
Abstract
Polyphenol-rich foods, such as fruits and vegetables, are protective against cardiovascular diseases, but the mechanisms of the beneficial effects are still unknown. The goal of this research was to clarify actions of procyanidin trimer (C1) in rat aortic endothelial cells (RAECs). Procyanidin C1 at concentrations up to 50 μM was not cytotoxic to the RAECs. The addition of procyanidin C1 to RAECs exerted a time-dependent hyperpolarization measured using a membrane potential-dependent fluorescent probe, bis-(1,3-dibutylbarbituric acid) trimethine oxonol, whereas the hyperpolarization was significantly inhibited by the nonspecific K+ channel inhibitor tetraethylammonium chloride (TEA). Moreover, procyanidin C1 elevated intracellular Ca2+ influx, which was totally abolished in the presence of Ca2+-free solution with EGTA. Procyanidin C1 caused a significant increase in nitric oxide (NO) production. The effect was significantly inhibited by an NO synthase inhibitor, N
G-monomethyl-
Introduction
C
Epidemiological studies have shown an inverse correlation between intake of polyphenol-rich foods and risk of CVD characterized by an endothelial dysfunction. 4 In this regard, polyphenols, containing several hydroxyl groups directly associated with a cyclic benzene ring, are widely distributed in plants, 5 and there have been several reports on the role of polyphenolic compounds from various sources such as red wine, grape juice, and tea in the regulation of vascular tone. 6 –8
Vasorelaxation is an important therapeutic strategy against the development of CVD associated with endothelial dysfunction 9 and appears to be mediated by nitric oxide (NO) production through the activation of intracellular Ca2+ ([Ca2+]i) in endothelial cells. 10 Previous studies have revealed that potential bioactivities contributing to the beneficial effect (e.g., vasorelaxation) of polyphenolic compounds include endothelium-derived hyperpolarizing factor (EDHF)-mediated responses such as hyperpolarization via increased [Ca2+]i action in endothelial cells. 11,12 It has been recently shown that EDHF exerts its effect by increasing [Ca2+]i levels and subsequent activation of Ca2+-sensitive potassium channels that cause vasorelaxation. 13 In addition, it has been demonstrated by Tokoudagba et al. 14 that procyanidin-rich fractions play a key role in the control of vascular tone via the release of potent vasorelaxing factors such as NO and EDHF. However, to the best of our knowledge, the mechanism or mechanisms underlying vasorelaxation effect by procyanidin trimer (C1) (Fig. 1) in endothelial cells remains unclear. Accordingly, the objective of the present study was to clarify whether procyanidin C1 affects the activation of NO production or hyperpolarization in rat aortic endothelial cells (RAECs) and, if so, to characterize the underlying mechanism of the Ca2+-dependent pathway.
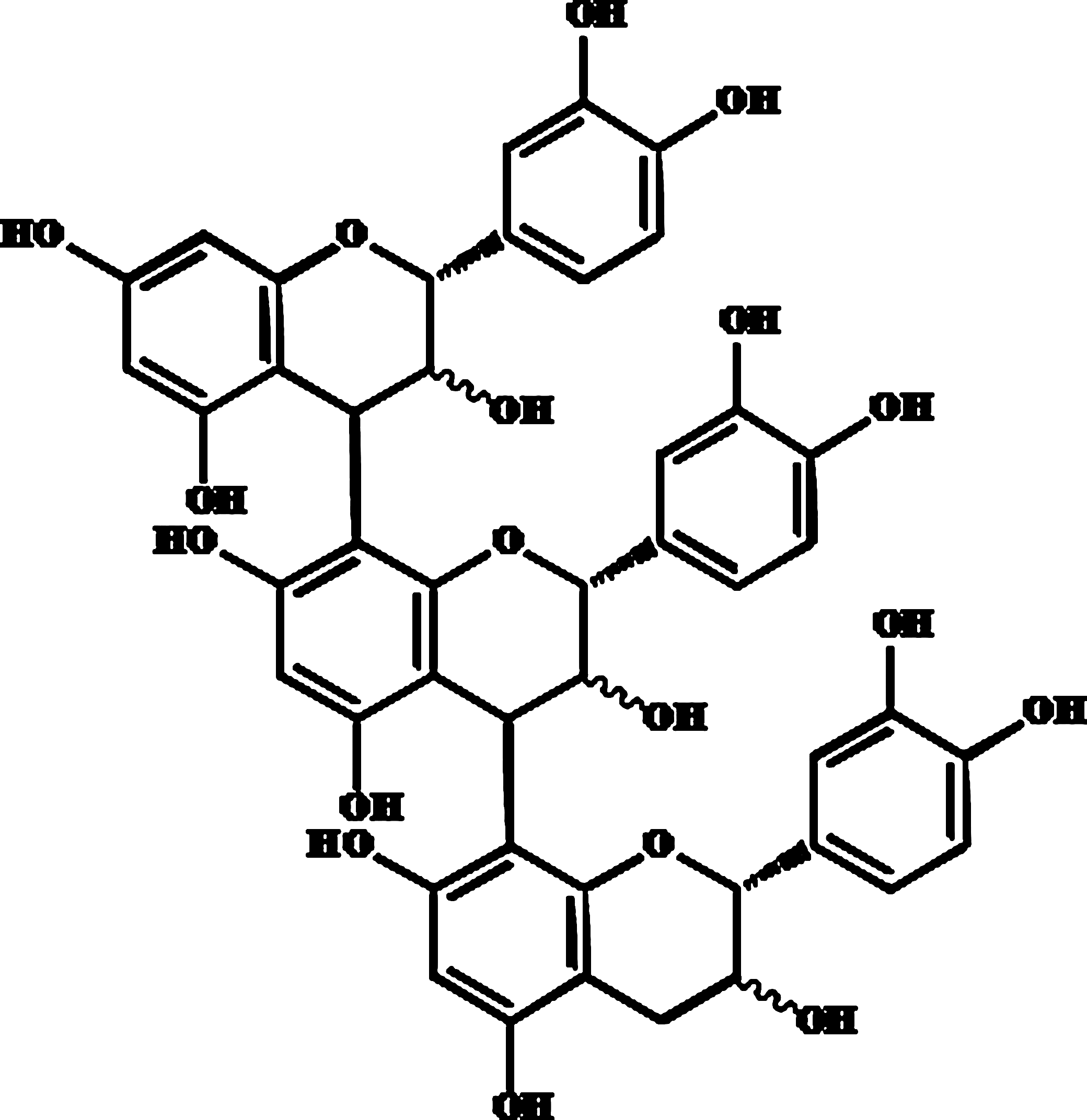
Chemical structure of procyanidin trimer (C1) [epicatechin-(4β→ 8)-epicatechin-(4β→ 8)-epicatechin] used in the study.
Materials and Methods
Materials
Procyanidin C1 was purchased from PhytoLab GmbH (Vestenbergsgreuth, Germany). Tetraethylammonium chloride (TEA) was obtained from Sigma-Aldrich (St. Louis, MO, USA). N
G-Monomethyl-
Cell culture
RAECs were purchased from Cell Applications Inc. (San Diego, CA, USA). RAECs were cultured in rat endothelial cell medium (RECM; Cell Applications) supplemented with 10% fetal bovine serum (FBS; GIBCO®, Invitrogen Corp., Carlsbad, CA, USA). RAECs were cultured in a 25-cm2 flask and then incubated at 37°C in a 5% CO2 incubator up to 95% confluence. RAECs at passage 7 were used in all experiments.
Measurement of cell cytotoxicity by WST-8 assay
Cytotoxicity against RAECs by apple procyanidin was conducted with a WST-8 assay. In brief, RAECs (5×103 cells per well) were cultured on 96-well plates with Dulbecco's modified Eagle's medium (DMEM) containing 10% FBS and then incubated at 37°C in a 5% CO2 incubator up to 95% confluence. The DMEM was changed to FBS-free medium for 24 h. RAECs were incubated with procyanidin C1 for 24 h in a volume of 100 μL. After the incubation, 10 μL of WST-8 solution (Dojindo Laboratories) was added to each well and incubated for 4 h. The absorbance was measured at 450 nm using a Wallac model 1420 microplate reader (Perkin Elmer Lifescience, Tokyo, Japan).
Measurement of membrane potential using bis-(1,3-dibutylbarbituric acid) trimethine oxonol
Changes in membrane potential were analyzed by a membrane potential-sensitive probe, bis-(1,3-dibutylbarbituric acid) trimethine oxonol [DiBAC4(3)] (Dojindo Laboratories). In brief, RAECs (1.5×104 cells per well) were cultured on a 96-well plate with DMEM containing 10% FBS and then incubated at 37°C in a 5% CO2 incubator up to 95% confluence. RAECs were washed twice with assay buffer (20 mM HEPES, 120 mM NaCl, 2 mM KCl, 2 mM CaCl2, 1 mM MgCl2·H2O, and 5 mM glucose) and were then incubated in assay buffer containing DiBAC4 (3) (0.5 μM) for 30 min with or without the nonspecific K+ channel inhibitor TEA (1 mM) at 37°C in the dark. After these pretreatments, procyanidin C1 (50 μM) was added to each well. Fluorescence was excited at 485 nm, and the emission was detected at 520 nm at intervals of 60 s using the Wallac model 1420 microplate reader. The first action point before change of the membrane potential was considered as 100%, and changes of DiBAC4(3) fluorescence intensity were set in relation to this measurement.
Measurement of [Ca2+]i by confocal microscopy
RAECs (2×105 cells per dish) were cultured on a 35-mm-diameter dish with 10% FBS in DMEM up to 95% confluence, and the medium was changed to serum-free medium 12 h prior to the measurement. Thereafter, RAECs were incubated in physiological saline solution buffer with 0.5% dimethyl sulfoxide and 0.02% Cremophor® EL (BASF Corp., Ludwigshafen, Germany) containing 5 μM Fluo-4 acetoxymethyl ester at 37°C for 60 min and rinsed twice with physiological saline solution buffer. RAECs were pretreated with EGTA (0.1 mM) for 10 min, and then procyanidin C1 (50 μM) was added to each well for 10 min. After stimulation, RAECs were visualized by a Nikon (Tokyo) confocal microscope with an attached camera.
Measurement of NO production by Griess reagent
The concentration of NO in culture supernatant was determined as nitrite, by the Griess reagent (1% sulfanilamide/0.1% naphthylethylenediamine dihydrochloride in 2.5% H3PO4) (Sigma-Aldrich Japan K.K., Tokyo). RAECs (1.5×104 cells per well) were cultured in 96-well plates with DMEM containing 10% FBS and then incubated at 37°C in a 5% CO2 incubator up to 95% confluence. RAECs were pretreated with the NO synthase (NOS) inhibitor
Statistical analysis
Results are expressed as mean±SEM values, where n refers to the RAEC preparations used in each experiment. The statistical difference between two groups was analyzed by the Tukey–Kramer's t test. P<.05 was considered to be statistically significant. All analyses were conducted with Stat View J5.0 software (SAS Institute Inc., Cary, NC, USA).
Results
Cytotoxic effect of procyanidin C1
To determine the adequate concentration of procyanidin C1 for all experiments, the cytotoxic effect of procyanidin C1 was primarily examined in RAECs. In a 5-day incubation of RAECs with procyanidin C1 (10–500 μM), procyanidin C1 (≥100 μM) significantly inhibited 10% FBS-induced cell proliferation, whereas the cytotoxic effect was not observed at concentrations of ≤50 μM (Fig. 2). For this reason, 50 μM procyanidin C1 was used as the maximum dose throughout subsequent experiments.
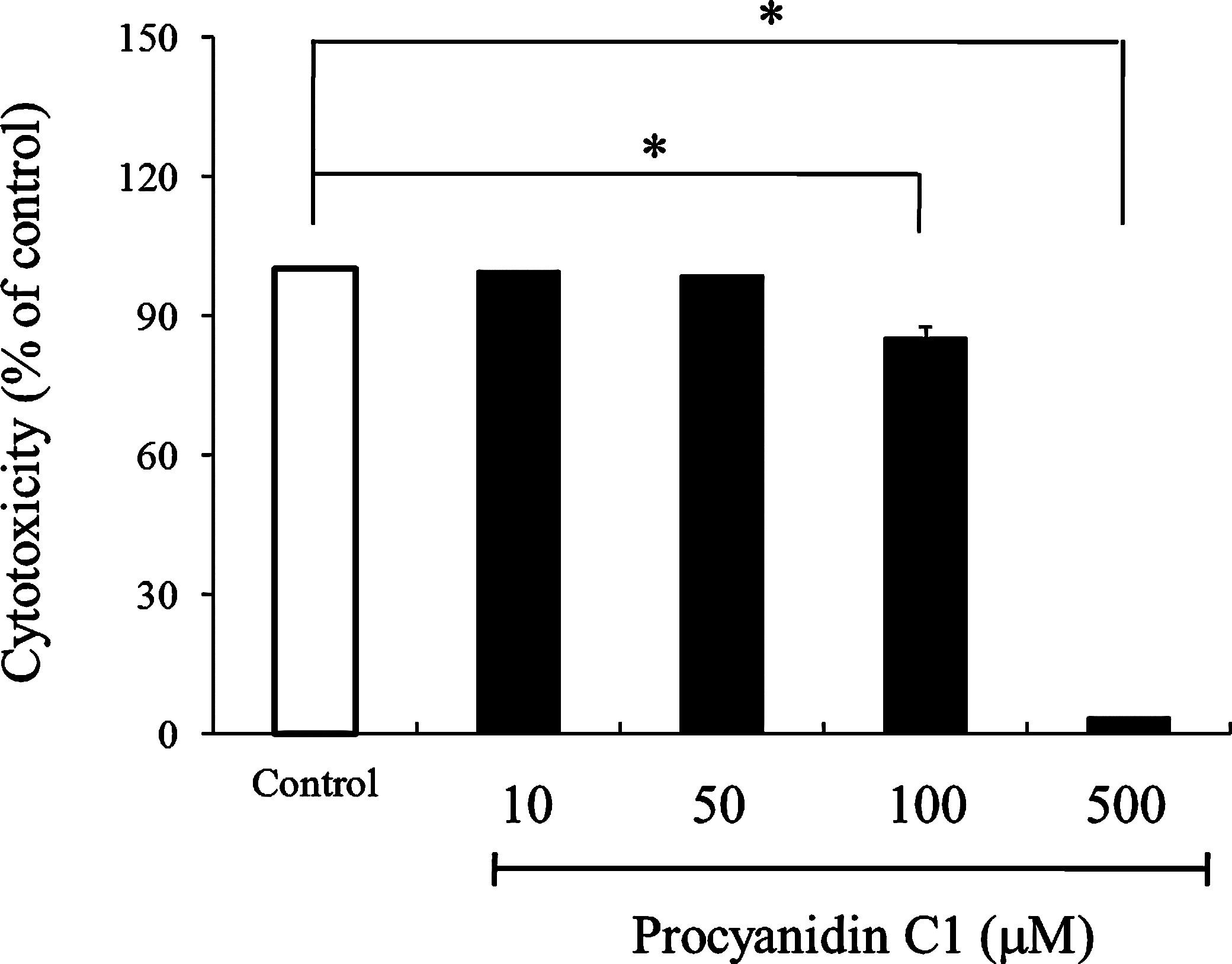
Cytotoxicity of procyanidin C1 at concentrations of 10 and 500 μM in rat aortic endothelial cells. Data are mean±SEM values (n=6). Significant differences between control and procyanidin groups were evaluated by the Tukey–Kramer t test: *P<.01 versus the control group.
Change of membrane potential by procyanidin C1
Procyanidin C1-induced hyperpolarization was examined with the membrane potential-sensitive probe DiBAC4(3) at a concentration of 50 μM. As shown in Figure 3, the addition of procyanidin C1 to RAECs resulted in a time- and dose-dependent decrease in DiBAC4(3) fluorescence, whereas the effect was significantly inhibited by the nonspecific K+ channel inhibitor TEA (1 mM), confirming that the procyanidin C1-induced hyperpolarization is mediated by activation of the endothelial K+ channel.
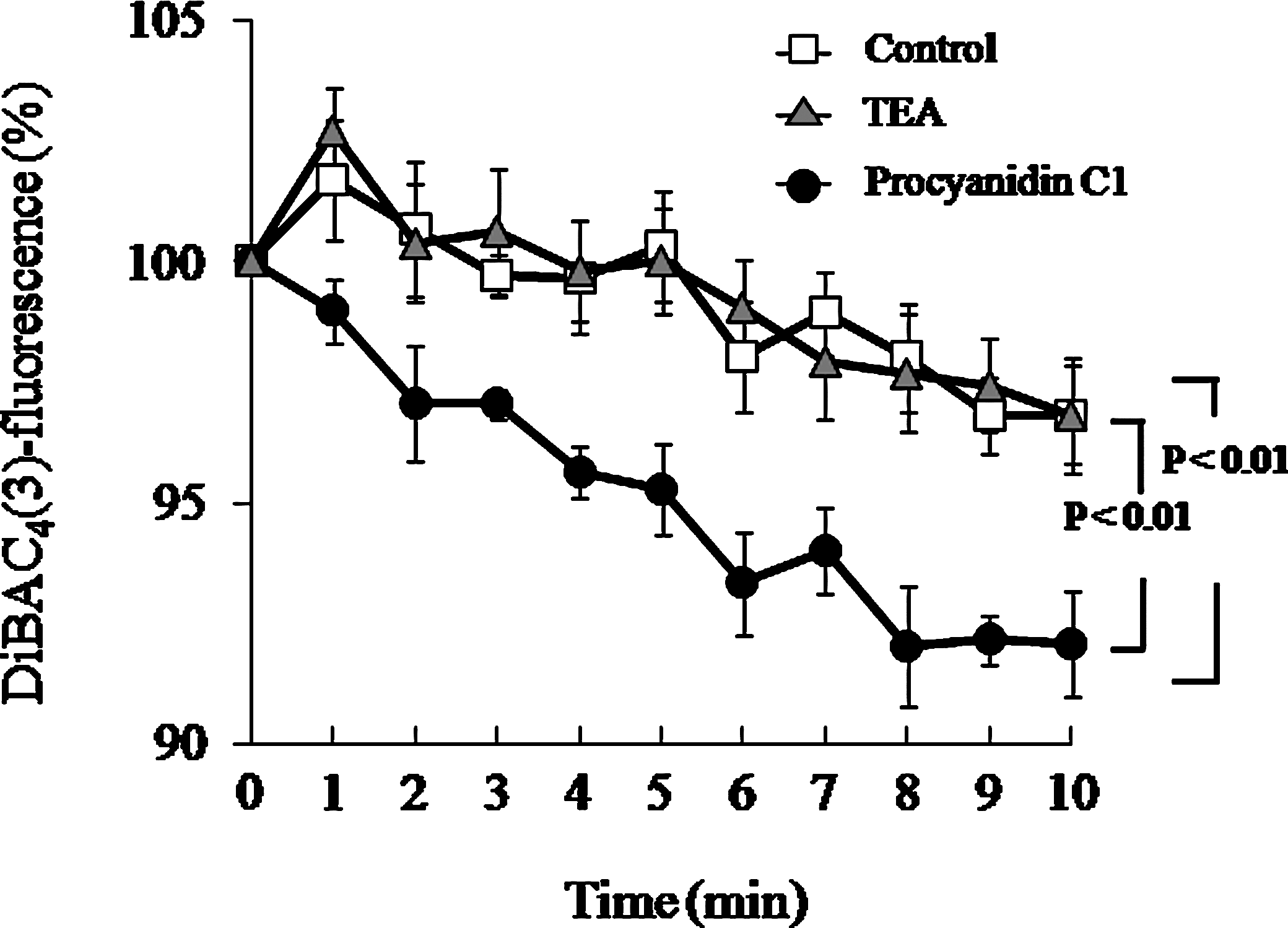
Change in procyanidin C1 (50 μM)-induced hyperpolarization in rat aortic endothelial cells by pretreatment with the nonspecific K+ channel inhibitor tetraethylammonium chloride (TEA) (1 mM). Data are mean±SEM values (n=4). Significant differences between values in the absence and the presence of inhibitor were evaluated by two-way analysis of variance. DiBAC4(3), bis-(1,3-dibutylbarbituric acid) trimethine oxonol.
Effect of procyanidin C1 on [Ca2+]i enhancement
As demonstrated by De Wit and Wölfle, 15 the elevation of Ca2+ could serve as a trigger for hyperpolarization via an activation of K+ channels. Accordingly, an experiment using Fluo-4 acetoxymethyl ester determined if procyanidin C1 increases [Ca2+]i concentrations. As shown in Figure 4, procyanidin C1 (50 μM) induced a significant elevation of [Ca2+]i in RAECs within 5 min following application, which was significantly abolished in the presence of Ca2+-free solution with EGTA (0.1 mM). These observations suggest that an elevation of [Ca2+]i by procyanidin C1 is involved in the signaling mechanisms promoting extracellular Ca2+ influx into RAECs.
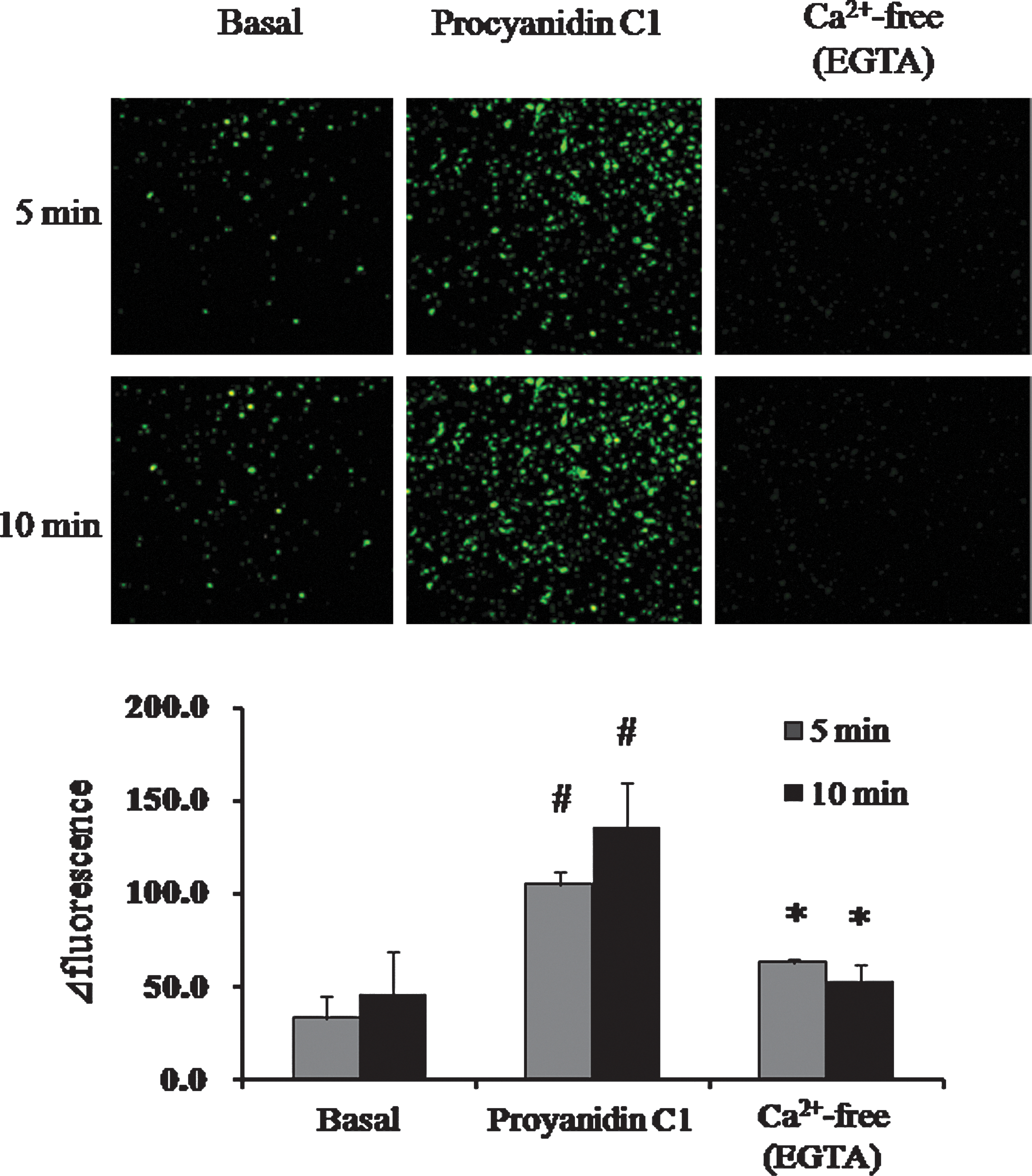
Effect of procyanidin C1 (50 μM) on intracellular Ca2+ concentration in rat aortic endothelial cells in the absence or presence of Ca2+-free solution with EGTA (0.1 mM). Data are mean±SEM values (n=3). #
P<.01 versus basal; *P<.05 versus the procyanidin C1 group by the Tukey–Kramer t test. Color images available online at
NO production induced by procyanidin C1
To define the role of K+ channel activity in procyanidin C1-induced NO production, the change of NO level in RAECs was tested using the Griess reagent. As shown in Figure 5, exposure of RAECs to procyanidin C1 increased NO production by fourfold compared with the control group. The enhanced NO production was significantly abolished in the presence of either the nonselective K+ channel inhibitor TEA (1 mM) or the NOS inhibitor
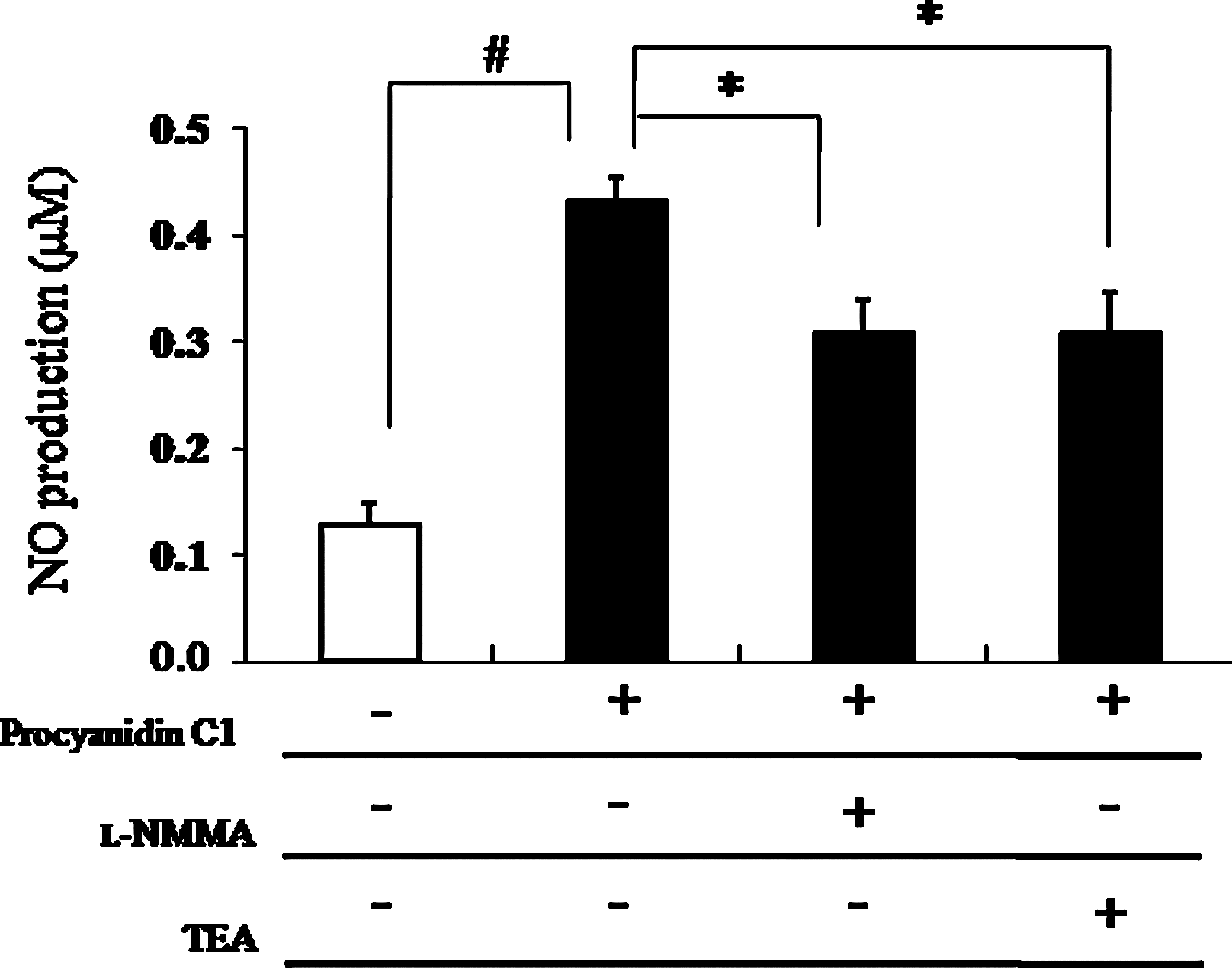
Effect of procyanidin C1 (50 μM)-induced nitric oxide (NO) production in rat aortic endothelial cells by pretreatment with the NO synthase inhibitor N
G-monomethyl-
Discussion
Polyphenolic compounds represent a wide variety of compounds that occur in fruits, vegetables, wine, tea, extra virgin olive oil, chocolate, and other cocoa products and that have well-characterized medicinal and preventive efficacy for the cardiovascular system.
4,16
In particular, those beneficial effects are also correlated with an improved vascular endothelium condition, indicating a great importance in the maintenance of vascular tone.
17,18
Regulation of vasorelaxation by vascular endothelium is achieved by several pathways, with the most important being endothelium-derived vasodilation mediated by NO synthesis through the action of
In order to establish such relationships, the cytotoxic effect was first evaluated. Procyanidin C1 resulted in a significant cytotoxicity at a concentration of ≥100 μM; for this reason, procyanidin C1 (50 μM) was used as the maximum dose throughout subsequent experiments. Treatment of RAECs with procyanidin C1 resulted in a time- and dose-dependent hyperpolarization. The effect was abolished by pretreatment with the nonspecific K+ channel inhibitor TEA, which raised the possibility that procyanidin C1-induced hyperpolarization is involved in the activation of K+ channels in RAECs.
In view of these findings, our working group was also able to establish in endothelial cells that K+ ion released from the endothelium may function as a vasorelaxing factor (i.e., EDHF) as previously reported.
22
It is important that our recent studies, as well as those conducted by other laboratories,
23
have linked hyperpolarization via K+ channel activation to endothelium-dependent vasorelaxation. In addition, there have been several reports showing that one of the most important regulatory and vasoactive substances produced by endothelial cells is NO,
24
and the endothelial NO pathway has been shown to be dependent on binding of calmodulin, thereby requiring elevation of both extracellular and intracellular Ca2+ concentrations.
25
In agreement with the above reports, procyanidin C1 promoted the activity of NO production, whereas the activity was abolished by pretreatment with either the nonselective K+ channel inhibitor TEA or the NOS inhibitor
Moreover, procyanidin C1 induced a significant elevation of [Ca2+]i in RAECs, which was significantly abolished in Ca2+-free solution with EGTA. This raises the possibility that the elevation of [Ca2+]i could serve as a trigger for hyperpolarization via an activation of K+ channels as reported by De Wit and Wölfle. 15 These findings are also consistent with previous observation from other investigators that quercetin-induced induction of the NO/cyclic GMP pathway depends on membrane hyperpolarization by Ca2+-mediated K+ channel activation. 26
Therefore, our working group has provided evidence for direct effects of procyanidin C1 on RAECs that could, in part, explain the clinical observations that polyphenol-rich compounds improve endothelial dysfunction associated with vascular disease. Taken together, we demonstrated for the first time that procyanidin C1 plays a potent role in an enhanced NO level via the elevation of [Ca2+]i concentration as well as membrane hyperpolarization via activation of the K+ channel in RAECs (Fig. 6), which could support a novel therapeutic mechanism for endothelial hyperpolarization as a key regulator of vascular tone.
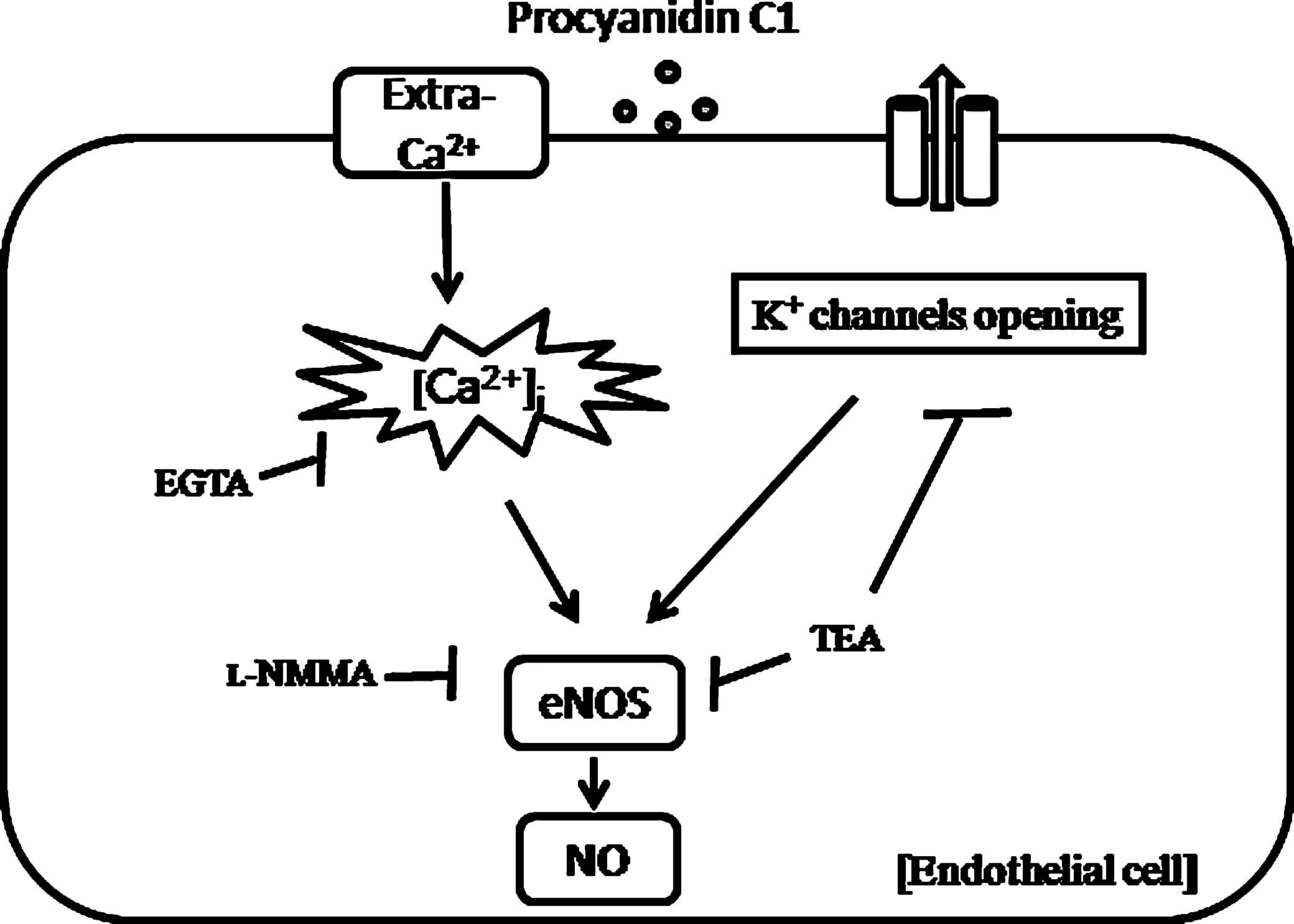
Proposed mechanism of procyanidin C1-induced vasorelaxation in rat aortic endothelial cells. [Ca2+]i, intracellular Ca2+; eNOS, endothelial NO synthase; Extra-Ca2+, extracellular Ca2+.
Footnotes
Author Disclosure Statement
None of the authors has financial or proprietary interest in any of the materials mentioned.