
Abstract
Although Alpinia officinarum has been used in traditional medicine for the treatment of several conditions, such as abdominal pain, emesis, diarrhea, impaired renal function, and dysentery, little is known about its function in obesity. In this study, we investigated the antiobesity effect of A. officinarum ethanol extract (AOE) on lipid accumulation in 3T3-L1 cells and obesity in mice fed a high-fat diet (HFD). AOE dose-dependently suppressed lipid accumulation during differentiation of 3T3-L1 preadipocytes by downregulating CCAAT enhancer binding protein α (C/EBPα), sterol regulatory element binding protein-1 (SREBP-1), and peroxisome proliferator-activated receptor-γ (PPAR-γ) genes. Galangin, a major component of A. officinarum, had antiadipogenic effects in 3T3-L1 cells. AOE supplementation in mice fed a HFD revealed that AOE significantly decreased HFD-induced increases in body, liver, and white adipose tissue weights and decreased serum insulin and leptin levels. To elucidate the inhibitory mechanism of AOE in obesity, lipid metabolism-related genes were identified. AOE efficiently suppressed protein expressions of C/EBPα, fatty acid synthase, SREBP-1, and PPAR-γ in the liver and adipose tissue. The protein expression patterns, observed by immunoblot, were confirmed by quantitative real-time polymerase chain reaction. Collectively, these results suggest that AOE prevents obesity by suppressing adipogenic and lipogenic genes. AOE has potential for use as an antiobesity therapeutic agent that can function by regulating lipid metabolism.
Introduction
O
Alpinia officinarum Hance, which belongs to the ginger family (Zingiberaceae) and is commonly known as galangal, is popular in Southeastern Asia as a traditional medicine for ailments, such as abdominal pain, emesis, diarrhea, stress-induced impotence, impaired renal function, and dysentery. 7 On the basis of the presence of active compounds, such as diaryl heptanoids, flavonoids, and essential oils, several studies have shown that the extract of A. officinarum inhibits fatty acid synthase (FAS) activity, 5α-reductase activity, inflammation, and lipid oxidation. 8 –12 In spite of its reported utility in many medicinal applications, the potential of A. officinarum as an antiobesity agent has not yet been rigorously investigated. Although Xia and colleagues recently showed that a 70% ethanol extract from A. officinarum can reduce body weight in rats fed a high-fat diet (HFD), the mechanisms involved in the prevention of obesity by A. officinarum have not yet been explored. 13
In the present study, we determined the effects of A. officinarum on the differentiation of 3T3-L1 preadipocytes and its in vivo inhibition of HFD-induced obesity in mice. To clarify the mechanism underlying the regulation of obesity, the expression of lipid metabolism-related genes in the liver and in adipose tissue were analyzed.
Materials and Methods
Preparation of A. officinarum ethanol extract
A. officinarum plants were purchased from the Omni Herb Company (Daegu, South Korea) and the voucher specimen (A011) was deposited in the herbarium of the Korea Food Research Institute. The material was authenticated by professor Seong-Gyu Ko of the College of Oriental Medicine, Kyung Hee University, Seoul, Korea. The dried A. officinarum plants were ground in a mill and passed through a 50-mesh sieve. One hundred grams of A. officinarum was subjected to extraction three times with 70% ethanol by repeat sonication, and then filtered. The ethanol extracts were combined and concentrated in a vacuum evaporator and freeze-dried to obtain a dry residue. The extraction yield of A. officinarum ethanol extract (AOE) was 22.1% (w/w).
Chemicals and reagents
The Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), bovine calf serum (CS), sodium pyruvate, and penicillin–streptomycin were obtained from Gibco BRL (Grand Island, NY, USA). Antibodies to CCAAT enhancer binding protein α (C/EBPα, sc-61) and sterol regulatory element binding protein-1 (SREBP-1, sc-367l) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). FAS (#3180), peroxisome proliferator-activated receptor-γ (PPAR-γ, #2443), β-actin (#4967), and secondary antibodies were purchased from Cell Signaling (Danvers, MA, USA). Oil Red O (#O0625), isobutylmethylxanthine (IBMX; #I5879), dexamethasone (#D1756), and galangin (#282200) were purchased from Sigma-Aldrich (St Louis, MO, USA). Triglyceride (TG, #21001) and total cholesterol (TC, #1Y004) kits were purchased from Shinyang Diagnostics (Seoul, South Korea). Insulin (#80-INSMSU-E01) and leptin (#22-LEPMS-E01) enzyme-linked immunosorbent assay (ELISA) kits were obtained from Alpco Diagnostic (Windham, ME, USA).
Cell culture and differentiation
3T3-L1 preadipocyte cells were cultured in DMEM with 1% penicillin–streptomycin and 10% CS at 37°C in 5% CO2. The cells were seeded at a density of 4×105 cells/well into a six-well plate. At 2 days postconfluence (day 0), the cells were exposed to an IBMX–dexamethasone–insulin (MDI) solution containing 0.5 mM of IBMX, 1 μM of dexamethasone, and 1 μg/mL of insulin for 2 days in DMEM with 10% FBS. After induction, the cells were switched to a solution containing DMEM supplemented with 10% FBS and 100 nM insulin for 2 days. The cells were maintained in a postdifferentiation medium (DMEM with 10% FBS) and changed every 2 days. To examine the effects of AOE on the differentiation of preadipocytes into adipocytes, the cells were treated with various concentrations of AOE or galangin at 2 days postconfluence (day 0).
Cell viability assay
The 3T3-L1 cells were seeded into a 24-well plate at a concentration of 2×104 cells/well. After 24 h of preconditioning, the cells were exposed to a variety of concentration of galangin for 48 h. Subsequently, 50 μL of cell counting kit-8 (#CK04, Dojindo, Kumamoto, Japan) solution was added into each wells and further incubated for 3 h to detecting cell survivals at 37°C. The cell viability was calculated by measuring the absorbance using a micro plate reader (Infinite M200; Tecan, Männedorf, Switzerland) at 450 nm.
Oil Red O staining
For quantification, the cells were fixed with 10% neutral formalin for 1 h at room temperature, washed with phosphate-buffered saline, and stained for 1 h with 0.5% Oil Red O in 60% isopropanol. After the stained cells were washed with distilled water, they were observed under an inverted microscope. The stained lipid droplets were extracted with isopropanol, followed by quantification by measuring absorbance at 490 nm.
Animals and experimental protocol
Four-week-old male C57BL/6J mice were purchased from Orient Bio, Inc. (Seoul, South Korea) and acclimatized for 1 week before being randomly assigned to experimental groups. The animals were housed in individual cages with free access to a regular diet and water in a room with a 12-h light cycle, a temperature of 24°C, and a humidity of 55%. The mice (5 weeks old) were randomly divided into three groups: a control group fed the American Institute of Nutrition AIN-76A diet (normal diet), an experimental group fed a HFD (the HFD group), and an experimental group fed a HFD with 0.5% AOE (the HFD+AOE group) for 8 weeks. The experimental diets were based on the AIN-76 diet, which contained 20% fat and 0.5% cholesterol (w/w) (Table 1). Body weights and average daily food intake were measured weekly. The experimental protocols were performed in accordance with the Institutional Animal Care and Use Committee of Korea Food Research Institute.
AIN-76 mineral and vitamin mixture.
AOE, Alpinia officinarum ethanol extract; C, control (AIN-76A diet); HFD, high-fat diet; HFD+AOE, high-fat diet+AOE supplementation.
Sample preparation
After the mice consumed the experimental diets for 8 weeks, they were sacrificed after a 12-h fast. Blood samples were collected from the abdominal aorta, centrifuged at 1000 g for 15 min, and stored at −80°C. Epididymal, perirenal, brown fat, and liver tissues were excised, weighed, and stored at −80°C until use. Liver and epididymal fat pads were either fixed in 4% formalin solution and processed for histological analysis or snap-frozen in liquid nitrogen and stored at −80°C until RNA extraction. The serum insulin and leptin concentrations were measured according to the protocol described by the manufacturers of the mouse insulin and leptin ELISA kits.
Hepatic lipid profiles
To determine liver total lipid, TG, and TC levels, the livers from each group of mice were homogenized in 5 mL NaCl. A 20-mL volume of Folch solution (chloroform:methanol=2:1) was added to each homogenate, and the solution was incubated at 4°C for 12 h. The sample was centrifuged for 10 min at 1000 g. The organic layer was removed and dried. The resulting pellet was dissolved in ethyl alcohol containing 25% Triton X-100, and then assayed for TG and TC by using commercial kits.
Morphologic evaluation
For histopathologic analysis, liver and epididymal fat pads fixed in 4% neutral-buffered formalin were embedded in paraffin, sliced into 5-μM-thick slices, and stained using the hematoxylin and eosin stain. Pathological changes were assessed and photographed under an Olympus BX-51 microscope (Olympus, Tokyo, Japan).
Western blotting
The liver and adipose tissues were homogenized in the PRO-PREP protein extraction solution (iNtRON Biotechnology,Gyeonggi, South Korea) using an MP 40 system (MD Biosciences, St. Paul, MN, USA). The homogenates were then centrifuged at 16,000 g for 20 min at 4°C. The supernatants were boiled for 5 min at 100°C with a sodium dodecyl sulfate (SDS)-loading buffer. Samples were separated by SDS–polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. Immunoblot analysis was performed with antibodies, FAS, SREBP-1, C/EBPα, and PPAR-γ, and was visualized by chemiluminescence by using an ECL kit (Amersham Bioscience, Piscataway, NJ, USA).
RNA extraction and real-time polymerase chain reaction
Total RNA was isolated from the liver by using the Trizol reagent (Life Technologies, Carlsbad, CA, USA) according to the manufacturer's protocol. After cDNA was prepared from isolated RNA by using a Maxime RT premix (iNtRON), the cDNA was used as a template for real-time polymerase chain reaction (RT-PCR) with the Light Cycler 480 System (Roche). The specific primers are given in Table 2. The mRNA level was expressed as the ratio of signal intensity for each gene, relative to that of β-actin.
High-performance liquid chromatography analysis
The galangin from AOE was analyzed using a Jasco high-performance liquid chromatography (HPLC) system consisting of a Jasco PU-2089 plus quaternary gradient pump, a Jasco AS-2057 plus intelligent sample injector, a Jasco UV-2075 intelligent UV/VIS detector, and Borwin chromatography software version 1.5. Separation was performed using a 5.0 μm, 4.6 mm (i.d.)×250 mm XTerra RP18 column (Waters, Milford, MA, USA) and a guard column. The analyses were performed using a mobile phase consisting of two solvents: solvent A (water containing 0.1% formic acid) and solvent B (methanol). The flow rate was 1.0 mL/min. The elution was performed using a programmed gradient elution as follows: 0–7 min isocratic elution with 30% B, 7–25 min elution with 30–35% B, 25–45 min elution with 35–40% B, 45–50 min elution with 40–50% B, 50–65 min elution with 50–100% B. Galangin was detected at 350 nm.
Statistical analysis
All data are expressed as mean±standard deviation values. The differences among the 3 groups were evaluated by analysis of variance followed by the Duncan's multiple range test. P values less than .05 were considered statistically significant.
Results
AOE inhibits cell differentiation in 3T3-L1 cells
To investigate the effect of AOE on cell differentiation, the cell viability of AOE was first determined. The results showed that AOE had no significant cytotoxicity up to 0.4 mg/mL (Fig. 1A). Oil Red O staining was used to determine whether AOE inhibits the MDI-induced differentiation of 3T3-L1 preadipocytes. AOE treatment also significantly inhibited 3T3-L1 adipocyte differentiation in a dose-dependent manner (Fig. 1B, C). The AOE expression levels of C/EBPα, SREBP-1, and PPAR-γ, which promote adipogenesis were also examined, and each was dose-dependently suppressed by AOE treatment (Fig. 1D).
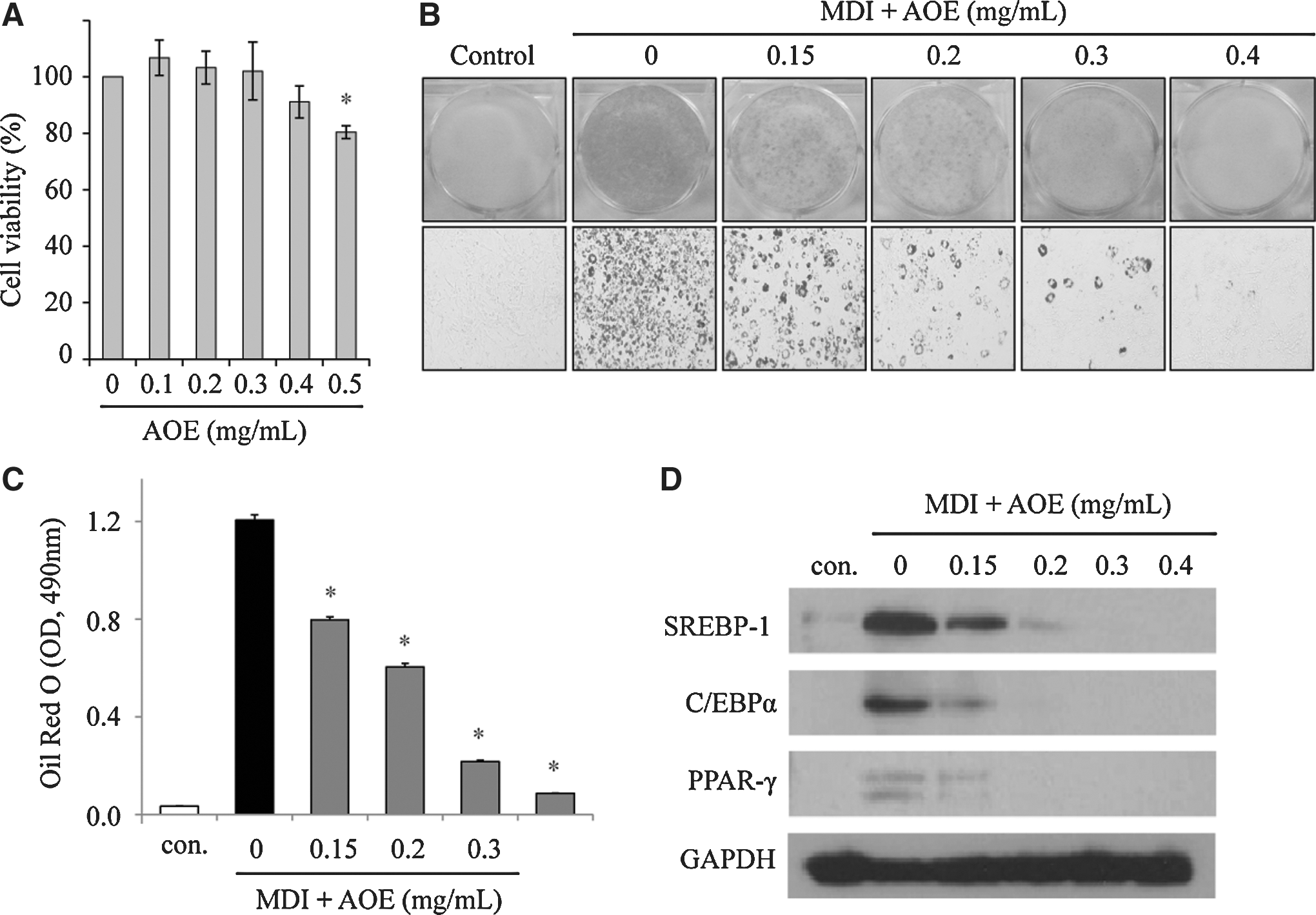
Alpinia officinarum inhibits adipogenic differentiation in 3T3-L1 cells.
Galangin, a major component of AOE, prevents differentiation of 3T3-L1preadipocytes
AOE was analyzed using HPLC to identify the primary component of AOE; galangin was identified as a representative compound (Fig. 2A). The amount of galangin in AOE was 4.2% (w/w). We tested whether galangin can inhibit adipocyte differentiation in 3T3-L1 cells. Galangin had no significant cytotoxic effects up to 50 μM (Fig. 2B). Thus, a concentration of less than or equal to 50 μM was used to examine the effects of galangin on cell differentiation. Galangin was also found to inhibit TG accumulation during cell differentiation (Fig. 2C). The protein levels of several adipogenic genes, including FAS, C/EBPα, CD36, and PPAR-γ, were significantly downregulated by galangin in a dose-dependent manner (Fig. 2D). These results suggest that galangin can be expected to be effective against obesity.
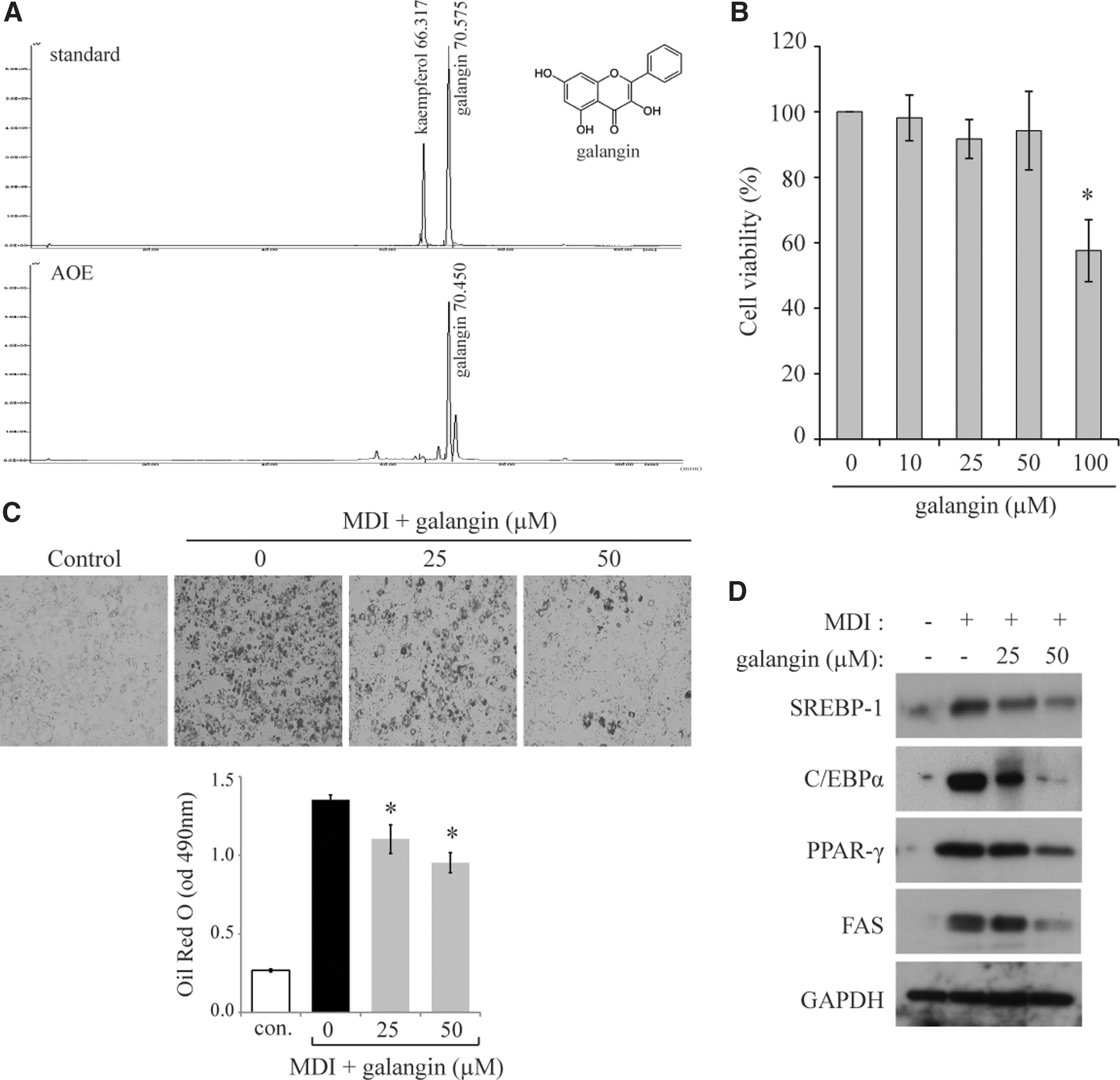
Galangin, major component of AOE, inhibits cell differentiation in 3T3-L1 cells.
AOE reduces body weight and fat mass in HFD-fed mice
To determine the antiobesity effect of AOE in HFD-fed mice, the weights of the body, liver, white adipose tissue (WAT), and brown adipose tissue after 8 weeks of supplementation were measured. Interestingly, despite no significant differences in the daily food intake between the HFD and HFD+AOE groups, the HFD+AOE group had a significantly lower body weight than the HFD group (P<.05) (Fig. 3A, Table 3). Remarkably, decreased weights of the epididymal and perirenal fat pads were also observed when the mice were fed a diet supplemented with AOE, indicating that AOE inhibited fat accumulation. Histologic analysis confirmed that the areas of lipid droplets in AOE-supplemented mice were smaller than those in mice fed only a HFD (Fig. 3B). In addition to the data on hepatic lipid profiles, histologic analysis supported the effectiveness of AOE in reducing fat accumulation in the liver (Fig. 3B). Collectively, these data suggest that AOE supplementation effectively reduces liver fat accumulation in HFD-fed obese mice.
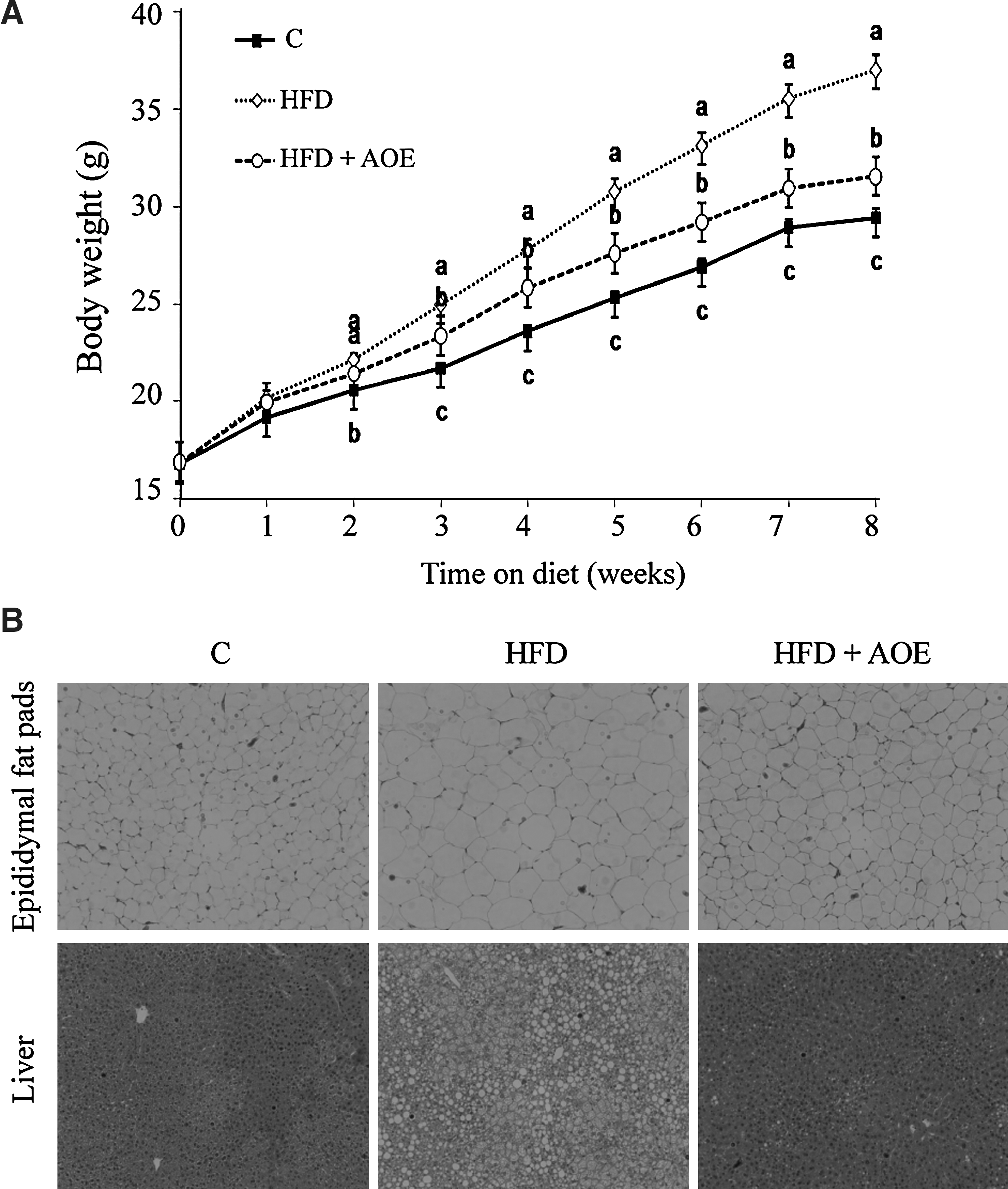
A. officinarum reduces body weight, white adipose tissue (WAT) mass, and fatty liver in high-fat diet (HFD)-fed mice.
Data are expressed in terms of mean±standard deviation (SD) values (in g).
Data in a single row that do not share common superscript letters represent significant differences (P<.05).
AOE reduces serum insulin and leptin levels in HFD-fed mice
In a manner similar to the increases observed in body weight and fat mass, serum insulin and leptin levels also increased significantly with HFD feeding (P<.05, Fig. 4A, B). AOE supplementation significantly reduced the levels of both insulin and leptin, suggesting that decreased leptin and insulin levels may indicate depleted fat stores and lower blood glucose levels.
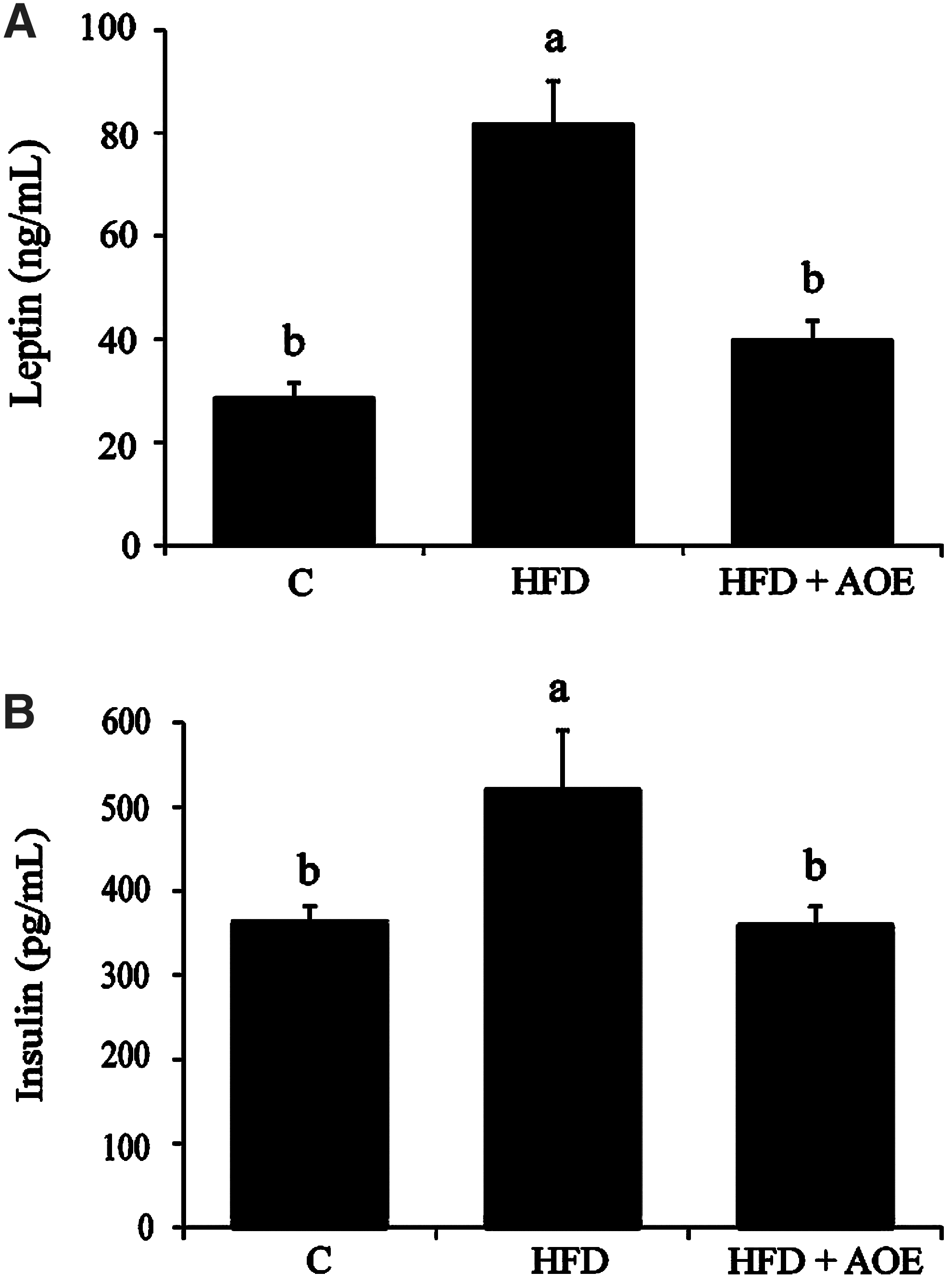
A. officinarum improves serum insulin and leptin levels in mice fed a HFD. Serum insulin
AOE decreases hepatic lipid as well as TG and TC levels in HFD-fed mice
We next examined the effect of AOE on total lipid, TG, and TC levels in the liver in HFD-fed mice. The HFD+AOE treated group had lower total hepatic lipid content compared to the control HFD group. The hepatic TG concentration in HFD-fed mice increased by 2.2-fold relative to that in normal-diet mice, and the TG level decreased by 17% in mice fed HFD with AOE (Table 4). The TC levels in the HFD+AOE group were also significantly lower than those in the HFD group, suggesting that AOE may exert hypolipidemic activity in HFD-fed mice.
Data are expressed in terms of mean±SD values (in mg/g).
Data in a single row that do not share common superscript letters represent significant differences (P<.05).
TG, triglyceride; TC, total cholesterol.
AOE regulates genes related to lipid metabolism of the liver and the epididymal fat pads in HFD-fed mice
The results from Western blotting indicated that AOE decreased the expression levels of SREBP-1, C/EBPα, and PPAR-γ in the epididymal fat pads of HFD-fed mice (Fig. 5A). AOE treatment also suppressed hepatic lipid metabolism-related genes, as determined by the levels of SREBP-1, FAS, C/EBPα, and PPAR-γ (Fig. 5B). RT-PCR assays showed that, with AOE treatment, the mRNA levels of Srebp1, Fas, PEPCK, G6Pase, and Pparg decreased by 53%, 37%, 56%, 60%, and 25%, respectively (P<.05), relative to those in mice fed with HFD alone (Fig. 5C). These results suggest that AOE improves lipid metabolism through altered transcriptional regulation.
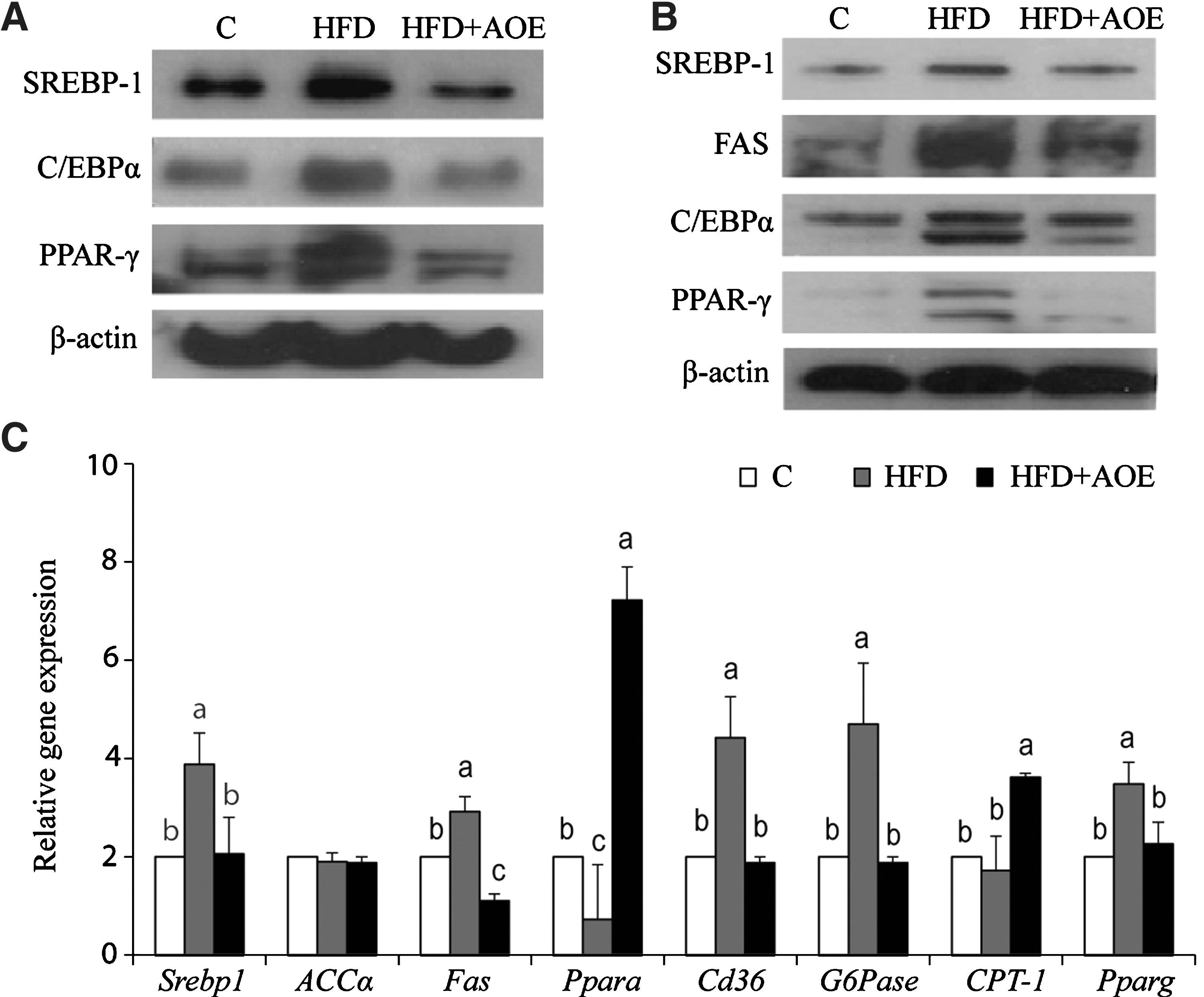
A. officinarum regulates adipogenesis and lipogenesis in the WAT and the livers in mice fed a HFD. The protein levels of adipogenic and lipogenic genes in epididymal fat pads
Discussion
Herbal medicines are being increasingly used as antiobesity treatments with little understanding of the mechanisms underlying their therapeutic effects. A. officinarum, a plant in the ginger family, is used as a spice and a traditional herbal medicine. 7 While a recent study has shown that AOE has an antiobesity effect in rats fed a HFD, the exact mechanism by which AOE achieves these effects remains unknown. 13 In the present study, we found that AOE reduces HFD-induced increases in body weight, WAT, and liver weight through suppression of adipogenesis and lipogenesis.
In cultured adipocyte models, AOE has been found to inhibit adipocyte differentiation in 3T3-L1 cells in a dose-dependent manner. Adipocyte differentiation of 3T3-L1 preadipocytes has been found to depend on adipo-specific genes, including C/EBPα and PPAR-γ, during the differentiation process. 14 It has been suggested that SREBP-1 regulates genes required for adipogenesis and that its expression is regulated by insulin. 15 Our results showed that AOE suppressed the adipogenic genes, C/EBPα, PPAR-γ, and SREBP-1. Collectively, these data suggest that AOE inhibits cell differentiation of 3T3-L1 preadipocyte cells through suppression of these genes.
This study presents several lines of evidence demonstrating that AOE has antiobesity effects in HFD-fed mice. First, AOE reduced body weight. Second, the levels of serum insulin and leptin, which reflect growth of adipose tissue in obese animals, were significantly lowered by AOE treatment; suggesting that AOE can improve serum insulin and leptin resistance in HFD-fed mice. Third, AOE treatment of HFD-fed mice decreased the WAT fat mass. This is consistent with previous observations that hyperinsulinemia and hyperleptinemia initiate fat accumulation in the body. Finally, AOE reduced hepatic lipid accumulation in HFD-induced obese mice. Taken together, these results indicate that AOE efficiently improves lipid accumulation in HFD-induced obese mice. Our data provide solid evidence of an antiobesity effect in an animal model.
Altered lipid metabolism through AOE supplementation in the liver and the epididymal fat pads suggests that the mechanism may relate to changes in transcription levels that regulate adipogenesis and lipogenesis. Consistent with cellular observations, AOE suppressed the expression of SREBP-1, C/EBPα, and PPAR-γ. These genes are expressed in adipose tissue and the liver as key transcription regulators of TG synthesis and fatty acid metabolism. 16,17 These results suggest that inhibition of fat accumulation in the liver and adipose tissue is responsible for the beneficial effects of dietary AOE through suppression of gene expression related to lipid metabolism. The antiobesity effects of AOE are further supported by the results of the mRNA analysis. The mRNA level of Pparg was significantly downregulated by AOE, whereas gene expression levels of major lipolytic regulators, such as Ppara and CPT-1, were significantly upregulated. These effects may be due to the combined effects of Ppara activation and downregulation of Pparg and Cd36. The phenotypes produced by AOE could be mediated by changes in lipolysis or through its effects on adipogenesis. This study indicates that AOE can directly regulate adipogenesis and lipogenesis.
Numerous studies have indicated that various natural extracts inhibit lipid accumulation in vitro and in vivo. This study showed that AOE has an inhibitory effect on HFD-induced obesity. Although several studies have reported that AOE exerts a variety of pharmacologic actions, including anticancer and antibacterial activities, 18,19 no study has examined the action of A. officinarum in the prevention of obesity in mice fed a HFD. Several studies have suggested that active components of A. officinarum are diarylheptanoids, flavonoids, and essential oils and that these compounds show 5-reductase inhibitory, COX-2 inhibitory, antiemetic, antigenotoxic, and antioxidative activities. 20 –22 Interestingly, we found that galangin, a major component of A. officinarum, can prevent cell differentiation in 3T3-L1 cells. This mechanism is believed to be active in preventing obesity, although the activity of galangin in animal models was not investigated. The present study demonstrates that AOE and its active compound, galangin, may be appropriate for development as therapeutic agents for the treatment of obesity. These treatments would operate via a mechanism involving downregulation of C/EBPα, PPAR-γ, and SREBP-1 in the liver and adipose tissue.
This study provides solid evidence that AOE has potent antiobesity activity in vitro and in vivo. The mechanism behind the antiobesity activity of AOE appears to be a reduction in lipid accumulation via inhibition of genes related to lipid metabolism in the liver and adipose tissue. Galangin, a major component of AOE, can inhibit adipogenesis during adipocyte differentiation and may be effective in improving obesity.
Footnotes
Acknowledgments
This study was supported by the National Platform Technology Project from the Ministry of Knowledge Economy in Korea, and the Korea Food Research Institute.
Author Disclosure Statement
No competing financial interests exist