
Abstract
β-Caryophyllene (trans-4,11,11-trimethyl-8-methylenebicyclo[7,2,0]undec-4-ene), found in various plants, is a natural bicyclic sesquiterpene with a low toxicity. Here, we show that a single intraperitoneal injection of β-caryophyllene (10 mg/kg) significantly reduced the cortical infarct volume by 67% when given immediately before middle cerebral artery occlusion (MCAO). Neurological deficits caused by MCAO were also significantly decreased by β-caryophyllene. β-Caryophyllene treatment of cortical cells exposed to oxygen–glucose deprivation revealed a significant protection in a dose-dependent manner. However, β-caryophyllene neither suppressed N-methyl-
Introduction
I
Cerebral ischemia triggers a complex series of biochemical and molecular mechanisms that impair the neurologic function through breakdown of cellular integrity mediated by excitotoxic signaling, ionic imbalance, reactive oxygen/nitrogen species (ROS/RNS) reactions, and inflammation. 4 Recent studies have indicated that brain ischemia, especially a clinically common focal stroke caused by occlusion of the middle cerebral artery (MCA), involves a secondary inflammation that significantly contributes to the outcome after an ischemic insult. 5,6 Major players in inflammatory injury are cytokines (interleukin [IL]-1, IL-6, and tumor necrosis factor [TNF]-α), eicosanoids and inducible nitric oxide synthase (iNOS), which are produced by endothelial cells, astrocytes, microglial cells, and leukocytes. 7
β-Caryophyllene (trans-4,11,11-trimethyl-8-methylenebicyclo[7,2,0]undec-4-ene) is a natural bicyclic sesquiterpene (Fig. 1), which is isolated from clove leaf oil, clove stem oil, cinnamon leaf oil, and pine oil fractions. It has been used as a flavoring agent or an ingredient of fragrances since the 1930s. 8 Further, it has been granted the generally-recognized-as-safe (GRAS) status by the Flavor and Extract Manufacturers Association 9 and has approval by the U.S. FDA for use in food 8 because of its low toxicity. This compound is known to be antimicrobial, 10 antioxidant 10,11 and anticarcinogenic, 12 and to possess skin penetration-enhancing properties. 13 In particular, essential oils that have β-caryophyllene as a major component (30.6%) showed a marked anti-inflammatory activity against carrageenan- and prostaglandin E (PGE)1-induced edema in rats as well as antiarthritic activity. 14,15 Recently, we demonstrated that β-caryophyllene exerts prophylactic or curative effects against inflammatory bowel disease through its antioxidative and/or anti-inflammatory action. 16 Thus, we hypothesized that β-caryophyllene may protect against cerebral ischemic insult possibly through its anti-inflammatory action. However, the influence of β-caryophyllene on cerebral ischemic injury has not yet been explored.
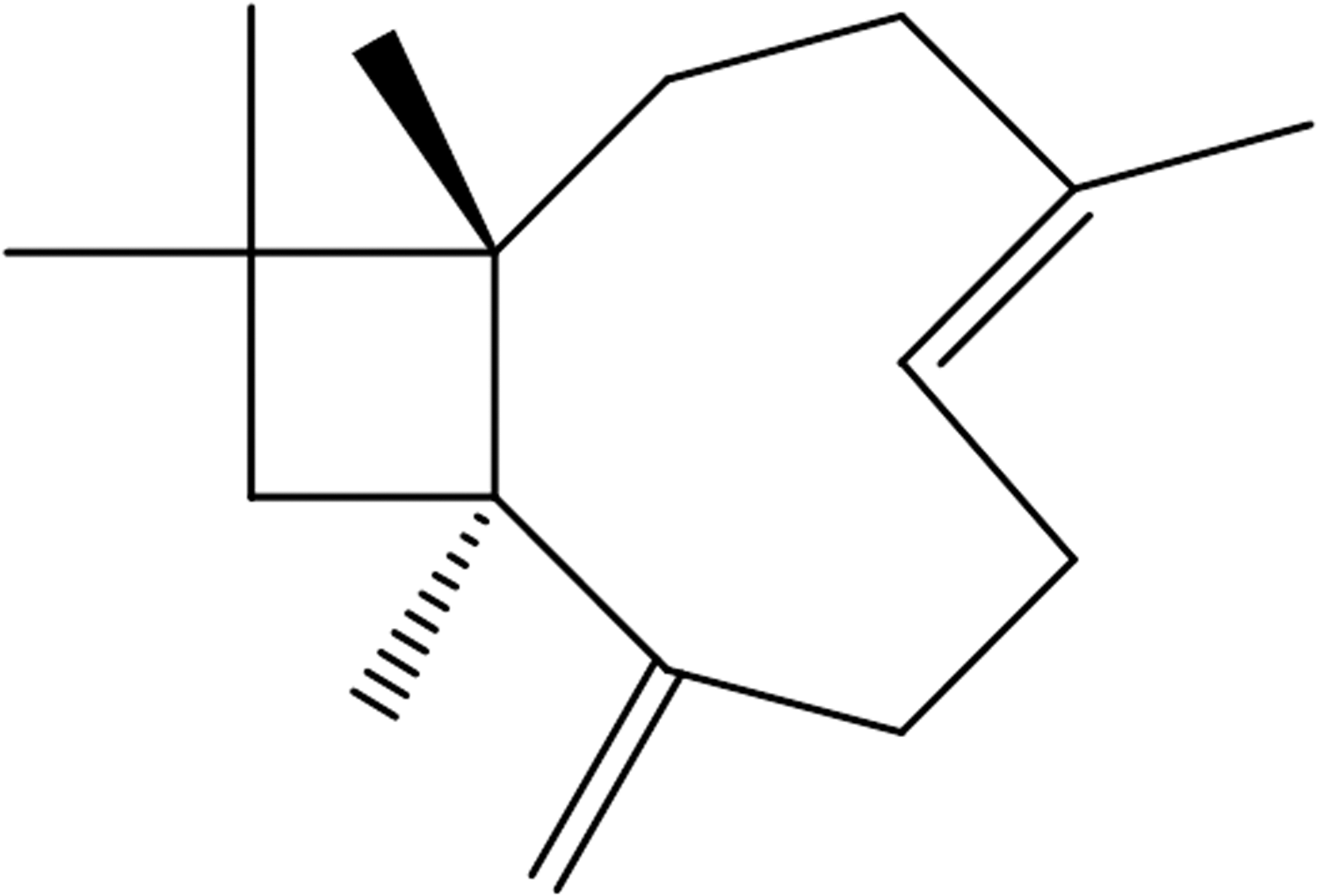
Structure of β-caryophyllene.
In the present study, we investigated the protective effect of β-caryophyllene on brain tissue after ischemic insults both in vitro and in vivo. We further examined whether the neuroprotective effect of β-caryophyllene was associated with the modulation of excitotoxicity, ROS generation, and/or inflammatory reaction using in vitro models.
Materials and Methods
Materials
β-Caryophyllene was provided by MP Biomedicals (Solon, OH, USA). Dulbecco's modified eagle's medium (DMEM), Ham's F-12, glucose-free DMEM, trypsin/EDTA, penicillin/streptomycin, and fetal bovine serum were purchased from Gibco BRL (Rockville, MD, USA). TNF-α, IL-1β, and IL-6 ELISA kits were purchased from Biosource International, Inc. (Camarillo, CA, USA). 2,2′-Azinobis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS), deoxyribose, 2,7-dihydrodichlorofluorescein diacetate (DCFH2-DA), dihydrorhodamine-123 (DHR-123), dimethyl sulfoxide (DMSO), 3-[4,5-dimethylthiazole-2-yl]-2,5-diphenyltetrazolium bromide (MTT), guaiacol, horseradish peroxidase, 6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox), lipopolysaccharide (LPS), N-methyl-
MCA occlusion
All surgical procedures were performed in accordance with the institution's guidelines. Male Sprague-Dawley rats weighing 270–290 g were initially anesthetized with 3.0% isoflurane in a 70% N2O and 30% O2 (v/v) mixture via a facemask. Anesthesia was maintained with 2.0% isoflurane. A rectal temperature probe was introduced, and a heating pad maintained the body temperature at 37°C during the entire surgical period. Focal cerebral ischemia was achieved by right-sided endovascular middle cerebral artery occlusion (MCAO). 17,18
Measurement of infarct volume and neurological deficit scoring
Rat brains were cut into 2-mm-thick coronal slices of using a rat brain matrix (Ted Pella, Redding, CA, USA) and incubated in 2% TTC at 37°C for 30 min to reveal the ischemic infarction. After the TTC reaction, the brain slices were fixed with 4% paraformaldehyde (pH 7.4) in 0.1 M phosphate-buffered saline (PBS) for 1 day, and subsequently cryoprotected in PBS containing 30% sucrose at 48°C for 2 days. The cross-sectional area of infarction with the bregma levels between +4 mm (anterior) and −6 mm (posterior) were determined with a computer-assisted image analysis (BioScan, Inc., Edmonds, WA, USA). On each slice, the brain infarct size was measured manually by outlining the margins of the infarct areas, and the infarct volume was calculated according to the slice thickness. Each side of the brain slices was measured separately, and the mean values were calculated. The total volume of infarction was determined by integrating six chosen sections and expressed as the percentage of the total brain volume. Because postischemic brain edema increases the brain volume in the infarcted area, the corrected infarct volumes were calculated to compensate for brain edema.
Twenty-four hours after reperfusion after MCAO for 2 h, neurological deficits were measured according to the method of Huang et al. 19 The criteria for scoring were as follows: 0 (no observable neurological deficit, normal); 1 (failure to extend left forepaw on lifting the whole body by tail, mild); 2 (circling to the contralateral side, moderate); and 3 (leaning to the contralateral side at rest or no spontaneous motor activity, severe).
Cerebral cortical cultures and oxygen–glucose deprivation
Primary cultures of cortical cells were performed as described previously.
18
The cerebral cortices were derived from embryonic (fetal day 15–17) Sprague-Dawley rats. The cerebral cortical cells from the individual brains were dissociated mechanically, and plated in 24-well plates precoated with poly-
Oxygen–glucose deprivation (OGD) was performed by placing the glucose-deprived cells inside an anaerobic chamber (ThermoForma, Marietta, OH, USA) maintaining a 90% N2/5% CO2/5% H2 environment. 20 One hour after starting the OGD, the cells were returned to the normoglycemic (5.5 mM glucose) and normoxic condition for 6 h before the MTT assay. Cortical cells in the culture were exposed to OGD/reoxygenation in the absence or presence of MK-801 (10 μM) and/or β-caryophyllene (0.1–100 μM).
NMDA-induced neurotoxicity
After 14 days of culture, the cerebral cortical cells were incubated in serum-free DMEM containing 100 μM NMDA for 10 min at room temperature. The medium was then replaced with a fresh medium without NMDA, and the cells were cultured at a humidified 5% CO2/95% air atmosphere for a further 24 h. Cells were treated with MK-801 (10 μM) or β-caryophyllene (10–100 μM) during and after the NMDA treatment. Cell death was assessed by lactate dehydrogenase (LDH) activity released into the culture medium. LDH activity was measured using a diagnostic kit (Sigma Chemical Co., St Louis, MO, USA). Briefly, 20 μL of NADH in pyruvate solution (1 mg/mL) was added to 20 μL of the culture medium, and then it was incubated for 30 min at 37°C. After incubation, 20 μL of the color reagent (2, 4-dinitrophenylhydrazine in 1 N HCl) was added, and then it was incubated for 20 min at room temperature. After 100 μL of 0.4 M NaOH was added, it was measured the absorbance at 450 nm using a microtiter plate reader (Molecular Devices, Sunnyvale, CA, USA).
Measurement of DCF fluorescence
Cerebral cortical cells were loaded with DCFH2-DA (100 μM) in PBS for 10 min and then rinsed with the same solution. Cells were deprived of glucose in the absence and presence of SIN-1 (200 μM) for 1 h. Cells were then exposed to ascorbic acid (100 μM) or β-caryophyllene (10–100 μM) for 1 h before and after DCF treatment. The fluorescence of DCF was measured at an excitation wavelength of 485 nm and an emission wavelength of 530 nm using a fluorescence microplate reader (FL600; Bio-Tek Instruments, Inc., Winooski, VT, USA).
BV-2 microglial cell culture
BV-2 microglial cells (generously provided by Dr. Tong H. Joh at Burke Institute, Cornell University) were cultured in DMEM supplemented with 10% fetal bovine serum and streptomycin (100 μg/mL)/penicillin (100 IU/mL). The cells were subcultured every 2 days, and maintained at 37°C in a humidified 5% CO2/95% air atmosphere. For the measurement of nitrite and cytokine production, the cells were plated into 96-well plates at a density of 5×104 cells/well. The cells were treated with β-caryophyllene (1, 10, and 100 μM) during stimulation with LPS (100 ng/mL) for 48 h. For reverse transcription–polymerase chain reaction (RT-PCR), the cells were plated into 100-mm culture dish at a density of 1×106 cell/mL. Cells were treated with β-caryophyllene (10 and 100 μM) for 1 h before and during stimulation with LPS (100 ng/mL) for 24 h.
Isolation of total RNA and RT-PCR
Total RNA from BV-2 microglial cells was isolated using TRIzol® reagent (Invitrogen Corp., Carlsbad, CA, USA), and cDNA was prepared using oligo-dT primer and M-MLV reverse transcriptase. PCR was performed using a PCR kit (Bioneer Corp., Daejeon, Korea) in Tetrad® Peltire Thermal Cyclers (MJ Research, Inc., Waltham, MA, USA) according to the manufacturer's instructions. The sequences of the sense and antisense primers were as follows: 5′-CCT TTC CGA AGT TTC TGG CAG CAG C-3′ and 5′-GGC TGT CAG AGC CTC GTG GCT TTG G-3′ for iNOS (497 bp); 5′-CCT GTA GCC CAC GTC GTA GC-3′ and 5′-TTG ACC TCA GCG CTG AGT TG-3′ for TNF (375 bp); 5′-TGC AGA GTT CCC CAA CTG GTA CAT C-3′ and 5′-GTG CTG CCT AAT GTC CCC TTG AAT C-3′ for IL-1β (388 bp); 5′-ATG AAG TTC CTC TCT GCA AGA GAC T-3′ and 5′-CAC TAG GTT TGC CGA GTA GAT CTC-3′ for IL-6 (638 bp); 5′-TGC ATG TGG CTG TGG ATG TCA TCA A-3′ and 5′-CAC TAA GAC AGA CCC GTC ATC TCC A-3′ for cyclooxygenase (COX)-1 (449 bp); 5′-GAA CAT TGT GAA CAA CAT CCC C-3′ and 5′-GGT GGC ATA CAT CAT CAG ACC-3′ for COX-2 (613 bp); and 5′-TGA TGA CAT CAA GAA GGT GGT GAA-3′ and 5′-TCC TTG GAG GCC ATG TAG GCC AT-3′ for GAPDH (240 bp). The PCR preparations were amplified using a three-temperature PCR system consisting of denaturation at 94°C for 30 sec, annealing at 65°C (for iNOS), 60°C (for TNF-α, IL-1β, COX-1 and 2, and GAPDH), or 55°C (for IL-6) for 30 sec, and extension at 72°C for 45 sec. The number of cycles used for iNOS, TNF-α, IL-1β, IL-6, COX-1 and 2, and GAPDH analysis were 32, 25, 25, 30, 29, 29, and 20 cycles, respectively. The PCR products were separated by 1.5% agarose gel electrophoresis, and visualized by ethidium bromide staining under UV light.
Measurement of nitric oxide, proinflammatory cytokines (TNF-α, IL-1β, and IL-6), and PGE2
The culture medium of BV-2 microglial cells was removed and assayed for nitrite accumulation. Briefly, 50 μL of culture supernatant was reacted with an equal volume of Griess reagent (0.1% naphthylethylenediamine/2.5% H3PO4 and 1% sulfanilamide in 2.5% H3PO4) for 10 min at room temperature in the dark. The absorbance at 540 nm was determined using a microplate reader (Molecular Devices Corp.). NaNO2 was used as a standard to calculate the nitrite concentration.
The levels of TNF-α, IL-1β, IL-6, and PGE2 in the culture medium of BV-2 microglial cells were measured with commercial ELISA kits according to the manufacturer's instructions. The absorbance at 450 nm was determined using a microplate reader.
Total antioxidant capacity and ROS/RNS-scavenging activity in an acellular system
The total antioxidant capacity was determined by a Trolox-equivalent antioxidant capacity (TEAC) assay according to the method of Re et al. 21 with slight modifications. The antioxidant capacity was expressed as scavenging activity relative to the water-soluble vitamin E analog, Trolox.
Hydrogen peroxide (H2O2) scavenging was measured using the guaiacol method of Aruoma et al. 22 Scavenging of superoxide radical (O2 •−) was measured according to the method of Robak and Gryglewski. 23 The nonenzymatic generation of superoxide anions was detected by reduction of NBT in a PMS-NADH system. Scavenging of the hydroxyl radical (OH•) was determined using the deoxyribose method, in which deoxyribose exposed to OH• generated by the Fenton system (ascorbic acid/FeSO4-EDTA/H2O2) was degraded to malonaldehyde, which generates a pink chromogen on heating with TBA. 24 Scavenging of nitric oxide (NO) was examined by the Griess reaction, and SNP was used as the NO generator. 25 Scavenging of peroxynitrite (ONOO−), formed from SIN-1, was determined by measuring the ONOO−-induced oxidation of DHR-123 into rhodamine-123 by fluorimetry. 26 Scavenging of hypochlorous acid (HOCl) was determined by measuring the chlorination of taurine according to the method of Nève et al. 27 Ascorbic acid for O2 −, H2O2, HOCl, and ONOO− scavenging, mannitol for HO− scavenging, and quercetin for NO scavenging were used as positive controls. The result was expressed as the concentration of β-caryophyllene that is required for 50% inhibition (IC50, μM).
Statistical analysis
Data were expressed as the means±S.E.M, and statistical comparison between groups was evaluated by Student's t-test. A value of P<.05 was considered statistically significant.
Results
β-Caryophyllene alleviates ischemic injury after MCAO
To test whether β-caryophyllene decreases ischemic injury, 10 mg/kg of β-caryophyllene was intraperitoneally administered. Two hours of MCAO and 24 h of reperfusion resulted in remarkable infarction as shown in the TTC-stained brain sections (Fig. 2A). After a single administration, right before MCAO, β-caryophyllene reduced the size of the infarct area. Quantitative analysis of the infarction showed that the total infarct volume was significantly smaller from 26.7%±3.92% to 8.9%±2.87% after β-caryophyllene treatment (Fig. 2B, P<.05). There was a 67% decrease in the infarct volume. The effect of β-caryophyllene on the neurological score after MCAO is shown in Figure 2C. A significant reduction in the neurological score was apparent in β-caryophyllene-treated rats (1.2±0.27, P<.05), compared with the untreated rats (2.8±0.41).
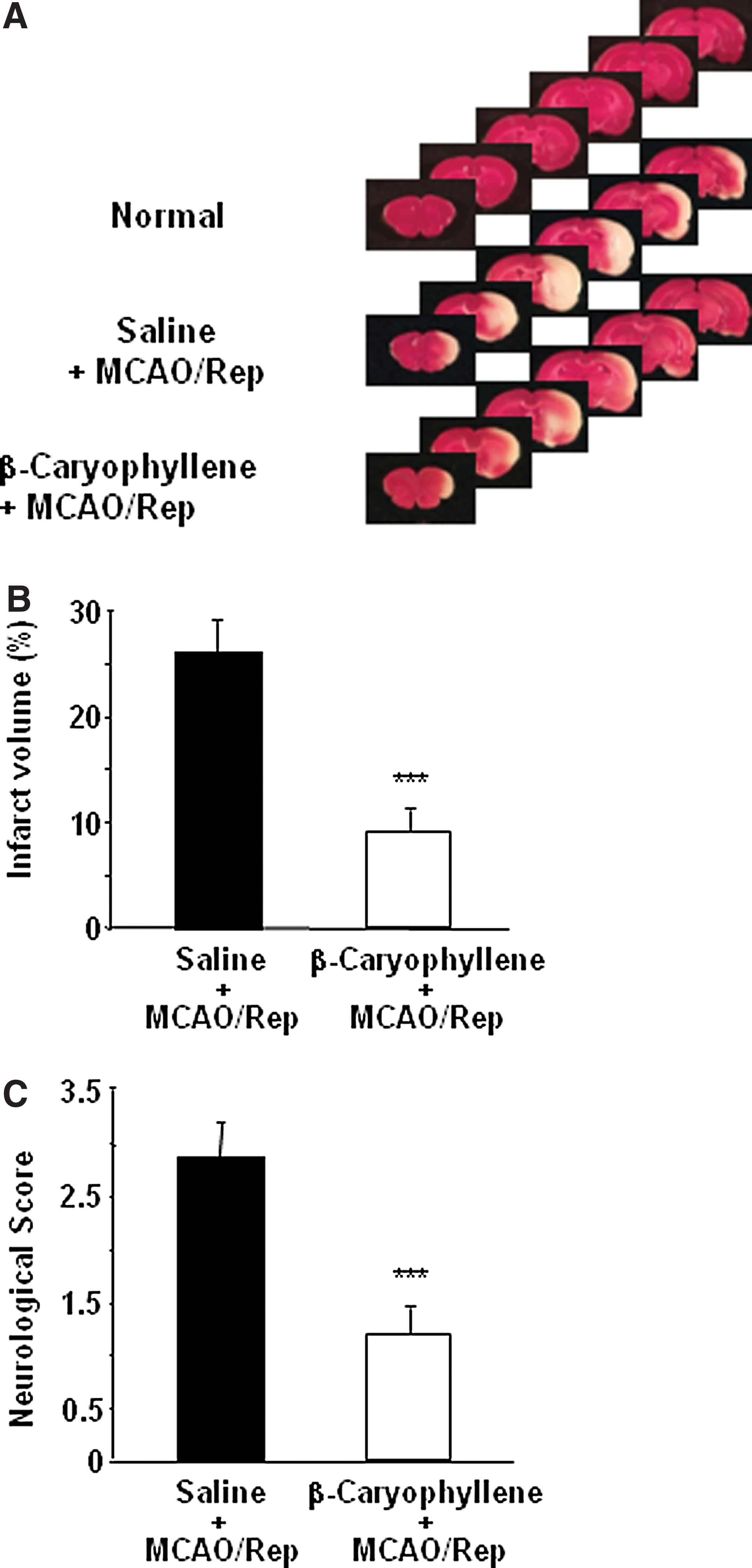
Neuroprotection by β-caryophyllene in the rat MCAO/reperfusion model.
Effect of β-caryophyllene on OGD-mediated and NMDA-induced cell death in cortical neurons
Rat cortical cells were exposed to a lethal OGD/reoxygenation, and the cell damage was assessed by MTT measurement in the supernatant medium. As shown in Figure 3A, 10 and 100 μM treatments of β-caryophyllene in cortical cells challenged with OGD provided significant and dose-dependent protection. To investigate the neuroprotective effect of β-caryophyllene against excitotoxicity, NMDA-induced cell death was assessed by measuring LDH activity, expressed as a percentage of the control activity in the culture medium of rat cortical cells. As presented in Figure 3B, MK 801, an NMDA receptor antagonist, significantly decreased LDH release compared with that of the control level, but β-caryophyllene did not show any decrease at the concentrations used.
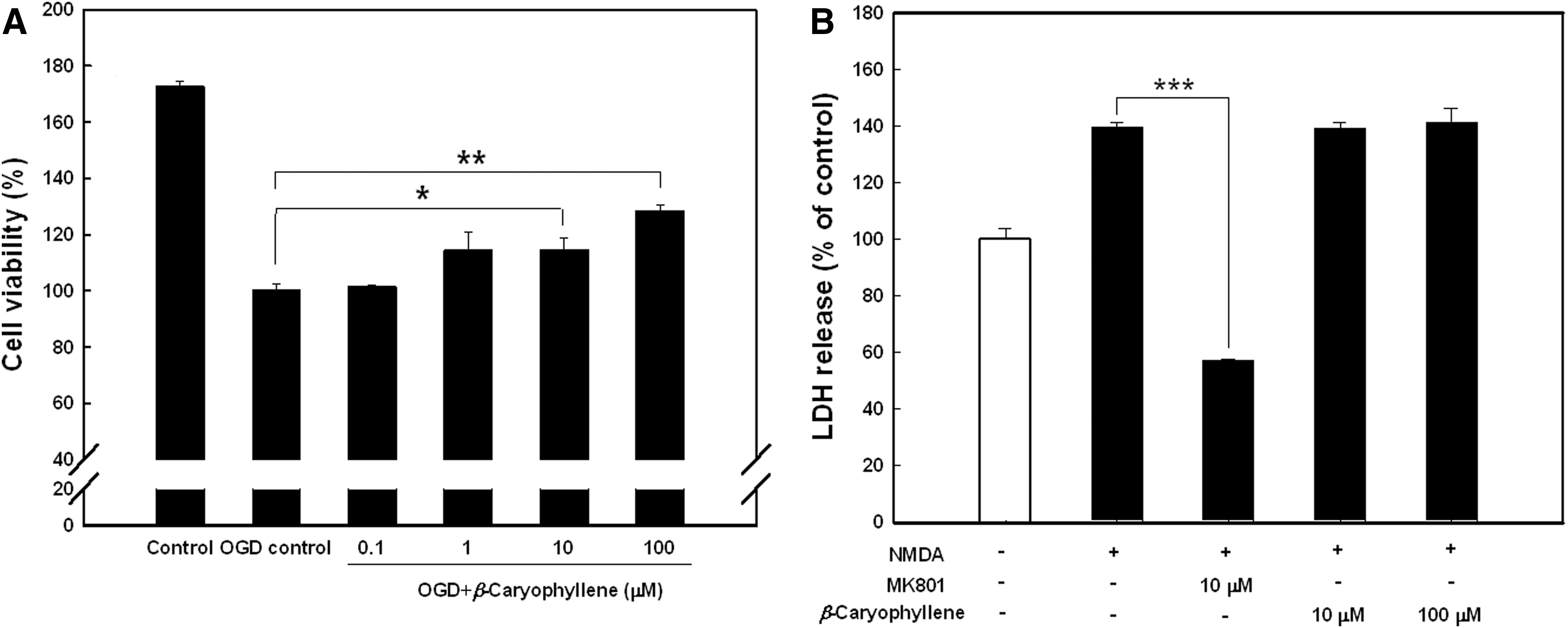
Effect of β-caryophyllene against OGD- and NMDA-induced toxicity in rat primary cortical cells.
Effect on the intracellular levels of ROS, TEAC, and free radical scavenging in the cellular and acellular systems
To characterize the features of β-caryophyllene as an antioxidant and/or free radical scavenger, both cellular and acellular systems were used. As shown in Table 1, ascorbic acid decreased DCF fluorescence induced by SIN-1 by 92%. β-Caryophyllene decreased SIN-1-induced DCF fluorescence by 13% and 10%, at 10 and 100 μM, respectively.
Intracellular level of ROS was determined by detection of DCF fluorescence in primary cultures of rat cortical neurons and is presented as a percentage of fluorescence intensity with respect to the SIN-1- (200 μM) treated control (n=3 independent experiments).
TEAC represents millimolar concentration of a Trolox solution that has an antioxidant capacity equivalent to a 1 mM solution of β-caryophyllene.
IC50 refers to the concentration of tested compounds that is required for 50% inhibition of radical generation in an acellular system.
Values represent DCF fluorescence (% control) treated with 10 and 100 μM of β-caryophyllene, respectively.
Positive controls used were ascorbic acid for intracellular level of ROS (100 μM), TEAC, and O2 •−, H2O2, HOCl, and ONOO− scavenging; mannitol for HO− scavenging; and quercetin for NO scavenging.
H2O2, hydrogen peroxide; HOCl, hypochlorous acid; O2 •−, superoxide radical; ONOO−, peroxynitrite; NE, not effective; ROS, reactive oxygen species; RNS, reactive nitrogen species; TEAC, Trolox-equivalent antioxidant capacity.
The total antioxidant capacity of β-caryophyllene was monitored by a TEAC assay using an ABTS radical. As shown in Table 1, β-caryophyllene showed little activity, as compared with that of the ascorbic acid (1.00). The radical scavenging activities of β-caryophyllene were measured against OH−, O2 •−, H2O2, HOCl, NO, and ONOO− in the acellular system. Out of six radicals, the scavenging activities were observed against HOCl and ONOO−, with IC50 values of 88.8 and 760 μM, respectively.
Effect on mRNA expression of iNOS, proinflammatory cytokines, and COX in BV-2 microglial cells
To evaluate the effect of β-caryophyllene on the mRNA expression level of the inflammatory mediators, iNOS, TNF-α, IL-1β, IL-6, COX-1, and COX-2, RT-PCR was carried out. A single timepoint of 24 h was chosen for RT-PCR analysis, because this was the timepoint at which the optimal expression of inflammatory mediators was induced by LPS in microglial cells. As presented in Figure 4A, β-caryophyllene decreased the mRNA expression of inflammatory cytokines. In particular, expression of IL-1β and IL-6 mRNA was strongly suppressed by β-caryophyllene in a dose-dependent manner (P<.01). iNOS and TNF mRNA expression was slightly influenced by β-caryophyllene treatment. Another proinflammatory mediator, COX-1, was not affected by β-caryophyllene treatment. By contrast, β-caryophyllene at 100 μM suppressed COX-2 expression induced by LPS. Densitometry analysis of the PCR product indicated a similar effect of β-caryophyllene on inhibiting the expression of IL-6, IL-1β, TNF-α, iNOS, and COX-2, but with no effects on COX-1. The relative intensities inhibited by 10 and 100 μM of β-caryophyllene, as compared with that of LPS-treatment, were iNOS: 85%, 78%; TNF-α: 99%, 83%; IL-1β: 77%, 58%; IL-6: 71%, 39%; COX-1: 91%, 87%; and COX-2: 98%, 56%. The expression of iNOS, IL-1β, and IL-6 was significantly decreased by 10 μM β-caryophyllene, and that of iNOS, TNF-α, IL-1β, IL-6, and COX-2 was significantly decreased by 100 μM β-caryophyllene (Fig. 4B).
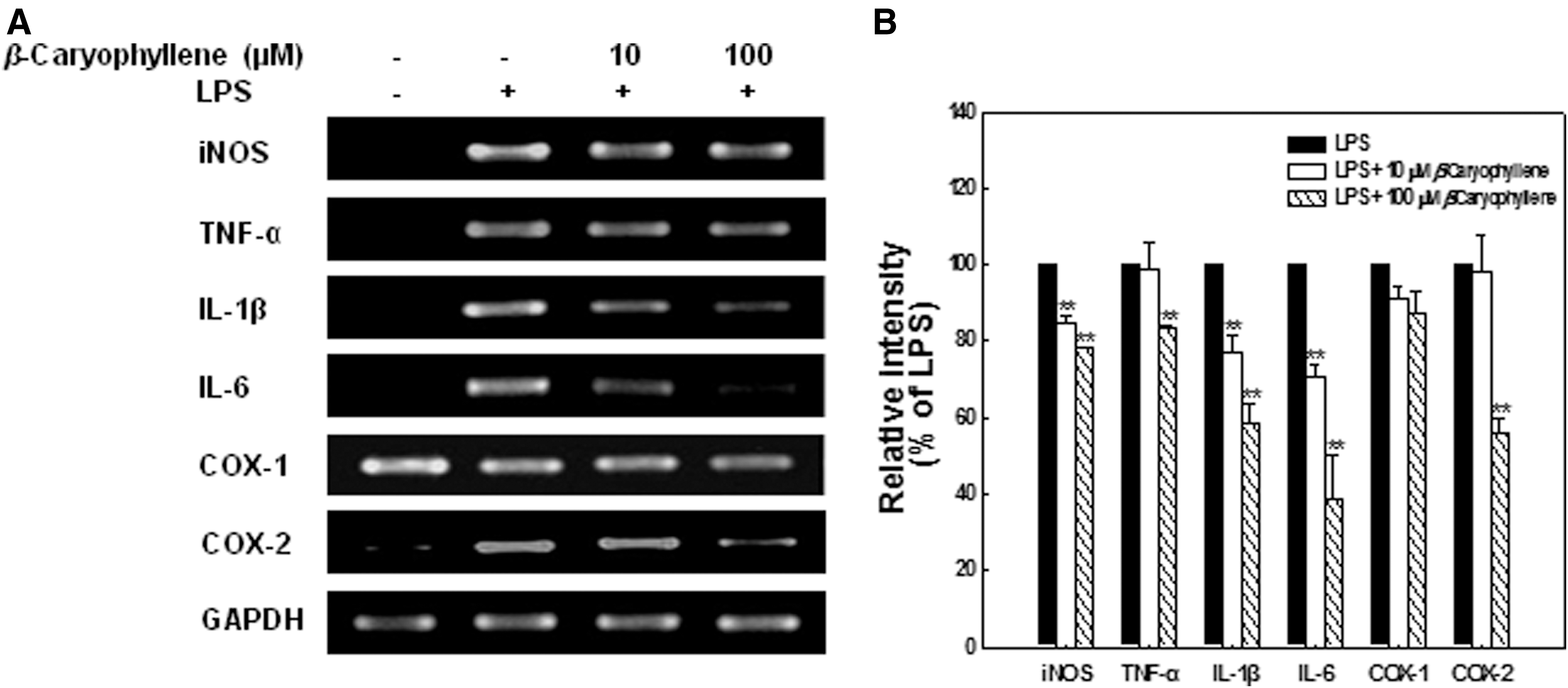
Effect of β-caryophyllene on mRNA expression of iNOS and proinflammatory cytokines in BV-2 microglial cells stimulated by LPS.
Effect on the production of NO, proinflammatory cytokines, and PGE2 in BV-2 microglial cells
To determine the inhibitory effect of β-caryophyllene on the production of NO and proinflammatory cytokines (TNF-α, IL-1β, and IL-6) in BV-2 microglial cells, the cells stimulated by LPS for 48 h and those secreted into the culture medium were measured (Fig. 5). NO production in BV-2 microglial cells stimulated by LPS was dose-dependently and significantly decreased at 48 h after treatment with 10 or 100 μM β-caryophyllene (Fig. 5A). LPS-induced IL-1β secretion was slightly inhibited by 10 μM β-caryophyllene (Fig. 5C, D), whereas LPS-induced IL-6 and TNF-α productions were not significantly inhibited (Fig. 5B). LPS-induced PGE2 production was also significantly decreased from 32.5±4.00 pg/mL to 16.2±1.88 pg/mL by 100 μM β-caryophyllene (Fig. 5E).
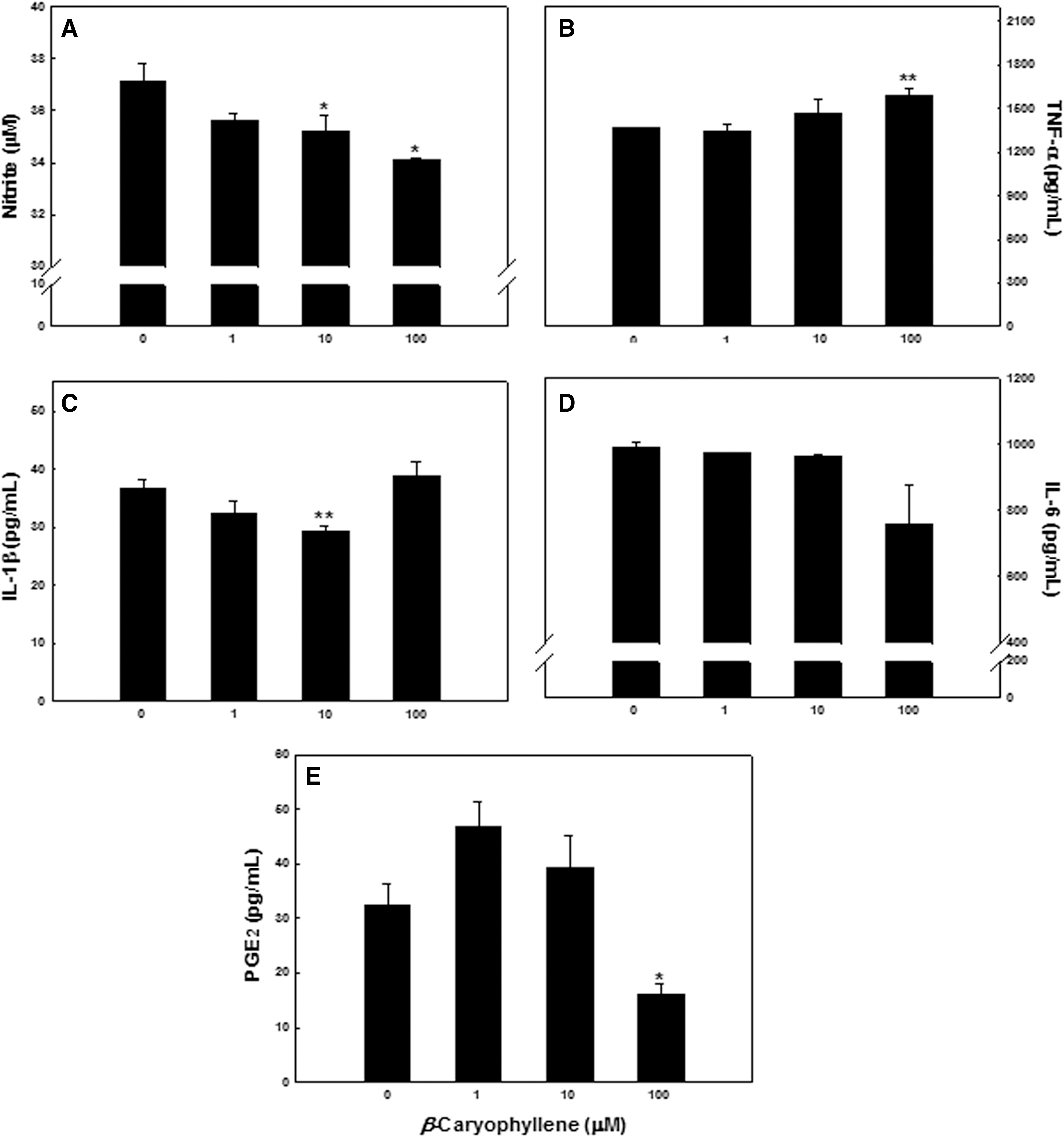
Effect of β-caryophyllene on the production of NO and proinflammatory cytokines in BV-2 microglial cells stimulated by LPS. Microglial cells were treated with LPS 100 ng/mL in the absence or presence of indicated amounts of β-caryophyllene for 48 h. The amount of
Discussion
The present study demonstrated that β-caryophyllene, a natural bicyclic sesquiterpene, can inhibit cortical neuronal damage when administered before the initial insult in the rat MCAO/reperfusion stroke model. Moreover, our results suggest that β-caryophyllene-induced neuroprotection may be partly related to the modulation of inflammatory mediators, although the molecular mechanism underlying its neuroprotection still remains unclear.
β-Caryophyllene significantly decreased the infarct volume in the rat brain exposed to transient MCAO/reperfusion and also cell death in the culture of rat cortical cells. Additionally, β-caryophyllene-treated rats demonstrated improved neurologic function after MCAO. It is well known that one of the major causes of neuronal death during or after OGD/reoxygenation includes excitotoxicity through overstimulation of glutamate receptor subtypes, including NMDA, an analog of excitatory amino acid neurotransmitters. 28,29 However, our present results demonstrated that β-caryophyllene did not inhibit NMDA excitotoxicity.
In cerebral hypoxic/ischemic insults, ROS are produced in a large amount, leading to massive injury of brain tissue. 30,31 Consistent with this finding, various kinds of antioxidants have been found to prevent ischemic brain damage. 32,33 Thus, we hypothesized that β-caryophyllene might ameliorate ischemia-induced neuronal cell death, possibly owing to its antioxidant activity. In this study, however, β-caryophyllene exhibited only weak antioxidant and scavenging effects on OH•, O2 •−, H2O2, HOCl, NO, and ONOO−. Further, β-caryophyllene neither directly scavenged the ABTS+ nor suppressed the production of ROS induced by SIN-1 in microglial cells. Therefore, these findings indicate that the antioxidant properties of β-caryophyllene may not play a crucial role in its neuroprotective action in neuronal cells exposed to ischemia/reperfusion and OGD/reoxygenation. The level of ONOO− has also been shown to increase markedly during an ischemic insult. 34,35 In the present study, even at a concentration as high as 100 μM, β-caryophyllene did not decrease the level of intracellular ROS, including ONOO− in microglial cells or ONOO− in a cell-free system generated from SIN-1. Thus, it is unlikely that β-caryophyllene exerts protective activity against cerebral ischemic injury via its antioxidative action.
There is extensive evidence indicating that neuroinflammation contributes to the pathogenesis of stroke. 1,5 The cellular mediators of innate immune responses are microglial cells, whose activation results in the production of several inflammatory molecules that contribute to neurodegeneration in the brain. There have been many epidemiological studies showing that long-term treatment with nonsteroidal anti-inflammatory drugs (NSAIDs) reduces the risk of neurodegenerative disease, delays disease onset, ameliorates symptomatic severity, and slows cognitive decline. 36
We therefore investigated the effect of β-caryophyllene on the production of NO and inflammatory cytokines in BV-2 microglial cells. Treatment with β-caryophyllene was found to decrease NO production at doses of 10 and 100 μM. In parallel with the decreased production of inflammatory mediators, the mRNA expression measured by a semiquantitative RT-PCR was also found to be slightly decreased by β-caryophyllene treatment. LPS-induced IL-1β and IL-6 mRNA expression was markedly inhibited by 10 and 100 μM β-caryophyllene, but their corresponding protein levels only slightly affected, if at all. Moreover, modulation of the TNF-α protein and mRNA levels by β-caryophyllene treatment was not consistent in terms of dose dependency, for which we do not have an adequate explanation. All together, these findings suggest that β-caryophyllene may have differential effects on the production of numerous proinflammatory mediators during activation of microglial cells.
COX-2 is highly expressed in the ischemic brain and produces proinflammatory prostaglandins such as PGE2. 5,7 In general, the expression of COX-2 is decreased by anti-inflammatory agents, but can be induced by cytokines, including IL-1β. 7 Because β-caryophyllene treatment inhibited the production of NO and inflammatory cytokines, we studied whether the treatment also affected PGE2 and COX-2 in BV-2 microglial cells. Treatment with 100 μM β-caryophyllene decreased the PGE2 concentration and inhibited the COX-2 expression by ∼50% in BV-2 microglial cells. Importantly, however, β-caryophyllene did not affect COX-1 expression. This property would be valuable because a major limitation of NSAIDs with regard to the prevention of neurodegenerative diseases is gastrointestinal toxicity, as well as occasional liver and kidney toxicities, caused by COX-1 inhibition. 36 Taken together, these findings indicated that β-caryophyllene exerted a less-potent anti-inflammatory activity, implying that the observed anti-inflammatory property may not play a crucial role in its neuroprotective action in cerebral ischemic injury.
β-Caryophyllene was granted the GRAS status by the FEMA and is approved by the U.S. FDA for food use 8,9 because of its low toxicity. β-Caryophyllene did not cause skin irritation or sensitization in human subjects, at concentrations up to 4%. Both the acute oral LD50 in rats and the acute dermal LD50 in rabbits exceeded 5 g/kg, 8 and intratracheal doses at 12, 24, or 48 mg/kg of β-caryophyllene were not toxic to the lungs of F344 male rats. 37 β-Caryophyllene was not mutagenic in Salmonella typhimurium strains or in an unscheduled DNA synthesis assay, at concentrations up to 150,000 μg/plate and 10,000 μg/mL. 38 Consequently, β-caryophyllene may serve as a potential parent compound for the development of safe therapeutics for ischemic stroke.
In conclusion, our data indicate that treatment with β-caryophyllene decreased ischemia/reperfusion-induced cortical cell death, production, and expression of inflammatory mediators (NO and PGE2/COX-2) in vitro, and cortical infarction and neurological deficit in vivo. However, β-caryophyllene neither suppressed NMDA-induced excitotoxicity nor reduced the oxidative stress caused by OGD/reoxygenation in the cortical cells or in the acellular system. These data suggest that β-caryophyllene protects neurons from ischemia-induced cell death and inhibits inflammatory responses to some extent. However, the mechanism underlying the neuroprotection of β-caryophyllene still remains unclear. To our knowledge, this is the first report demonstrating that β-caryophyllene exerts neuroprotective activity against in vitro and in vivo ischemic insults. Further studies will be required to examine the therapeutic window and the dose–response effect of β-caryophyllene, as well as the mechanism of action.
Footnotes
Acknowledgments
This work was supported by a grant from the Agriculture R&D Promotion center, under the Ministry of Agriculture and Forestry, and in part by a grant from the National Research Foundation for functional food research and development, Ministry of Science and Technology, in the Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.