
Abstract
We identified four anti-inflammatory sulfur-containing compounds from garlic, and their chemical structures were identified as Z- and E-ajoene and oxidized sulfonyl derivatives of ajoene. The sulfur compounds inhibited the production of nitric oxide (NO) and prostaglandin E2 (PGE2) and the expression of the pro-inflammatory cytokines tumor necrosis factor-α, interleukin-1β, and interleukin-6 in lipopolysaccharide (LPS)-activated macrophages. Western blotting and reverse transcription–polymerase chain reaction analysis demonstrated that these sulfur compounds attenuated the LPS-induced expression of the inducible NO synthase (iNOS) and cyclooxygenase-2 (COX-2) proteins and mRNA. Moreover, these sulfur-containing compounds suppressed the nuclear factor-κB (NF-κB) transcriptional activity and the degradation of inhibitory-κBα in LPS-activated macrophages. Furthermore, we observed that they markedly inhibited the LPS-induced phosphorylations of p38 mitogen-activated protein kinases and extracellular signal-regulated kinases (ERK) at 20 μM. These data demonstrate that the sulfur compounds from garlic, (Z, E)-ajoene and their sulfonyl analogs, can suppress the LPS-induced production of NO/PGE2 and the expression of iNOS/COX-2 genes by inhibiting the NF-κB activation and the phosphorylations of p38 and ERK. Taken together, these data show that Z- and E-ajoene and their sulfonyl analogs from garlic might have anti-inflammatory therapeutic potential.
Introduction
G
Garlic contains diverse organosulfur compounds such as S-allyl-
We previously showed that garlic extract exerts anti-inflammatory activity by inhibiting the LPS-induced Toll-like receptor-4 dimerization followed by the suppression of NF-κB transcriptional activity and the expression of iNOS and cyclooxygenase (COX)-2.
23
NO synthase (NOS) catalyzes the oxidative deamination of
In this study, we identified (Z, E)-ajoenes and their oxidized analogs as anti-inflammatory principles of garlic (A. sativum L.) through the activity-guided purification procedure. Although the effect of the mixture of Z- and E-ajoene on the production of inflammatory mediators in LPS-induced macrophages has been investigated, 17,22 little is known about the effect of pure Z- and E-ajoene and oxidized (sulfonyl) derivatives of ajoene on inflammatory responses. The present study compared the efficacy of Z- and E-ajoene and their analogs for inhibiting production of NO/PGE2 and the expression of iNOS/COX-2 in LPS-stimulated macrophages. We also disclosed the possible molecular mechanisms of these actions.
Materials and Methods
Test material
Fresh Korean garlic (A. sativum L.) (2 kg) was purchased from a Korean market in January 2007. Authentication of plant material was carried out by Prof. K.S. Yang at Sookmyung Women's University (Seoul, Korea). A voucher specimen (number 0070108) was deposited in the Herbarium of Sookmyung Women's University. Four sulfur-containing compounds,
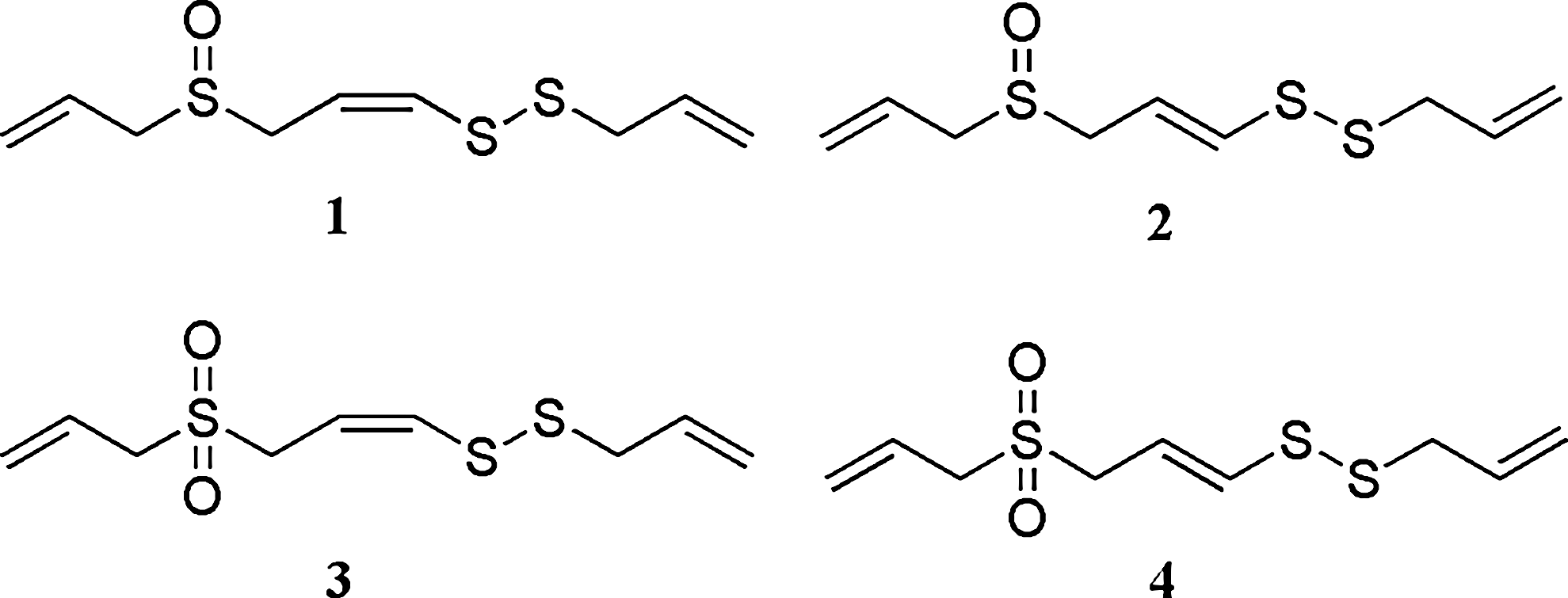
The structures of sulfur compounds
Cell culture
RAW 264.7 cells (a murine macrophage cell line) (American Type Culture Collection, Rockville, MD, USA) were cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum, 100 units/mL penicillin, and 100 μg/mL streptomycin (Invitrogen, Carlsbad, CA, USA). T-RAW cells, stably transfected RAW 264.7 cells with a reporter construct of pNF-κB-SEAP-NPT encoding four copies of NF-κB binding κB sequence and secretory alkaline phosphatase (SEAP) as a reporter, were the kind gift of Prof. Yeong Shik Kim (Seoul National University, Seoul). T-RAW 264.7 cells were grown in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 500 μg/mL geneticin. All cells were incubated at 37°C in 5% CO2 in a humidified atmosphere.
Measurements of NO in LPS-activated RAW 264.7 cells
Cells (6×104 cells/mL in a 48-well plate) were incubated for 20 h in the absence or presence of test samples with LPS (1 μg/mL). NO was assessed by measuring the accumulated nitrite by the Griess method. 30 In brief, samples (100 μL) of culture medium were incubated with 150 μL of Griess reagent (1% sulfanilamide and 0.1% naphthylethylenediamine in 2.5% phosphoric acid solution) at room temperature for 10 min in a 96-well microplate. Absorbance at 540 nm was measured by using a microplate reader (Molecular Devices, Sunnyvale, CA, USA). The concentration of NO was determined by the sodium nitrite standard curve.
Measurement of PGE2
The accumulated PGE2 in culture medium was determined by using an enzyme immunoassay kit from Cayman Chemical Co. (Ann Arbor, MI, USA) according to the manufacturer's instruction. A standard curve was prepared simultaneously with PGE2 standards ranging from 0.06 to 6 ng/mL.
Western blot analysis
Whole-cell lysates and cytosol and nuclear extracts were prepared and subjected to western blotting as described previously. 31 In brief, RAW 264.7 cells (8×105 cells per 60-mm-diameter dish) were treated with 1 μg/mL LPS in the presence or absence of test compounds. Following 20-h treatment, cells were harvested and gently lysed with cell lysis buffer (Cell Signaling Technologies, Beverly, MA, USA). Cell lysates were then centrifuged at 10,000 g for 20 min at 4°C. Supernatants were collected, and protein concentrations were determined by the Bradford method.
To prepare cytosol and nuclear extracts, cells were treated with test compounds for 30 min prior to activation with 1 μg/mL LPS. Following a 15-min treatment with LPS, cells were harvested by using NE-PER nuclear and cytoplasmic extraction reagents (Pierce Biotechnology, Rockford, IL, USA) according to the manufacturer's instructions. Antibodies against iNOS (BD Biosciences, Franklin Lakes, NJ, USA), COX-2 (Cayman Chemical), and inhibitory-κBα (I-κBα) and p65 (Santa Cruz Biotechnologies Inc., Santa Cruz, CA, USA) were used for immunoblot analysis.
Reverse transcription–polymerase chain reaction analysis
RAW 264.7 cells (1×106 cells per 60-mm-diameter dish) were stimulated for 6 h with LPS (1 μg/mL) in the presence or absence of test compounds. Total RNA was isolated by TRIzol® (Invitrogen) extraction according to the manufacturer's instructions and then reverse-transcribed into cDNA using reverse transcriptase (Invitrogen) and random hexamer (Cosmo, Seoul, Korea). Then polymerase chain reaction analyses were performed on the aliquots of the cDNA preparations to detect expression of the genes for iNOS, COX-2, IL-1β, IL-6, TNF-α, and β-actin using a recombinant Taq polymerase (Promega, Madison, WI, USA).
Measurement of NF-κB transcriptional activity
NF-κB transcriptional activity was measured by using the stably transfected RAW 264.7 cells with pNF-κB-SEAP-NPT (T-RAW 264.7 cells) as described previously with some modifications. 32,33 In brief, T -RAW 264.7 cells were seeded on a 24-well plate and incubated for 24 h. Test compounds were added to cells 2 h before the treatment with LPS (1 μg/mL). After a 16-h incubation, aliquots of culture medium were heated at 65°C for 6 min, and then the activity of SEAP was measured. The transcriptional activity was expressed as fold induction over that of vehicle-treated cells.
Statistical analysis
The results were expressed as mean±SD values of three experiments, and statistical analysis was performed by one-way analysis of variance and Student's t test. A P value of < .01 was considered to indicate a significant difference.
Results
Effects of sulfur compounds on production of NO and PGE2 in LPS-stimulated RAW 264.7 cells
Compounds
RAW 264.7 cells were cultured in 48-well plates and activated with 1 μg/mL lipopolysaccharide in the presence or absence of various concentrations (0.1–20 μM) of compounds
The accumulated prostaglandin E2 (PGE2) in culture medium was determined by using an enzyme immunoassay kit from Cayman Chemical according to the manufacturer's instruction. A standard curve was prepared simultaneously with PGE2 standard ranging from 0.06 to 6 ng/mL.
IC50, concentration causing 50% inhibition; NO, nitric oxide.
Effects of sulfur compounds on expression of iNOS and COX-2 in LPS-stimulated RAW 264.7 cells
To elucidate the mechanism of active compounds for the inhibition of NO and PGE2 production, we examined the effects of sulfur compounds on the expression of iNOS/COX-2 protein and mRNA. As shown in Figure 2A, sulfur compounds
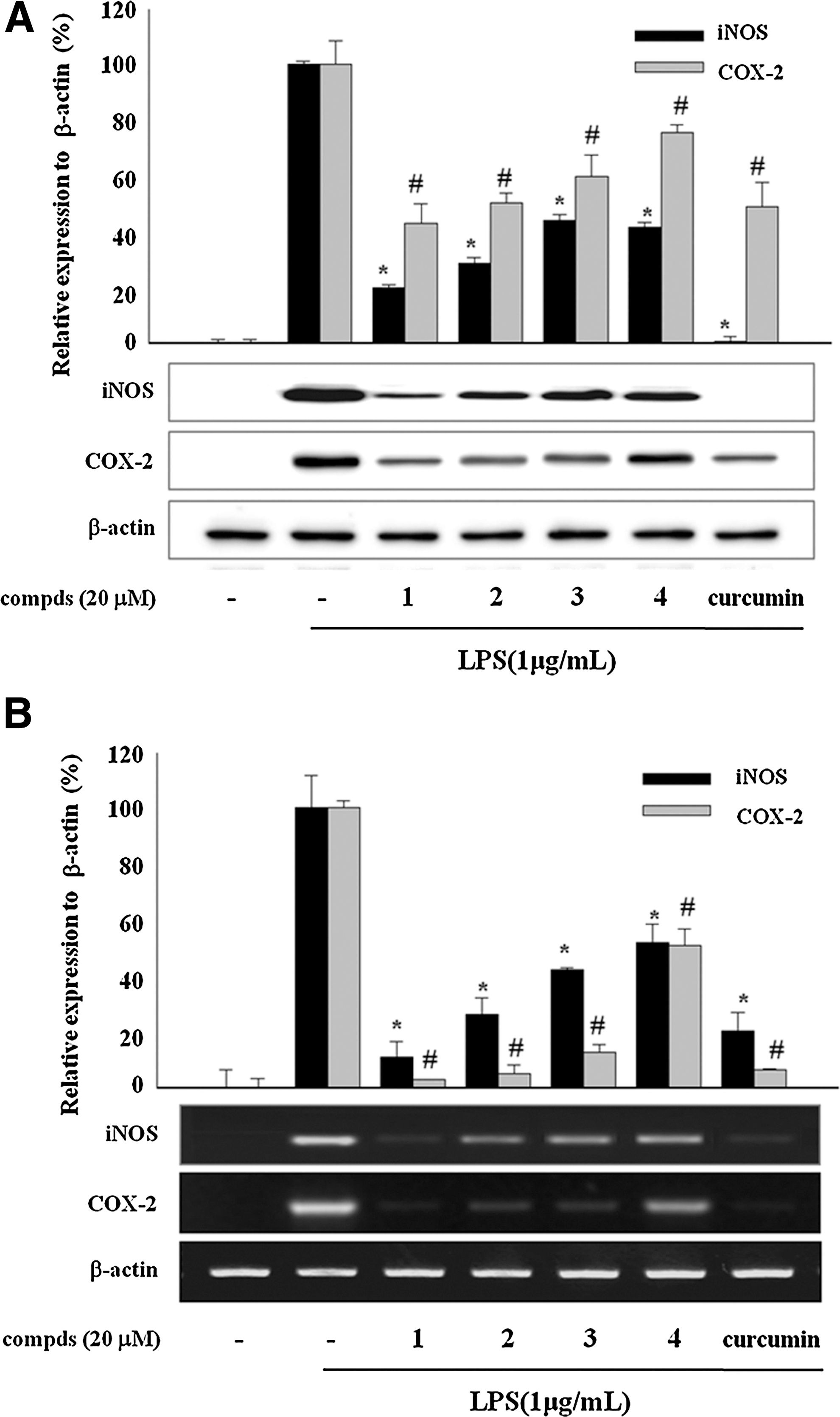
Effects of sulfur compounds (compds)
Effects of sulfur compounds on mRNA expression of pro-inflammatory cytokines in LPS-stimulated RAW 264.7 cells
To examine the anti-inflammatory potential of these sulfur compounds from garlic, we investigated the effect of the compounds
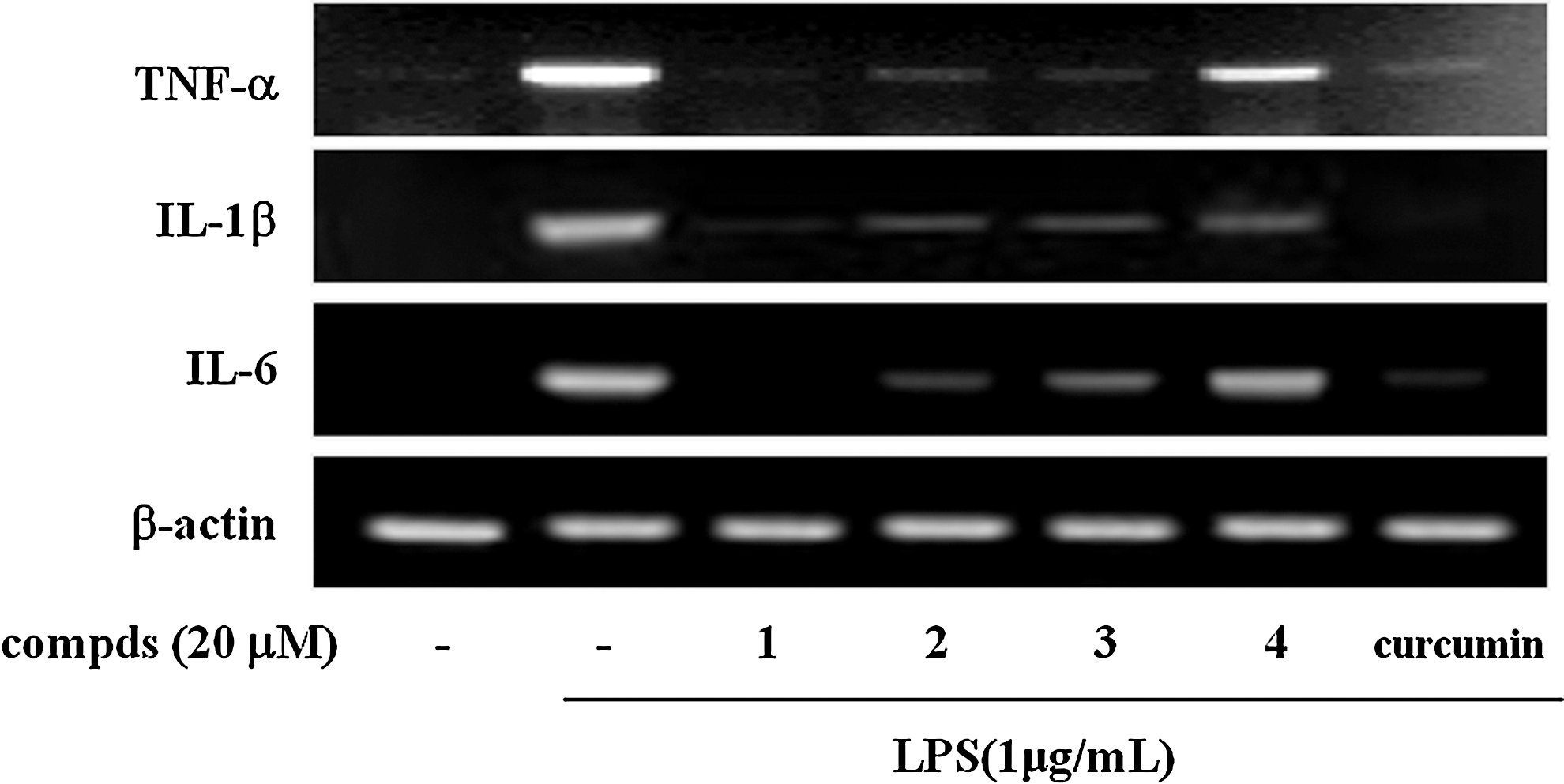
Effects of sulfur compds
Inhibition of NF-κB transcriptional activity by sulfur compounds via the suppression of I-κBα degradation and nuclear translocation of p65 in LPS-stimulated macrophages
To reveal the molecular mechanism of the suppression of LPS-induced pro-inflammatory enzyme and cytokine expressions by the sulfur compounds
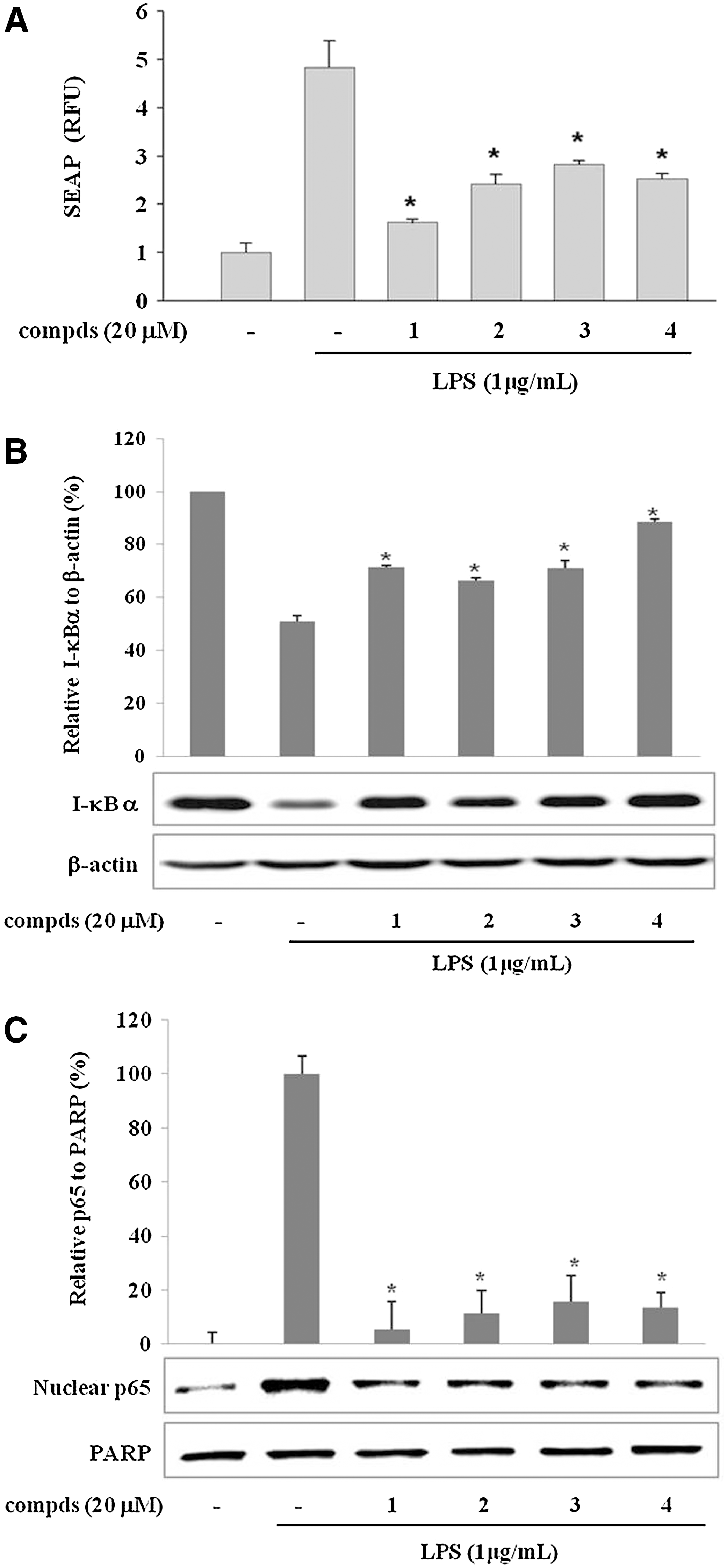
Sulfur compounds
Next, we checked whether the active sulfur compounds affect the LPS-induced I-κBα degradation. NF-κB, composed of p50 and p65 subunits, is located in the cytoplasm as an inactive p50/p65 dimer that is physically combined with I-κB.
35
In response to pro-inflammatory stimuli, I-κB is phosphorylated, ubiquitinated, and rapidly degraded to release and activate p50/p65. Active NF-κB (p50/p65 dimer) translocates to the nucleus and induces the expression of pro-inflammatory genes.
35
The I-κBα was fully degraded by a 15-min incubation with LPS (1 μg/mL) and regenerated gradually afterwards (data not shown). As shown in Figure 4B, the LPS-induced degradation of I-κBα was suppressed by the treatment of 20 μM compounds
Suppression of phosphorylation of mitogen-activated protein kinases by sulfur compounds in LPS-stimulated RAW 264.7 cells
The mitogen-activated protein kinase (MAPKs) pathways are implicated in the up-regulation of LPS-induced expression of pro-inflammatory mediators in macrophages.
36,37
To investigate whether the inhibition of compounds
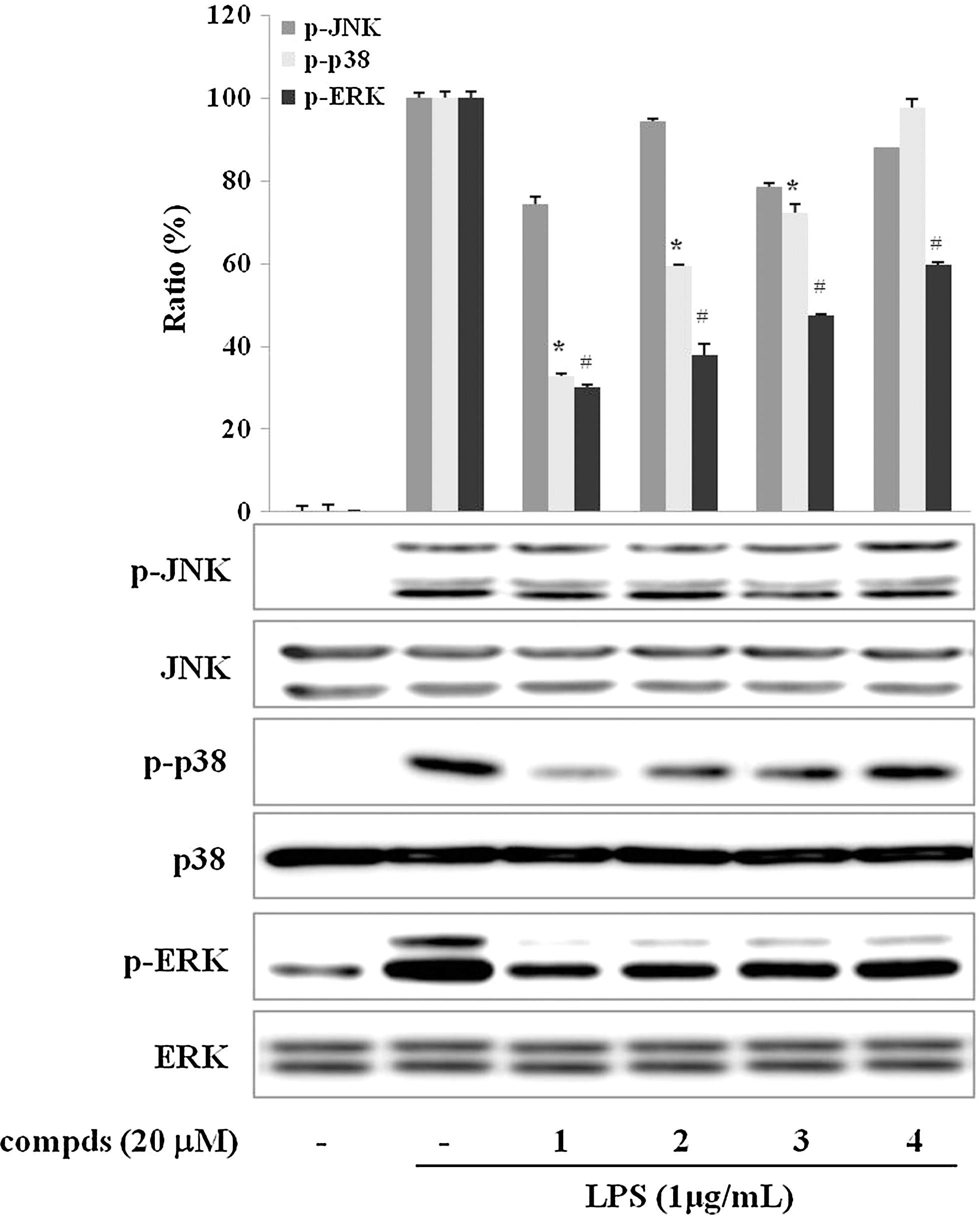
Effect of sulfur compds
Discussion
Garlic (A. sativum L.) is a common spice for cooking and also a popular herbal remedy for the treatment of wide variety of health problems, including infection, cancer, and cardiovascular and inflammatory diseases. Garlic is rich in sulfur-containing compounds, which are responsible for the most of its biological activities. 38
Our previous study showed that garlic extract exerted anti-inflammatory activity by inhibiting Toll-like receptor-mediated signaling pathways at the receptor level.
23
In the present study, we isolated four sulfur compounds,
During inflammatory responses, the activation of macrophages contributes to host damage by excessive release of various inflammatory mediators such as NO and PGE2 and of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α. Many studies have demonstrated that the overexpressions of these inflammatory mediators and cytokines are responsible for many chronic inflammatory diseases such as diabetes, atherosclerosis, and rheumatoid arthritis.
34,39
–41
We demonstrated the inhibitory effect of sulfur compounds
Yoshida et al. 42 demonstrated that Z-ajoene exhibits an antimicrobial activity at least twofold higher than that of the E isomer. The Z-ajoene isomer is also reported to be more potent than its E isomer for antithrombotic activity and the inhibition of cancer cell proliferation. 28,43 –45 The study of Block et al. 28 was undertaken to compare the antithrombotic activity of two ajoene isomers and its homologs. It has been suggested that Z-ajoene and its homologs are more active than E isomers, and these activities are retained upon the oxidation of the sulfonyl derivatives of ajoene but are lost upon reduction of the sulfinyl group to a sulfide group. These trends are also observed in the present study, suggesting that Z-ajoene exhibits more potent anti-inflammatory activity than the E isomer and that the oxidized derivatives of ajoene also possess significant anti-inflammatory activity.
NF-κB is a master switch for the regulation of pro-inflammatory genes, including IL-1β, IL-6, and TNF-α, as well as the hallmarks of inflammation, NO and PGE2.
34
NF-κB is present in cytoplasm as a heterodimer consisting of p50 and p65, which are bound by I-κBα in resting macrophages. Upon activation by proper stimulation, phosphorylation and degradation of I-κBα release the p50/p65 complex. The released NF-κB translocates to the nucleus and regulates the transcription of target genes through the binding to specific sequences in the DNA.
35
We observed that sulfur compounds
The MAPKs signaling pathways are involved in LPS-induced iNOS and COX-2 expression in activated macrophages. Moreover, it has been demonstrated that MAPKs play critical roles in the activation of NF-κB. 46 However, they seems be differently involved in the response of anti-inflammatory compounds in macrophages. 47,48 In the present study, the four sulfur compounds were found to inhibit the phosphorylation of p38 and ERK, but not that of JNK, in LPS-activated RAW 264.7 macrophages (Fig. 5).
When the data are taken as a whole, the four sulfur compounds
Footnotes
Acknowledgments
This work was supported by Sookmyung Women's University Research Grants in 2010 and by the MRC program (2011-0030699) through the National Research Foundation of Korea funded by the Korean Government.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.