
Abstract
This study examined the effect of fermentation on the ability of antler to act as a stimulator of hematopoietic activity. Hemolytic anemia was induced by phenylhydrazine (PHZ) in female Sprague–Dawley rats. The vehicle or antler extract (nonfermented or fermented) mixed in drinking water was administered from Days 2 to 15 after PHZ injection. On Day 15, red blood cell counts in the fermented antler group (6.33×106/μL) were significantly higher than those in the nonfermented antler group (5.90×106/μL) (P<.05), and rats treated with fermented antler extract tended to have higher hemoglobin compared with rats treated with nonfermented antler extract, but not significantly. In addition, rats treated with fermented antler extract had slightly lower serum erythropoietin levels compared with nonfermented antler extract, which were not statistically different from serum erythropoietin levels of nonanemic rats. We conclude therefore that the hematopoietic activity of antler might be increased by the fermentation process.
Introduction
T
For more than 2000 years, deer and deer parts have been sources of medicine in Asian countries such as China, Taiwan, Mongolia, and Korea. The first report of antler use in medicine appeared on a silk scroll unearthed from a Han tomb in China dated 100 BC. Since that time, antler has been used as a popular medicine in these countries. 5 There are many medical and pharmaceutical uses of antler, which is believed to possess restorative and anti-aging properties, provide sexual support for sexual disorders in men, aid in menstrual disorders and menopause in women, promote rapid healing, treat weight loss, improve growth in children, strengthen weak bones, and bestow hematopoietic activity. 6 –11
Antler potentiates the recovery of RBCs, serum erythropoietin (EPO) levels and the hepatic activity of δ-aminolevulinic acid dehydratase (ALAD) in animal models of hemolytic anemia. 12 The increased activity would partly support the stimulating effect of antler on hematopoietic activity. Finally, with antler, a lower occurrence of anemia and a faster recovery from anemia have been reported. 13 Other studies also support the view that antler improves hemolytic anemia in many species. 14,15
To increase the beneficial activity of antler, we tried fermentation, which may have merits for standardizing efficacy. Fermentation by microorganisms is used to make products useful to humans, including various foods. Fermentation in food processing usually implies that the action of microorganisms is desirable, and the process is used to produce alcoholic beverages such as wine, beer, and cider. Fermentation is also used in the leavening of bread (CO2 produced by yeast activity) and for food preservation with lactic acid in sour foods such as sauerkraut, dry sausages, kimchi, and yogurt or with vinegar (acetic acid) for use in pickling foods. The primary benefit of fermentation is the conversion of sugars and other carbohydrates into other molecules (e.g., converting juice into wine, grains into beer, carbohydrates into CO2 to leaven bread, and sugars in vegetables into preservative organic acids). 16 Food fermentation has been said to serve five main purposes: enrichment of the diet through development of a diversity of flavors, aromas, and textures in food substrates; preservation of substantial amounts of food through lactic acid, alcohol, acetic acid, and alkaline fermentations; biological enrichment of food substrates with protein, essential amino acids, essential fatty acids, and vitamins; elimination of antinutrients; and a decrease in cooking times and fuel requirements. So fermentation increases the absorption rate and has merits for standardizing efficacy. 16,17 In our previous studies, we demonstrated that bone growth, immunomodulating, and antioxidant activities of antler might be increased through the fermentation process. 18,19 The present study was performed to examine the effect of fermentation on the ability of antler to stimulate hematopoietic activity. We evaluated the effect of fermented antler on the changes of hematopoietic factors, such as hematological parameters, EPO, ALAD activity, and osmotic hemolysis induced by PHZ exposure in rats.
Materials and Methods
Fermented antler extract preparations
Dried antler (Cervi Parvum Cornu) from adult male Russian elks was obtained from Kwang-Dong Pharmaceutical (Gyeonggi-do, Korea). The samples were a combination of three sections: top (37.5%), middle (37.5%), and tip (25%). Our previous study was conducted to isolate strains for the preparation of fermented antler and evaluate its physiological activities. From that study, Bacillus subtilis KH-15 was selected among 21 samples from Bacillus sp., Lactobacillius sp., and mushroom strain. 18 B. subtilis KH-15 isolated from soybean paste was grown on LB broth at 30°C for 1 day. The stock culture was maintained on potato dextrose agar slants and subcultured every month. The slants were incubated at 25°C for 7 days and eventually stored at 4°C. Dried antler was initially grown on potato dextrose agar medium in a Petri dish. It was transferred to the seed culture medium by punching out 5-mm disks of the agar plate culture with a sterilized self-designed cutter. The seed culture was grown in a 250-mL flask containing 50 mL of potato dextrose broth at 25°C on a rotary shaker incubator at 150 rpm for 5 days. Flask cultures were incubated in 500-mL flasks containing 100 mL of antler powder medium (15 g of antler powder in 100 mL of distilled water) at 25°C on a rotary shaker at 150 rpm for 5 days. Alternatively, cultures were incubated in a 3-L stirred-tank fermenter (Fermentech, Gyeonggi-do, Korea) containing 1 L of antler powder medium (24 g of potato dextrose broth, 20 g of agar, and 300 g of antler powder in 1 L of distilled water) after inoculation with 2% (vol/vol) flask culture for 7 days (temperature, 25°C; aeration rate, 1 vol/vol/min; agitation speed, 150 rpm; initial pH 5.6). After fermentation, the ferment mass was adjusted to 300 mL, and it was extracted twice at 100°C for 3 h. The combined extract was centrifuged at 2800 g for 20 min. The supernatants were concentrated using a vacuum evaporator at 40°C and lyophilized to produce the fermented antler extract. Nonfermented antler extract was prepared by soaking 45 g of antler powder in 300 mL of distilled water followed by refluxing for 3 h and cooling. The extracts were centrifuged, concentrated, and lyophilized.
Experimental protocol
The experimental protocol was reviewed and approved by the Korea University Animal Care Committee. Female Sprague–Dawley rats were obtained at 4 weeks of age from Daehan Biolink (Chungchungbuk-do, Korea). Hemolytic anemia is more common in females than in males; females are about twice as likely to be anemic as males. 1 The animal room was maintained at 24±1°C with 60% atmospheric humidity and a 12-h light/12-h dark cycle. Eight rats were kept as a normal control group (control), whereas anemia was induced in 32 rats by intraperitoneal injection of PHZ (40 mg/kg of body weight) daily for 2 days. Rats that developed anemia with hemoglobin (Hgb)<14 /dL were used for the study. Anemic rats were randomly divided into four groups (n=8 rats per group) and treated as follows: PHZ-con, the equal volume of normal saline (0.9% NaCl) as a vehicle; Non-200, non-fermented antler extract (200 mg/kg of body weight); B-100, fermented antler extract (100 mg/kg of body weight); and B-200, fermented antler extract (200 mg/kg of body weight). The saline or antler extract was administered orally from Days 2 to 15 after PHZ injection (for 14 days).
Hematological parameters
On Day 0 (before anemia induction), 2 (after anemia induction and before treatment), and 15 (after treatment), blood samples were taken from the tail veins of prewarmed rats to determine blood biomarkers. Heparin-added whole blood samples, immediately after collection, were used for hematological analyses. The serum was separated by centrifugation at 2800 g for 15 min at 4°C and was then stored at −70°C until analysis. Hematological analysis was performed using a model KN-21N automatic hematological analyzer (Sysmex, Seoul, Korea). Parameters included RBC count, white blood cell (WBC) count, hematocrit (Hct), Hgb, mean corpuscular volume (MCV), mean corpuscular Hgb (MCH), MCH concentration (MCHC), platelet count, and reticulocyte count.
EPO analysis
Serum EPO level was determined using a rat EPO enzyme-linked immunosorbent assay kit (catalog number E0028r; Uscn Life Science & Technology, Beijing, China). For renal EPO gene expression, the medullary region of the kidney was removed, and only the cortical tissue was used for the analysis. 20 RNA isolation and reverse transcriptase–polymerase chain reaction analysis were performed according to the procedure of Gazzaniga et al. 21 Primer sequences used to amplify the desired cDNA were EPO forward and reverse primers: 5′-CACGAAGCCATGAAGACAGA-3′ and 5′-GGCTGTTGCCAGTGGTATTT-3′.
ALAD activity
The hepatic ALAD activity was assayed according to the procedure of Berlin and Schaller.
22
The livers were removed, and their wet weights were subsequently measured. They were then washed in 0.15 M NaCl solution, blotted onto filter paper, weighed, and homogenized in 4.5 mL of potassium phosphate buffer (pH 7.0). The liver homogenate was centrifuged at 18,928 g for 4 min at 4°C, and the supernatant was used as a liver tissue solution. In brief, 0.2 mL of a liver tissue solution was mixed with 1.3 mL of distilled water and incubated for 10 min at 37°C for complete hemolysis. After addition of 1 mL of standard aminolevulinic acid, the tubes were incubated for 60 min at 37°C. Enzyme activity was stopped after 1 h by adding 1 mL of 10% trichloroacetic acid. After centrifugation of the reaction mixture at 1008 g for 10 min, an equal volume of Ehrlich reagent was added to the supernatant, and the absorbance was determined at 555 nm after 5 min. The hepatic ALAD activity was calculated by the following equation:
where Abs sample is the absorbance in the presence of sample and Abs control is the absorbance in the absence of sample, respectively.
Osmotic resistance
The osmotic resistance of RBCs was evaluated according to the method of Diallo et al. 23 Solutions varying in saline concentration from 0% to 9% were prepared, and 50 μL of blood was mixed with each saline solution. The mixture was incubated for 60 min. After the incubation, the mixture was centrifuged at 448 g for 10 min. The supernatant was collected, and the absorbance was determined at 540 nm.
Statistical analyses
All statistical analyses were performed using SPSS version 12.0 (SPSS, Inc., Chicago, IL, USA). The differences among groups were evaluated statistically by one-way analysis of variance and Tukey's multiple tests. All data were two-sided with a 5% significance level and were reported as mean±SD values.
Results
Weight gain and food intake
Body weight gain and food intake recorded during the experimental period are shown in Table 1. Food intake did not differ among all groups. Despite identical food intake, anemic rats (PHZ-con, Non-200, B-100, and B-200 groups) gained significantly less weight than nonanemic rats (control group) (P<.05). Hemolytic anemia decreased the weight gain of rats, but treatment with antler extract, especially fermented antler extract, tended to suppress this decrease. The weight gains of B-100 and B-200 groups were as high as that of the control group.
Data are mean±SD values for eight rats per group.
Means with different superscript letters are significantly different at P<.05 by Tukey's multiple comparison test.
Control, normal rats; PHZ-con, anemic rats treated with phenylhydrazine (PHZ) vehicle; Non-200, anemic rats treated with nonfermented antler extract (200 mg/kg of body weight); B-100, anemic rats treated with fermented antler extract (100 mg/kg of body weight; B-200, anemic rats with treated fermented antler extract (200 mg/kg of body weight).
Hematological analysis
We examined the effect of fermented antler extract on hematological parameters in anemic rats. The results are summarized in Figure 1 and Table 2. PHZ-induced hemolytic anemia was accompanied by changes of hematological parameters. The PHZ-con group showed significant decreases in RBC, Hct, and Hgb levels, indicating anemia, compared with the control group (P<.05) (on Day 15). Antler extract moderated these PHZ-induced anemia-related hematological changes, which were slightly more pronounced using fermented antler extract than using nonfermented antler extract. In particular, the RBC level in the fermented antler group was significantly higher than that in the nonfermented antler group (P<.05), and rats treated with fermented antler extract had slightly higher Hgb compared with the treatment of nonfermented antler extract; the B-200 group did not display Hgb levels statistically different from those of the control group (P>.05) (on Day 15). PHZ-induced hemolytic anemia also induced increases of MCV, MCH, MCHC, and reticulocyte levels; these hematological parameters were significantly higher in the PHZ-con group than in the control group (P<.05) (on Day 15). When rats were treated with antler extract, the increases in MCV, MCH, MCHC, and reticulocyte levels induced by PHZ exposure were suppressed, and the fermentation accelerated the activity of antler on the improvement of these hematological parameters. MCV, MCH, MCHC, and reticulocyte levels in fermented antler groups were slightly lower than those in the nonfermented antler group. However, we did not observe significant differences in WBC and platelet levels among all groups.
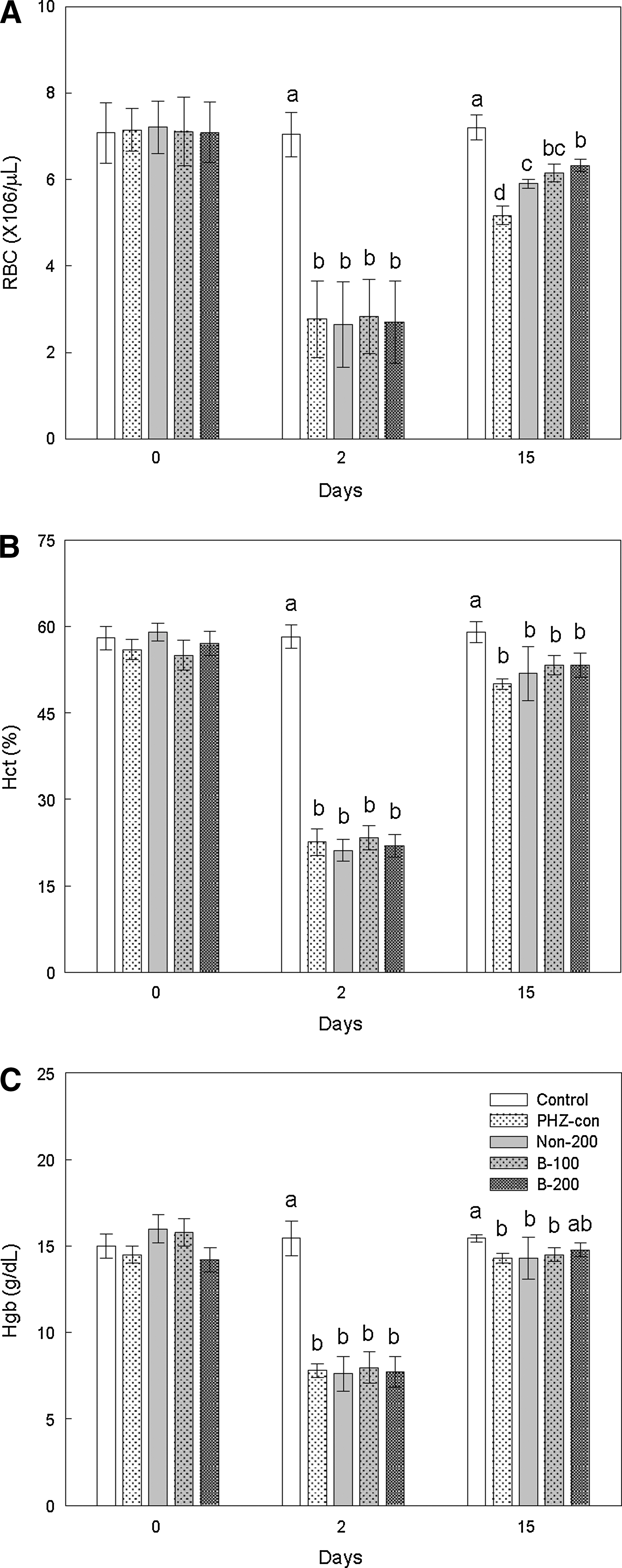
Effect in PHZ-induced anemic rats of fermented antler extract on hematological parameters:
Data are mean±SD for eight rats per group.
Means with different superscript letters are significantly different at P<.05 by Tukey's multiple comparison test.
See Table 1 for the description of groups. MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; MCV, mean corpuscular volume; WBC, white blood cell.
Serum EPO level and renal EPO gene expression
We evaluated serum EPO levels and renal EPO gene expression using reverse transcriptase–polymerase chain reaction analysis in anemic rats. As shown in Figure 2, PHZ-induced hemolytic anemia significantly increased serum EPO levels (control vs. PHZ-con, 2.70 ng/mL vs. 6.33 ng/mL; P<.05). In addition, antler extract suppressed the increase in serum EPO induced by PHZ exposure. The serum EPO levels of antler-treated groups tended to be lower than that of the PHZ-con group. Moreover, rats treated with fermented antler extract (B-100 and B-200 groups, 4.67 ng/mL and 4.03 ng/mL, respectively) had slightly lower serum EPO levels compared with those treated with nonfermented antler extract (Non-200 group, 5.20 ng/mL) and did not display statistically different serum EPO levels from nonanemic rats (control group, 2.70 ng/mL). PHZ-induced hemolytic anemia significantly increased renal EPO mRNA expression as well as serum EPO (PHZ-con as a percentage of control, 144.58%; P<.05). Increases in renal EPO mRNA expression in anemic rats were significantly suppressed by treatment with antler extract (P<.05). In addition, the renal EPO mRNA expression in rats treated with fermented antler extract (B-100 and B-200 groups, 99.21% and 106.06%, respectively) were lower than in rats treated with nonfermented antler extract (Non-200 group, 113.38%), although the differences were not significant.

Effect of fermented antler extract on
Hepatic ALAD activity
As shown in Figure 3, hepatic ALAD activity was decreased significantly in PHZ-induced anemic rats compared with nonanemic rats (control vs. PHZ-con, 62.54% vs. 44.06%; P<.05). Decreases in hepatic ALAD activities in anemic rats were improved significantly by treatment with antler extract (P<.05). However, significant differences in hepatic ALAD activities were not observed between fermented antler extract treatment and nonfermented antler extract treatment.

Effect of fermented antler extract on hepatic δ-aminolevulinic acid dehydratase (ALAD) activity in PHZ-induced anemic rats. Data are mean±SD values for eight rats per group. The percentage of ALAD activity is determined by the ratio (absorbance with sample – absorbance without sample)/absorbance with sample. abcMeans with different superscript letters are significantly different at P<.05 by Tukey's multiple comparison test. See Table 1 for descriptions of groups.
Osmotic resistance
Effects of treatments on osmotic hemolysis of RBCs due to hypotonic pressure are shown in Figure 4. The percentage of hemolysis was determined by the absorbance ratio of test/full hemolysis. A hemolysis curve was generated according to NaCl concentration. Hemolysis in hypotonic saline was the highest in the control group at all saline concentrations, and the percentage hemolysis of anemic rats was less than that of nonanemic rats. These results indicated that PHZ-induced hemolytic anemia tended to increase the osmotic resistance of RBCs. RBCs in PHZ-treated rats may be more susceptible to osmotic pressure than those in nonanemic rats. Furthermore, the treatment with antler extract, especially fermented antler extract, tended to enhance this increase in the osmotic resistance of RBCs in anemic rats. However, the changes of osmotic resistance did not differ significantly among all groups at all saline concentrations.
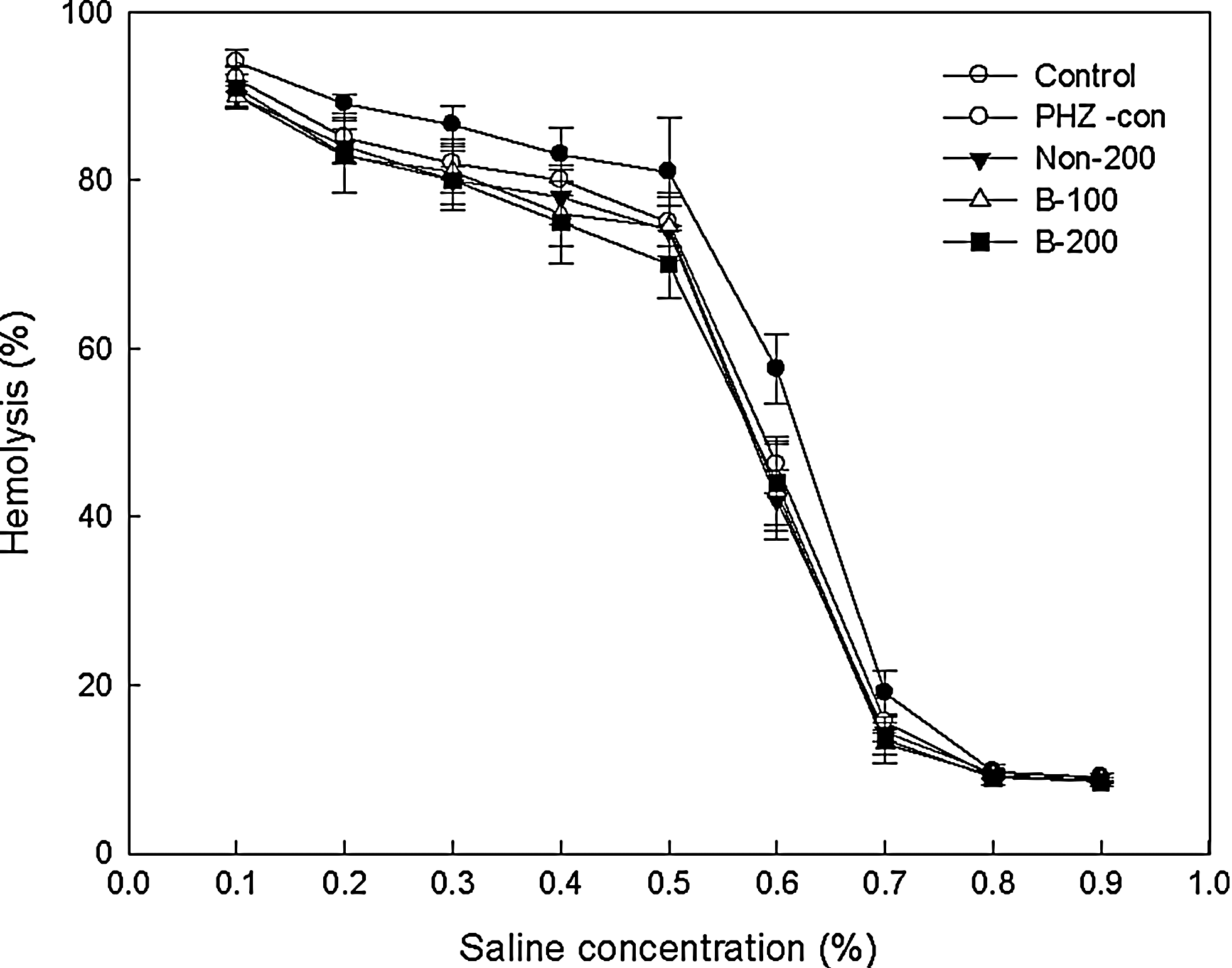
Effect of fermented antler extract on osmotic hemolysis of RBCs in response to hypotonic pressure in PHZ-induced anemic rats. Data are mean±SD values for eight rats per group. The percentage of hemolysis is determined by the ratio absorbance of test/absorbance of full hemolysis. See Table 1 for descriptions of groups.
Discussion
Antler accelerates recovery from hematopoietic injuries. 7,8 Many substances from antler have been identified and claimed to be active components, including free amino acids, polypeptides, trace elements, carbohydrates, hexosamines, mucopolysaccharides, uronic acids, sialic acids, prostaglandins, glycolipids, phospholipids, gangliosides, nucleic acids, hypoxanthine, cholesterol, and cholesterol esters. 6 –10,24 Recently, Yang et al. 7 reported that monoacetyldiglycerides purified from antler possess a significant clinical potential for the acceleration of hematopoiesis as the active hematopoiesis stimulating component.
In this study, the changes of some hematopoietic factors, such as EPO and hepatic ALAD activity, induced by PHZ exposure were restored to within the normal range when anemic rats were treated with antler extract. These results on the hematopoietic action of antler extract agree with previous studies. Yong 13 observed that hemolytic anemic rabbits receiving antler extract had higher RBC, Hct, and Hgb levels and lower reticulocyte levels compared with anemic rabbits treated with vehicle. Kim and Park 12 also reported that antler extract accelerated the recovery of RBCs and hepatic ALAD activity (maximum of 36.8%) of anemic rats. They concluded that these activities partly support the stimulating effect of antler on hematopoietic activity. In the study of Lee et al., 15 RBC, Hct, and Hgb levels were also increased significantly in anemic rats treated with antler extract compared with nontreated anemic rats. Moreover, they found that the administration of antler extract significantly improved serum EPO and hepatic ALAD. Therefore, we can conclude that antler improves hemolytic anemia by improving hematopoietic factors, including hematological values, EPO levels, and hepatic ALAD activity.
Food fermentation has been used for biological enrichment of food. Fermentation increases absorption rate and has merits for standardizing efficacy. 12,13 The sialic acid content and protein content of antler extract increase during fermentation. 19 Sialic acid is a component of gangliosides, and the biological activity of antler is attributed to the gangliosides. Gangliosides, which are concentrated in the plasma membrane of cells and modulate transmembrane signaling, are expressed as a surface marker of the cells in hematopoietic organs committed to a specific cell lineage. 25 We suggest that antler cell membranes were hydrolyzed by fermentation; thus, active compounds, such as proteins and gangliosides, were released. We surmise that the activity increases were related to increased silalic acid content and ganglioside production. In our previous study, we demonstrated that fermented antler can enhance the proliferation of osteoblasts and bone matrix proteins, and the stimulatory effects on cell proliferation and alkaline phosphatase production of fermented antler were greater than those of nonfermented antler. 19 In our another study, the anticomplementary activity of fermented antler extract was higher than that of nonfermented antler extract, and the ability of fermented antler extract to scavenge 2,2-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) and 1,1-diphenyl-2-picrylhydrazyl radicals was also significantly greater than that of nonfermented antler extract. 18 In this study, antler extract improved the hemolytic anemia, and these effects were slightly more pronounced in the treatment with fermented antler extract compared with nonfermented antler extract. In particular, the RBC level in the fermented antler group was significantly higher than that in the nonfermented antler group, and fermented antler extract induced the Hgb and serum EPO levels within the ranges of those of nonanemic rats, whereas nonfermented antler extract did not. Therefore, we conclude that the hematopoietic activity of antler as well as bone growth, immunomodulating, and antioxidant activities might be increased through the fermentation process.
In conclusion, fermented antler appears to have the potential to improve hemolytic anemia and confers beneficial properties as a useful functional food. Although the experimental studies demonstrated the pharmacological effects of fermented antler, more studies are needed to support its role in medical care.
Footnotes
Acknowledgment
This research was supported by the Technology Development Program for Food, Ministry for Food, Agriculture, Forestry and Fisheries, Republic of Korea.
Author Disclosure Statement
S.H.L. is an employee of Kwang Dong Pharmaceutical Co., Ltd. H.J.S., H.-S.L., S.P., J.-W.P., and E.Y.J. declare no competing financial interests exist.