis (Oliv.) Diels. The molecular weight of ASP was determined to be 78 kDa and was 95.0% sugars consisting of mostly arabinose, glucose, and galactose with a molar ratio of 1:5.68:3.91. A previous study indicated that ASP may increase plasma iron levels by suppressing the expression of hepcidin, a negative regulator of body iron metabolism, in the liver. The present study aims to clarify the inhibitory effect of ASP on hepcidin expression in rat models of iron deficiency anemia (IDA), and clarify the mechanisms involved. It was demonstrated that ASP significantly reduced hepcidin expression by inhibiting the expression of mothers against decapentaplegic protein 4 (SMAD4) in liver and stimulating the secretion of erythropoietin, which further downregulated hepcidin by repressing CCAAT/enhancer-binding protein α (C/EBPα) and the phosphorylation of signal transducer and activator of transcription 3/5. The results indicate that ASP can suppress the expression of hepcidin in rats with IDA, and may be useful for the treatment of IDA.
Introduction
A
Angelica sinensis (Oliv.) Diels (family Umbelliferae, Chinese name danggui) is a well-known oriental herb used for the treatment of anemia, uterine bleeding, abnormal or painful menstruation, and other diseases affecting women. 3 Many customary applications of the root of A. sinensis are based on empirical observations of its effects on blood tonification and promotion of the blood circulation. 4 As one of the active components, polysaccharide isolated from A. sinensis was found to be effective for enhancing hematopoiesis by increasing the secretion of some hematopoietic growth factors, such as erythropoietin, via stimulating hematopoietic cells and muscle tissues. 5 In our previous research, we found that A. sinensis polysaccharide (ASP) can improve serum iron (SI) status and hypothesized that ASP might participate in the regulation of iron homeostasis. 6
Hepcidin, a 25-amino-acid antimicrobial peptide produced by hepatocytes, is the central regulator of body iron metabolism. 7 Hepcidin binds to ferroportin, the channel for cellular iron efflux, leading to internalization and degradation of ferroportin. The binding diminishes duodenal transfer of iron into the circulation and promotes iron retention within macrophages. 8,9 In this manner, hepcidin lowers plasma iron levels. Chronic elevation of hepcidin levels causes systemic iron deficiency. Therefore, hepcidin has become an attractive therapeutic target for the treatment of iron deficiency diseases. Recently, erythropoietin administration has been used to treat hepcidin-induced anemia since erythropoietin was demonstrated to be effective in suppressing hepcidin expression by enhancing body erythropoiesis. 10
Accordingly, it was proposed that ASP that can stimulate erythropoietin secretion may affect iron homeostasis in vivo by regulating hepcidin expression. The present study was undertaken to evaluate the hepcidin-suppressing efficacy of ASP in IDA rats. Further, to clarify the underlying mechanism of the hepcidin suppression caused by ASP, we monitored the serum concentration of erythropoietin and the liver expression of six proteins belonging to three pathways that were previously demonstrated to be important for hepcidin regulation. 11 These proteins include STAT3/phospho-STAT3 and STAT5/phospho-STAT5 in the Janus-kinase/signal transducer and activators of transcription proteins (JAK/STAT) signal transduction pathway, SMAD4, which is a key protein in Smads family, in the bone morphogenetic protein-mothers against decapentaplegic protein (BMP-SMAD) pathway, and CCAAT/enhancer-binding protein α (C/EBPα) in the erythropoietic pathway.
Materials and Methods
Isolation and characterization of polysaccharides
The dry roots of A. sinensis (Oliv.) were collected by J.-Y.L. in November 2010 at Min County of Gansu Province, China. The material was identified at the Department of Traditional Chinese Medicine, Tongji Medical College of Huazhong University of Science and Technology. A voucher specimen (number 78) was deposited in the herbarium of the Tongji Medical College. Two hundred grams of powdered material was extracted twice with boiled distilled water (1000 mL) for 0.5 h. The filtered extract was combined and concentrated under reduced pressure at 50°C. After removing acid and alkaline proteins by modulating pH, the aqueous extract was precipitated with ethanol (final concentration 75% [v/v]). The resulting precipitate was dissolved in distilled water and then applied to a Sephadex G-50 column (3.0 cm×35 cm; Pharmacia), eluted with distilled water. The main fraction named ASP was collected and lyophilized (yield: 2.87%).
The molecular weights of the polysaccharides were determined by high-performance size exclusion chromatography using an Agilent-LC 1100 instrument (Agilent, Santa Clara, CA, USA), equipped with a TSKGEL G2000 PWXL column and eluted with 0.05 M Na2SO4 solution at a flow rate of 0.8 mL/min. Elution was monitored by an Agilent refractive index detector. A series of solutions made from standard dextrans were run under the same conditions; a standard curve linear over a wide range (0.2–2000 kDa) was obtained by plotting the elution volume versus the logarithm of the corresponding molecular weight (Supplementary Figs. S1 and S2; Supplementary Data are available online at
The percentage of total sugar was estimated by the phenol–sulfuric acid method. 12 ASP was hydrolyzed with 1 M H2SO4 and converted to their alditol acetates. The resulting alditol acetates were analyzed by gas chromatography (GC) using an Agilent 7890A instrument (Agilent), equipped with a HP-5 fused-silica capillary column (30 m×0.32 mm×0.25 μm) and an Agilent 5975CMS detector.
Iron-deficient diet
Referencing the formulation recommended by American Association of Analytical Chemists (AOAC), 13 iron-deficient diet was prepared using corn starch (corn flour and corn contour slag mixed) as its major components. Formula was as follows: corn starch about 99%, salt about 0.7%, choline chloride about 0.1%, mixed Vitamin ∼0.1%, trace elements about 0.1%, and the content of iron was 10.4 mg/kg determined by flame atomic absorption spectrometry.
Rat model and treatment
Establishment of IDA model
Male Sprague Dawley rats (250±20 g) were obtained from the Experimental Animal Center of Huazhong University of Science and Technology, housed in clear plastic cages and kept under controlled conditions, temperature (22°C±2°C), humidity (50%±10%), and 12-h day per light cycle (lights on at 7:00 a.m.). All animals were allowed to acclimate for 7 days prior to beginning the study. Experimental IDA was established by feeding an iron-deficient diet combination with repeated bloodletting from the orbital vein plexus three times a week. Rats had free access to food and distilled water from an iron-free apparatus that was used to preclude extraneous iron intake from the water. The entire experimental procedure was strictly controlled to avoid iron contamination.
Experimental groups and drug administration
Thirty-six IDA rats were randomly assigned into three groups of 12 animals each: ASP treatment group was administered intragastrically with ASP (1 g/kg) each day for 14 days. 14 RhEPO treatment group was given recombinant human erythropoietin (rhEPO; Sunshine Pharmaceutical Co., Shenyang, China) by intraperitoneal injection at a dose of 2000 U/kg each day for 3 days. Negative control group was administered intragastrically with 1 mL normal saline each day for 14 days. Dosages and time were determined according to preliminary experiments. All experiments were done in accordance with the procedures approved by the University Animal Care and Ethics Committees (approval number: SCXK2004-0007).
Blood collection (for evaluating hepcidin-suppressing effect of ASP and rhEPO)
Seven rats in each group were used to investigate the suppression of hepcidin levels by ASP and rhEPO. Animals were anesthetized (ketamine 70 mg/kg and acepromazine 3 mg/kg, intraperitoneally) and blood was taken from the retro-orbital sinus before administration and 0, 1, 2, 4, 8, 12, 24, and 48 h after the last administration.
Liver collection (for mechanistic study of hepcidin suppression by ASP and rhEPO)
The other five rats of each group were used to investigate the expression of six related signal proteins in liver. Livers were taken 8 h (rhEPO group) or 24 h (ASP and control groups) after the last administration, and kept at −80°C.
Analytical procedures
SI and hematology
Hematology was performed in blood collected with EDTA as an anticoagulant using a blood cell automatic analyzer (Model CD-1700; Abbott Co., Santa Clara, CA, USA). SI was measured using a flame atomic-absorption spectrometer (Model AA-240 FS, VARIAN Co., Palo Alto, CA, USA).
Enzyme-linked immunosorbent assay
Blood samples were centrifuged at 2500 g for 10 min after coagulation. Supernatant serum was stored at −20°C. Erythropoietin and hepcidin in serum were determined by enzyme-linked immunosorbent assay according to the manufacturer's instructions (USCN Life Co., Missouri, TX, USA).
Western blot analysis
Liver tissue lysates were prepared as follows: minced liver samples were lysed in a pH 7.5 buffer containing 50 mM HEPES, 150 mM NaCl, 1 mM EDTA, 5 mM EGTA, 10% glycerol, 0.1 mM phenylmethylsulfonyl fluoride, and protease inhibitor mixture (Roche Diagnostics, Indianapolis, IN, USA). Protein concentrations were determined using a Bio-Rad protein assay kit. Total cellular proteins (100 μg per lane) were resolved with sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to nitrocellulose membrane (Amersham Biosciences, Uppsala, Sweden). Membranes were blocked in 5% nonfat milk in Tris-buffered saline (TBS), incubated in primary antibody washed and incubated with peroxidase-conjugated secondary antibody, washed, and then incubated with chemiluminescence substrate (ECL Plus). The signal was visualized using X-ray film in dark room. The relative and normalized protein expression was calculated by a ratio of density of each estimated protein to density of β-actin.
The antibodies used in the Western blot assay were rabbit polyclonal directed against pY705-STAT3 (1/1000; No. 44380G, Invitrogen, Carlsbad, CA, USA), mouse monoclonal directed against STAT3 (1/1000; No. 9139, Cell Signaling Technology, Boston, MA, USA), rabbit monoclonal directed against pT694-STAT5 (1/1000; No. 9359, Cell Signaling Technology), rabbit polyclonal directed against STAT5 (1/1000; No. 9363, Cell Signaling Technology), mouse monoclonal directed against Smad4 (1/50; No. 39-1200, Invitrogen), rabbit monoclonal directed against C/EBP α (1/1000; No. ab40761, Abcam, Cambridge, United Kingdom), mouse monoclonal directed against β-actin (1/2000; Sigma, St. Louis, MO, USA), peroxidase-conjugated goat anti-rabbit immunoglobulins (1/4000; No. KC133423, Thermo, Varian, Palo Alto, CA, USA), peroxidase-conjugated goat antimouse immunoglobulins (1/4000; No. LA142254, Thermo, Varian).
Statistical analysis
Results were expressed as mean±S.D. Statistical analyses were performed using SPSS version 13.0 software. Differences between groups were assessed by one-way analysis of variance and least significant difference to judge whether the results were statistically significant. The critical P value of both tests was set to 0.05.
Results
Isolation and characterization of polysaccharides
A novel polysaccharide named ASP was obtained from the powdered and defatted roots of A. sinensis (Oliv.) Diels with a molecular weight estimated to be 78 kDa and 95% total sugars. From the chemical analysis, the component sugars of ASP, determined by GC, were arabinose, glucose, and galactose with a molar ratio of 1:5.68:3.91.
Establishment of IDA
Rats began to exhibit symptoms of IDA 2 weeks after modeling, such as pale skin, rough and sparse fur, as well as slow growth. Additional symptoms, such as body weight loss, fur disluster, and alopecia, were observed over time. The ears and paws were noticeably pale and animals displayed listlessness, decreased activity, unresponsiveness, and scrunching. As shown in Table 1, hemoglobin, hematocrit, mean corpuscular volume, mean corpuscular hemoglobin (MCH), MCH concentration, and SI were significantly decreased (P<.01) but red blood cell distribution width markedly increased (P<.05) when compared with those observed before modeling. These data collectively indicated that the IDA model was established successfully.
Iron deficiency anemia was seen in rats by 6 weeks after initiation of the iron-deficient diet combined with repeated bloodletting from the orbital vein plexus three times a week. The data are expressed as mean±SD of 36 animals.
P<.05, ** P<.01 as compared to the values measured before modeling.
Hb, hemoglobin; RBC, red blood cell numbers; HCT, hematocrit; MCV, mean corpuscular volume; MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; RDW, red blood cell distribution width; SI, serum iron; BW, body weight; EPO, erythropoietin.
After the IDA model was established, EPO concentration in rats' serum was significantly increased by 60.3% and hepcidin levels were significantly decreased by 29.4% through a long-term self-regulation. Hepcidin expression was downregulated by iron deficiency, anemia, and hypoxia, it in turn promoted intestinal iron absorption and monocyte-macrophage system (MMS) iron release, to meet the iron demand of heme synthesis.
Erythropoietin promotion and hepcidin suppression
After 3-day treatment of rhEPO and 14-day treatment with ASP, hepcidin levels were significantly reduced by 13.4% and 31.8%, respectively (Fig. 1). In the negative control group, EPO and hepcidin levels were not significantly changed at each time point as compared to the treatment group (Supplementary Fig. S3). In both ASP and rhEPO groups, erythropoietin levels in serum rose significantly, and after 8–12 h, hepcidin levels began to decrease noticeably (Fig. 1). After the last administration of ASP, serum erythropoietin reached the maximum level at 12 h and went back to the initial level at 24 h; serum hepcidin began to decrease at 12 h and reached the minimum level at 24 h (Fig. 1B). However, in rhEPO group (Fig. 1A), the changes in both erythropoietin and hepcidin levels progressed more quickly than those in ASP group. It could be inferred that the maintenance of the high level of erythropoietin after ASP administration might result in consistently low levels of hepcidin.
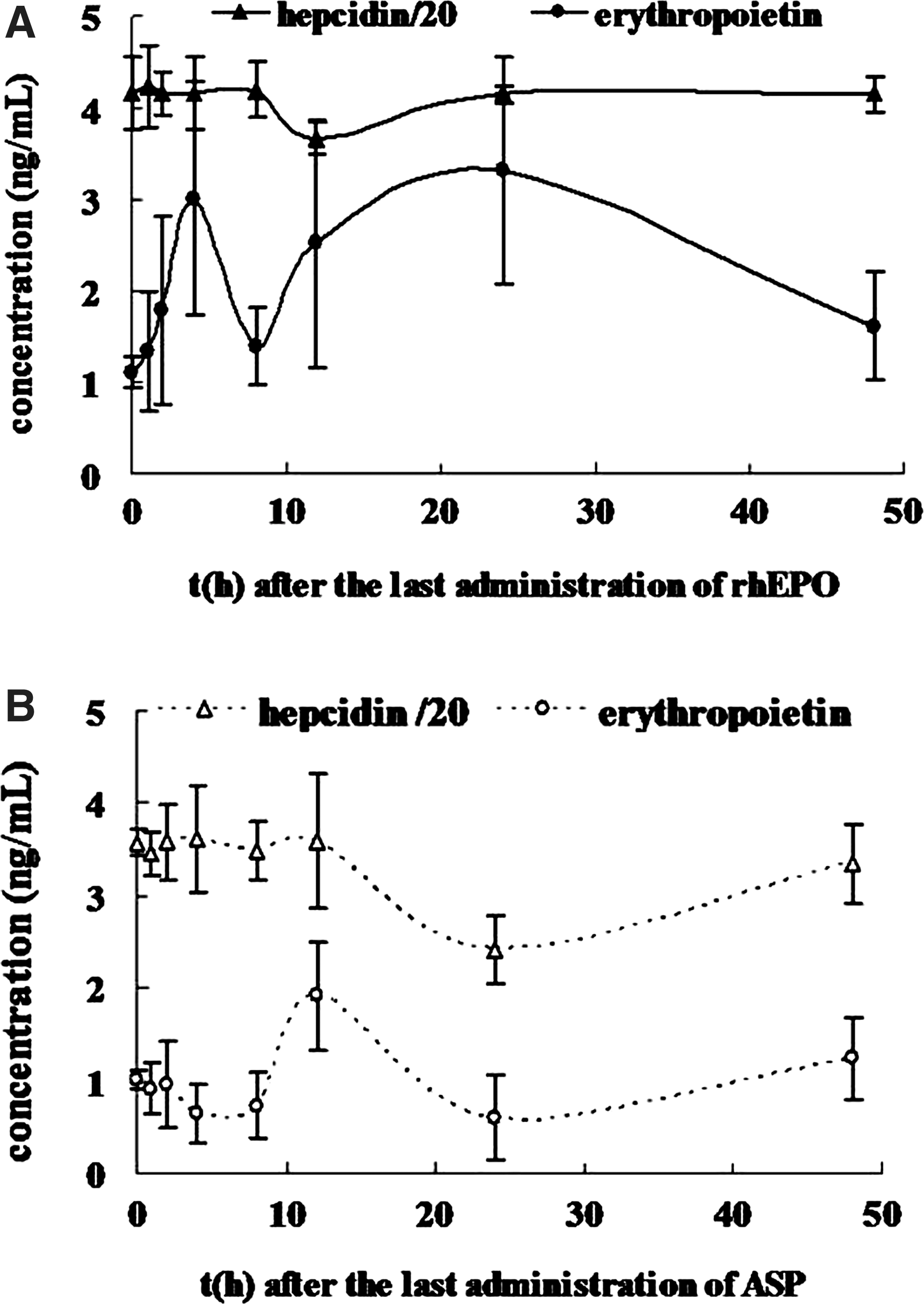
Erythropoietin promotion and hepcidin suppression induced by recombinant human erythropoietin (rhEPO) and Angelica sinensis polysaccharide (ASP).
Inhibition of signal proteins involved in hepcidin regulation
STAT3/phospho-STAT3 and STAT5/phospho-STAT5 expression
ASP administration reduced the mean levels of hepatic phospho-STAT3 and phospho-STAT5 by 54.4% and 60.6%, respectively (Fig. 2). In the rhEPO group, the mean levels of phospho-STAT3 and phospho-STAT5 were reduced by 62.8% and 81.8%, respectively. Nevertheless, the expression of STAT3 and STAT5 was not affected by ASP and rhEPO. There were significant decreases in the ratio of phospho-signal transducer and activator of transcription 3/5 (STAT3/5) and STAT3/5, indicating that both ASP and rhEPO inhibited the phosphorylation of STAT3/5 rather than the levels of the proteins themselves.
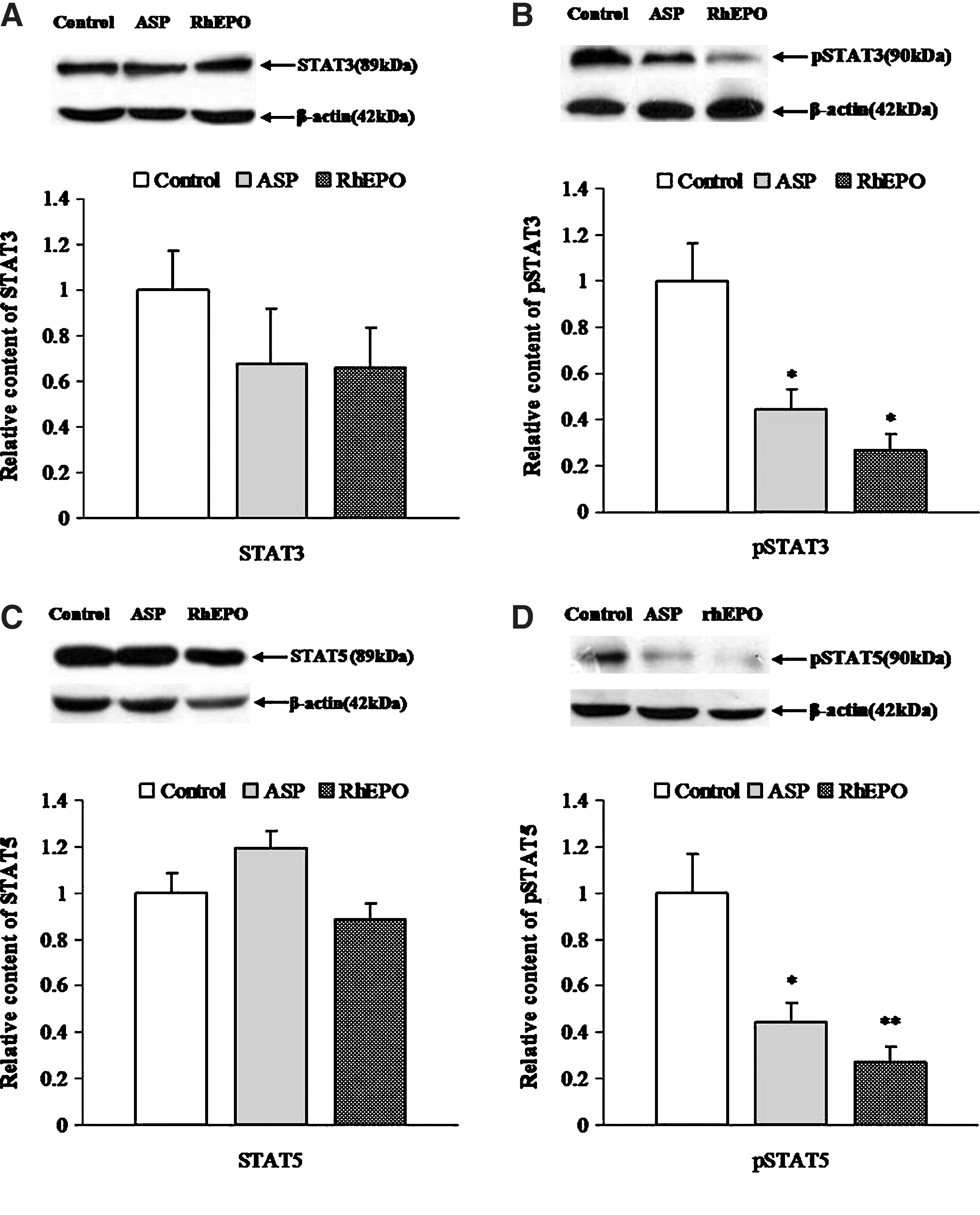
Suppression of Janus-kinase/signal transducer and activators of transcription proteins (JAK/STAT) by ASP and rhEPO.
SMAD4 expression
Previous reports showed that SMAD4 is the key protein in BMP-SMAD pathway and is essential for hepcidin production in response to several stimulants. 15 Western blot analysis of SMAD4 indicated that the expression of SMAD4 in liver tissues from ASP group was significantly decreased by about 64.7%, but there was no significant difference in rhEPO group (Fig. 3).
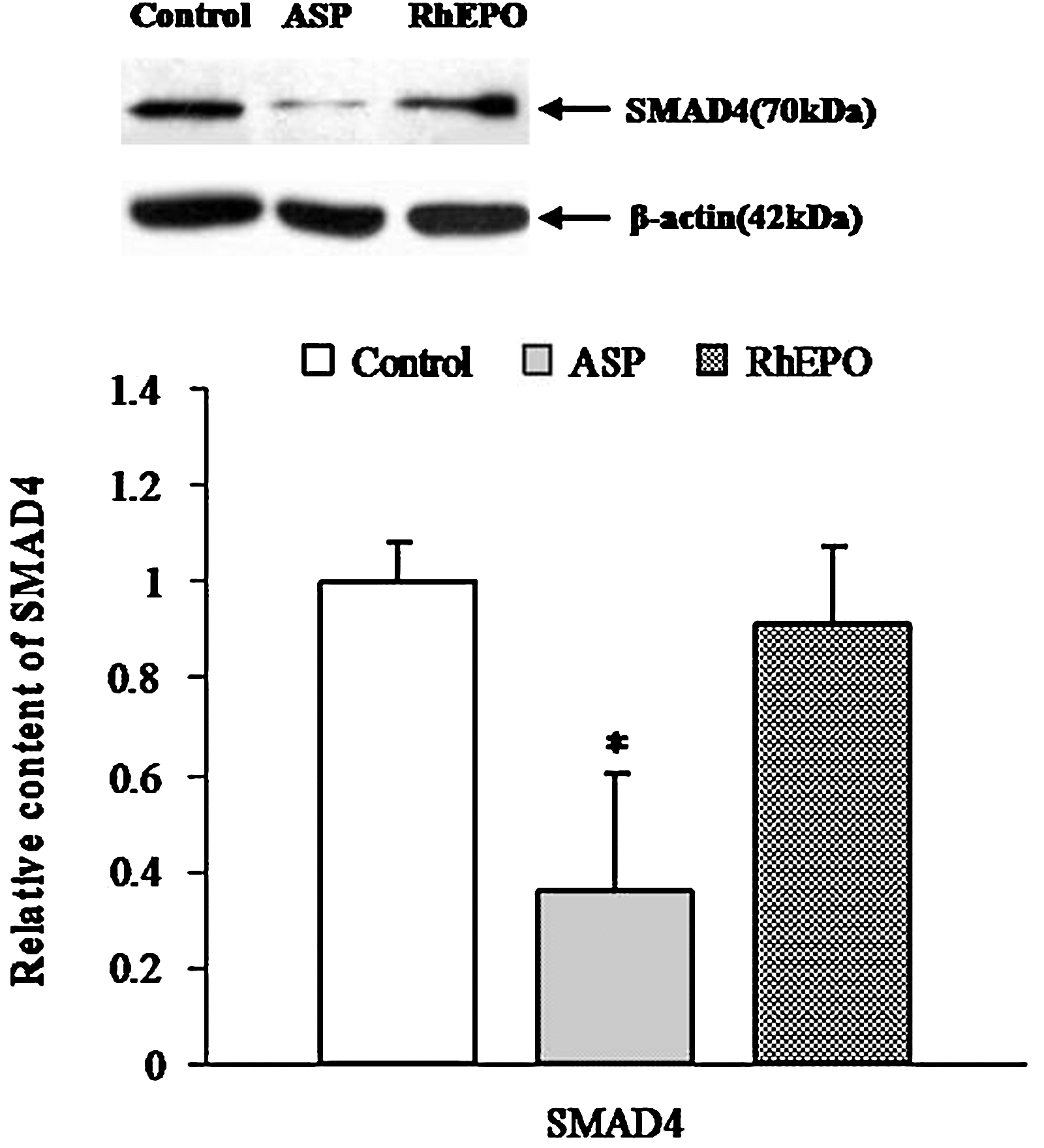
Suppression of SMAD4 by ASP and rhEPO. SMAD4 is a key protein in the SMADs family, which is a group of TGF-β signal-specific intracellular signal transducers. Bars represent mean±SD of ADU following Western blot analysis. Values are expressed as percentages relative to a negative control group. Control: rats (n=5) administered normal saline (1 mL) for 14 days; ASP: rats (n=5) administered ASP (1 g/kg) for 14 days; RhEPO: rats (n=5) administered rhEPO (2000 U/kg) for 3 days. *P<.05 as compared with control.
C/EBPα expression
C/EBPα was the first transcription activator of the hepcidin gene identified. Its importance was supported by the fact that liver-specific deletion of C/EBPα gene in mice prevented hepcidin expression and induced a severe iron overload. 16 In this study, C/EBPα levels were reduced by 49.4% and 75.9%, respectively, after ASP and rhEPO treatment (Fig. 4).
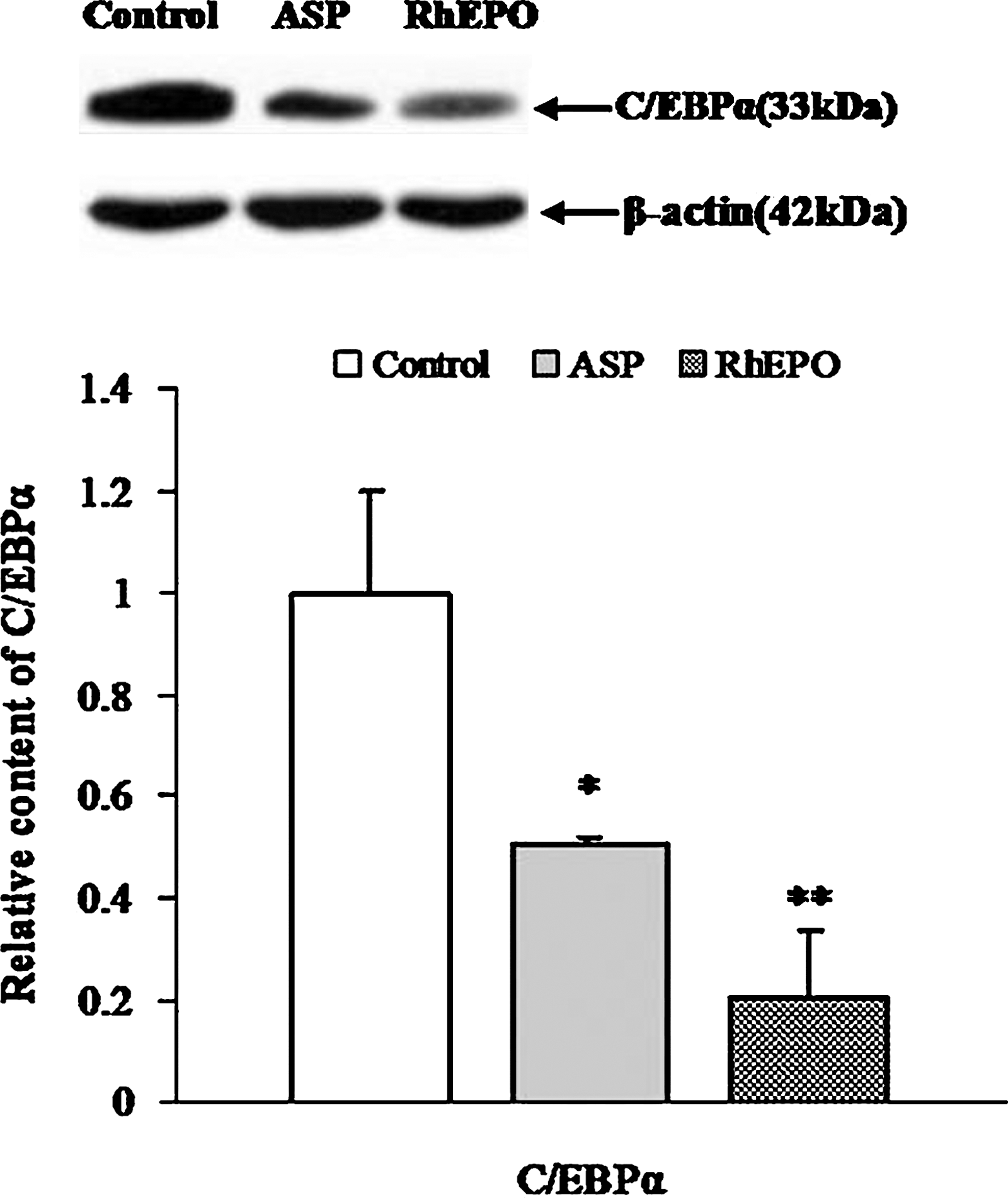
Suppression of CCAAT/enhancer-binding protein α (C/EBPα) by ASP and rhEPO. Bars represent mean±SD of ADU following Western blot analysis. Values are expressed as percentages relative to negative control group. Control: rats (n=5) administered normal saline (1 mL) for 14 days; ASP: rats (n=5) administered ASP (1 g/kg) for 14 days; RhEPO: rats (n=5) administered rhEPO (2000 U/kg) for 3 days. *P<.05, **P<.01 as compared with control.
Discussion
Hepcidin, mainly produced by the liver, is the principal negative regulator of iron homeostasis. Studies in transgenic mice showed that hepcidin deficiency leads to iron overload and that overexpression of hepcidin leads to severe iron deficiency and anemia. Thus, the discovery of hepcidin inhibitor will bring about new breakthroughs in the treatment of diseases induced by hepcidin overexpression. Recent research indicated that erythropoietin can directly combine with erythropoietin receptor on the cell surface, which would cause the intracellular signal transduction to suppress the expression and DNA binding activity of transcription factor C/EBPα and phospho-STAT3/5, thereby inhibiting the expression of hepcidin. 17 However, because of the short half-life of erythropoietin in serum, frequent injection of high doses of erythropoietin is needed, resulting in poor tolerance and expensive cost in patients.
In this study, we investigated the effect of ASP on hepcidin suppression in IDA rats, as well as its related mechanism. The results showed that ASP can promote erythropoietin secretion and decrease hepcidin levels in IDA rats. Consistent with earlier reports, 18,19 we demonstrated that rhEPO administration suppresses hepcidin expression. Interestingly, no variation in hepcidin levels was observed when erythropoietin levels reached its maximum; however, hepcidin concentrations began declining when erythropoietin levels fell back. This phenomenon demonstrated that the directly inhibiting factor of hepcidin expression was not erythropoietin, 18,20 but the downstream molecules of erythropoietin. This speculation was consistent with literature that supports the concept that the decrease of hepcidin following repeated administration of erythropoietin is secondary to the effect of erythropoietin on erythropoiesis. Comparing the duration of hepcidin suppression in the two treatment groups, it could be noticed that the rats' hepcidin concentration stayed low for a longer time in the ASP-treated group, implicating the prolonged action of ASP in hepcidin suppression.
Studies conducted during the last few years indicate that the regulation of hepcidin expression is a complex process. 21 It is becoming increasingly clear that hepcidin gene transcription is influenced by three main pathways: the inflammatory pathway, which increases hepcidin transcription through JAK/STAT signal transduction in response to inflammatory mediators; the BMP-SMAD pathway, which mediates hepcidin upregulation by iron and hypoxia; and the erythropoietic pathway, which decreases hepcidin mRNA levels following an increase in the rate of erythropoiesis. 11 Accordingly, inhibitory effects of erythropoietin and ASP on key proteins in these pathways were investigated to clarify the molecular mechanism involved in hepcidin suppression.
Western blot analysis showed that ASP and rhEPO can significantly downregulate phospho-STAT3 and phospho-STAT5 without affecting STAT3 and STAT5, suggesting that ASP and rhEPO may affect Janus-kinases that phosphorylate STATs. Therefore, it could be speculated that the slight inhibitory activity of ASP on the phosphorylation process is attributable to the effect of erythropoietin stimulated by ASP.
In hepcidin regulation, BMP signaling can enhance the phosphorylation of SMAD1/5/8. 22 Then, SMAD4 is needed to bind these activated SMADs and the complex moves to the nucleus where it can stimulate hepcidin expression. As a further support for the role of SMAD4 in the regulation of hepcidin expression, ablation of SMAD4 specifically in the liver triggers an iron overload in multiple organs due to decreased levels of liver hepcidin. 15 The results presented in this study suggest that ASP can suppress hepatic SMAD4 expression and may interrupt the entire BMP-SMAD pathway.
Pinto et al. recently proposed that erythropoietin could act directly to suppress hepcidin in hepatocytes through the regulation of the transcription factor C/EBPα. Similarly, we found that erythropoietin can significantly inhibit hepatic C/EBPα in IDA rats. Nevertheless, the suppression of C/EBPα by ASP administration was to a less extent than that by rhEPO administration. This may be due to the lower levels of erythropoietin in blood promoted by ASP than that by rhEPO. These results also indicated that erythropoietin stimulated by ASP could take part in the hepcidin suppression of ASP.
Recent researches clarified that erythropoietin can inhibit hepcidin expression directly through the inhibition of C/EBPα in vitro 17 and indirectly via the suppression of STAT3/phospho-STAT3 signaling in vivo. 18 Similarly, we confirmed that erythropoietin can interrupt the two signal transduction ways to suppress hepcidin expression in IDA rats. In addition, the STAT5/phospho-STAT5 pathway was also affected by erythropoietin in our study. It is proposed that erythropoietin mediates hepcidin by inhibiting C/EBPα and the phosphorylation process of STAT3/5 (Fig. 5).
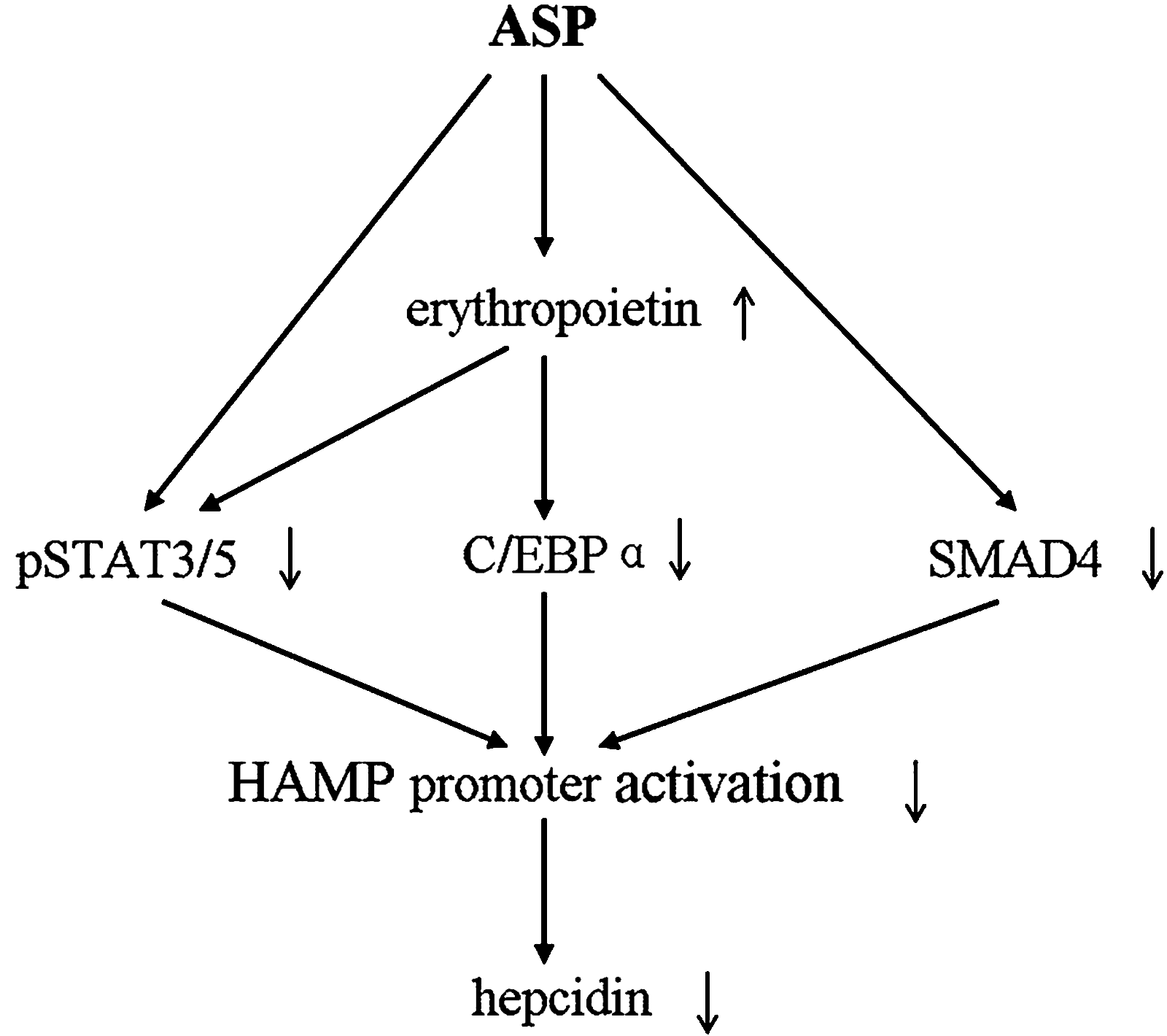
Molecular mechanism speculated to be involved in hepcidin (HAMP) suppression by ASP. ASP downregulates the transcription regulators pSTAT3/5 and SMAD4, and stimulates endogenous erythropoietin secretion that further decreases the signal proteins in liver. As a result, hepcidin expression was suppressed. Erythropoietin inhibits the phosphorylation of pSTAT3/5 and the expression of C/EBPα, which causes the reduction of activating factors and the suppression of hepcidin.
Effects of Angelica polysaccharides on blood promotion and erythropoiesis activation have been already confirmed by many previous studies. 3,23 We further demonstrated its activity in the stimulation of endogenous erythropoietin secretion as well as the positive correlation between erythropoietin promotion and hepcidin repression. Therefore, it is speculated that the stimulation of erythropoietin as well as the inhibition of phospho-STAT3/5 and SAMD4 are involved in the ASP-induced suppression of hepcidin expression (Fig. 5).
In conclusion, results from the present study showed that ASP can suppress hepcidin expression in vivo by stimulating erythropoietin secretion and interrupting two other major pathways of hepcidin regulation. This finding supports the hypothesis that ASP could participate in the regulation of iron homeostasis. As a hepcidin inhibitor, both ASP and rhEPO could be utilized in the treatments of diseases caused by hepcidin overexpression, such as IDA. Moreover, because of its safety, various bioactivities, 24 and convenience in administration, ASP would be more widely used than rhEPO. Further work will employ other models of hepcidin overexpression to characterize the effect of ASP on hepcidin expression and the mechanisms continue to be more fully elucidated.
Footnotes
Acknowledgment
This research was supported by a grant from National Natural Science Foundation of China (30973910).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.