
Abstract
Bamboo salt is a traditional food widely used in Korea. The in vitro anticancer effects of this salt were evaluated in HCT-116 human colon cancer cells using a 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide (MTT) assay. A 1% salt concentration of bamboo salt baked nine times (9×) inhibited the growth of HCT-116 cells by 53%, which was higher than salt baked three times (3×) or once (1×; 44% and 41%, respectively) and much higher than solar sea salt (Korean sea salt) and purified salt (22% and 18%, respectively). To elucidate the inhibitory mechanisms underlying the anticancer effect of the salt samples in cancer cells, expression of genes associated with apoptosis, inflammation, and metastasis was measured with reverse transcription–polymerase chain reaction and Western blotting. Bamboo salt (9×) significantly induced apoptosis in cancer cells (P<.05) by upregulating Bax, caspase-9, and caspase-3, and downregulating Bcl-2. The expression of genes associated with inflammation (NF-κB, iNOS, and COX-2) was significantly downregulated (P<.05) by 9× bamboo salt, demonstrating its anti-inflammatory properties. The 9× bamboo salt also exerted a greater anti-metastatic effect on cancer cells than the other salts as demonstrated by decreased mRNA expression of MMP genes and increased expression of tissue inhibitors of metalloproteinases, which was confirmed by the inhibition of tumor metastasis induced in colon 26-M3.1 cells in BALB/c mice. In contrast, purified and solar salts increased metastasis in the mice. Our results demonstrated that 9× bamboo salt had the most potent in vitro anticancer effect, induced apoptosis, had anti-inflammatory activities, and exerted in vivo anti-metastatic effects. Additionally, the anticancer, anti-inflammatory, and anti-metastatic effects of the 1× and 3× bamboo salts were stronger than those of the purified and solar salts.
Introduction
B
Apoptosis induction in cancer cells is initially identified by morphological changes, including cell shrinkage, membrane blebbing, chromatin condensation, and nuclear fragmentation. 9 Apoptosis is an important defense against cancer. This process involves the elimination of potentially harmful cells. Many diseases have been associated with dysregulated apoptotic processes, ultimately leading to the inhibition of cell death and propagation of diseases, such as cancer. Elucidating the critical events associated with carcinogenesis provides an opportunity for preventing cancer development via dietary intervention by inducing apoptosis, particularly with bioactive agents or functional foods. Diet is a significant environmental factor in the overall cancer process, and can exacerbate or interfere with disease progression. In addition to dietary effects on protein expression and function, evidence is also accumulating that a large number of food components can exert effects on the human genome, either directly or indirectly modulating gene expression. 10,11
Caspases are central components of the apoptotic response. Caspase-9 is an apical isoform involved in mitochondria-dependent apoptosis. This factor primarily activates caspase-3, which then serves as a gateway for the activation of downstream caspases. 12 Nuclear factor-κB (NF-κB) is involved in the inhibition of apoptosis, stimulation of cell proliferation, inflammation, immune response, and tumorigenesis. Activation of NF-κB generally prevents apoptosis. Expression of inducible nitric oxide synthase (iNOS) and cyclo-oxygenase-2 (COX-2), two genes regulated by NF-κB, is induced by inflammation, and these genes are frequently overexpressed in cancer cells. Increased NF-κB activity localized in the nucleus is particularly found in cells where there is abundant expression of iNOS and COX-2. 13
A previous epidemiological study showed that chronic inflammation predisposes individuals to various types of cancer. 14 Hallmarks of inflammation-related cancers include the presence of inflammatory cells and mediators in tumor tissues, tissue remodeling and angiogenesis similar to that seen during chronic inflammatory responses, and tissue repair. The study of mechanisms underlying inflammation-related cancer has focused on the early stages of cancer, but inflammatory mediators and cells are also involved in the migration, invasion, and metastasis of malignant cells. 15 Metastasis is the leading cause of death among cancer patients, and involves the spread of cancer from a primary site and formation of new tumors in distant organs. Matrix metalloproteases (MMPs) play important roles in many physiological or pathological processes, including embryonic development, morphogenesis, reproduction, tissue remodeling, arthritis, cardiovascular disease, and metastasis. 16 MMP activity is inhibited by specific endogenous tissue inhibitors of metalloproteinases (TIMPs). 17 To prevent most types of cancer, improved treatments for metastasis are needed. 18,19
Previously, we found that bamboo salt produces stronger in vitro anticancer effects than solar or purified salts in HT-29 colon cancer cells 20 and HepG2 liver cancer cells (unpublished data). In the present study, we further examined the anticancer and anti-metastatic effects of bamboo salts. Bamboo salt was administered to HCT-116 human colon cancer cells and the molecular mechanisms underlying the anticancer effects of the salt were studied. We evaluated changes in activities of bamboo salts baked for different periods of time and also assessed their anti-metastatic effects in mice with tumors propagated by 26-M3.1 colon carcinoma cells.
Materials and Methods
Preparation of bamboo salts
Bamboo salts (baked 1, 3, and 9 times) and solar salt were provided by Taesung Food Company (Gochang, Jeonbuk, South Korea). Purified salt manufactured by Hanju Corporation (Ulsan, South Korea) was purchased in a local market (Busan, South Korea). The salts were dissolved in sterilized distilled water to concentration of 20% before conducting the experiments.
Cancer cell preparation
HCT-116 human colon carcinoma cells obtained from the American Type Culture Collection (Manassas, VA, USA) were used for the experiments. The cells were cultured in RPMI-1640 medium (Welgene, Inc., Daegu, South Korea) supplemented with 10% fetal bovine serum (FBS; Gibco Co., Birmingham, MI, USA) and 1% penicillin-streptomycin (Gibco) at 37°C in a humidified atmosphere containing 5% CO2 (model 311 S/N29035; Forma, Waltham, MA, USA). The medium was changed two or three times each week.
3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide assay
Anticancer effects of the bamboo salt were assessed by 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide (MTT) assay. HCT-116 human colon carcinoma cells were seeded in a 96-well plate at a density of 2×104 cells/mL in a volume 180 μL per well. Salt solutions with concentrations of 0.5% and 1.0% (20 μL) were added and then the cells were incubated at 37°C in 5% CO2 for 48 h. An MTT solution (200 μL, 5 mg/mL; Amresco, Solon, OH, USA) was added and the cells were cultured for another 4 h under the same conditions. After removing the supernatant, 150 μL of dimethyl sulfoxide was added per well and mixed for 30 min. Finally, the absorbance of each well was measured with an enzyme-linked immunosorbent assay reader (model 680; Bio-Rad, Hercules, CA, USA) at 540 nm. 21
Flow cytometry analysis
After treatment with salt samples, the cells were trypsinized, collected, washed with cold phosphate-buffered saline (PBS), and resuspended in 2 mL PBS. DNA contents of the cells were measured using a DNA staining kit (CycleTEST™ PLUS kit; Becton Dickinson, Franklin Lakes, NJ, USA). Nuclear fractions stained with propidium iodide were obtained by following the manufacturer's protocol. Fluorescence intensity was determined using an FACScan flow cytometer (EPICS XL-MCL; Beckman Coulter KK, Brea, CA, USA) and analyzed with CellQuest software (Becton Dickinson). 22
Reverse transcription–polymerase chain reaction to measure mRNA expression
Total RNA from HCT-116 human colon carcinoma cells was isolated using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's recommendations. The RNA was digested with RNase-free DNase (Roche, Basel, Switzerland) for 15 min at 37°C and purified using an RNeasy kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. cDNA was synthesized from 2 μg of total RNA by incubation at 37°C for l h with avian myeloblastosis virus reverse transcriptase (GE Healthcare, Little Chalfont, United Kingdom) with random hexanucleotides according to the manufacturer's instruction. Sequences of primers used to specifically amplify the genes of interest are shown in Table 1. Amplification was performed in a thermal cycler (Eppendorf, Hamburg, Germany). The polymerase chain reaction (PCR) products were separated in 1.0% agarose gels and visualized with ethidium bromide staining. 23
COX-2, cyclo-oxygenase-2; iNOS, inducible nitric oxide synthase; MMP, matrix metalloprotease; NF- κ B, nuclear factor-κB; TIMP, tissue inhibitors of metalloproteinases; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Protein extraction and Western blot analysis
Total cell lysates were obtained with an extraction buffer as previously described. 24 Protein concentrations were determined using a protein assay kit (Bio-Rad, Hercules, CA, USA). For Western blot analysis, the cell lysates were separated by 12% SDS-PAGE, transferred onto a polyvinylidene fluoride membrane (GE Healthcare), blocked with 5% skim milk, and incubated with the primary antibodies (1:1000 dilution). Antibodies against Bax, Bcl-2, iNOS, and COX-2 were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). After incubation with the horseradish peroxidase-conjugated secondary antibody at room temperature, immunoreactive proteins were detected using a chemiluminescent enhanced chemiluminescence assay kit (GE Healthcare) according to the manufacturer's instructions. Bands in the blot were visualized using a LAS3000 luminescent image analyzer (Fujifilm Life Science, Tokyo, Japan).
Measuring lung metastasis after salt treatment in BALB/c mice bearing 26-M3.1 colon carcinoma cell tumors
26-M3.1 colon carcinoma cells were obtained from Prof. Yoon, Department of Food and Nutrition, Yuhan University, Bucheon, South Korea. These highly metastatic cells were maintained as monolayers in EMEM (Welgene, Inc., Daegu, South Korea) supplemented with 7.5% FBS, a vitamin solution, sodium pyruvate, nonessential amino acids, and L-glutamine (Gibco). The cultures were maintained in a humidified atmosphere of 5% CO2 at 37°C. Experimental lung metastasis was induced by injecting colon 26-M3.1 cells into the lateral tail vein of 6-week-old female Balb/c mice (Hyochang Science, Daegu, South Korea). 25 Different salt solutions (400 and 800 mg/kg) were subcutaneously injected into the mice, and the animals were then intravenously inoculated with 26 M-3.1 cells (2.5×104/mouse) after 2 days. After 2 weeks the mice were sacrificed and their lungs were fixed in Bouin's solution (saturated picric acid:formalin:acetic acid; 15:5:1, v/v/v). The rate of metastasis was assessed by counting lung tumor colonies using a digital camera (Canon D550; Tokyo, Japan). The protocol for these experiments was approved by the Animal Ethics Committee of Pusan National University.
Statistical analysis
Data are presented as the mean±standard deviation. Differences between the mean values for individual groups were assessed with a one-way analysis of variance with Duncan's multiple-range test. Differences were considered significant when P<.05. SAS version 9.1 (SAS Institute, Inc., Cary, NC, USA) was used for statistical analyses.
Results
In vitro anticancer effect of bamboo salt on HCT-116 cells
Anticancer effects of purified, solar, and bamboo (1×) salts on HCT-116 cells were evaluated using an MTT assay. At concentrations ranging from 0.1% to 5.0%, cell viability was decreased by the salt samples in a concentration-dependent manner. At a concentration of 5%, the survival rate of the cells treated with bamboo salt reached 0%. Consequently, concentrations of 0.5% and 1% were selected for subsequent experiments (Fig. 1A). HaCaT keratinocyte normal cells were also evaluated, and dose–response rates were similar to those of HCT-116 cells. Most of the cells treated with three different salts survived at 1.0% (Fig. 1B). The growth inhibitory rates of HCT-116 human colon carcinoma cells treated with the different salt samples are shown in Table 2. When solutions of the salt samples (0.5%) were administered to HCT-116 cells, the growth inhibitory rates observed with purified, solar, 1× bamboo, 3× bamboo, and 9× bamboo salts were 6%, 8%, 16%, 21%, and 29%, respectively. At a concentration of 1%, the growth inhibitory rate associated with 9× bamboo salt was 53%, higher than that of the 3× and 1× bamboo salts (44% and 41%, respectively). On the other hand, treatment with solar and purified salts resulted in relatively low inhibitory rates of 22% and 18%, respectively (P<.05). These results demonstrated that bamboo salt had greater antiproliferative effects on HCT-116 cells compared with purified and solar salts. In addition, the more times the bamboo salt was baked, the stronger the anticancer effects. In addition, DNA content of the sub-G1 cancer cells was evaluated by flow cytometric analysis. The induction of apoptosis was almost negligible (3.30%) in control cancer cells; however, cells treated with 9×bamboo salt had a higher level of apoptosis (29.6%) than ones treated with the other salts (Fig. 2).
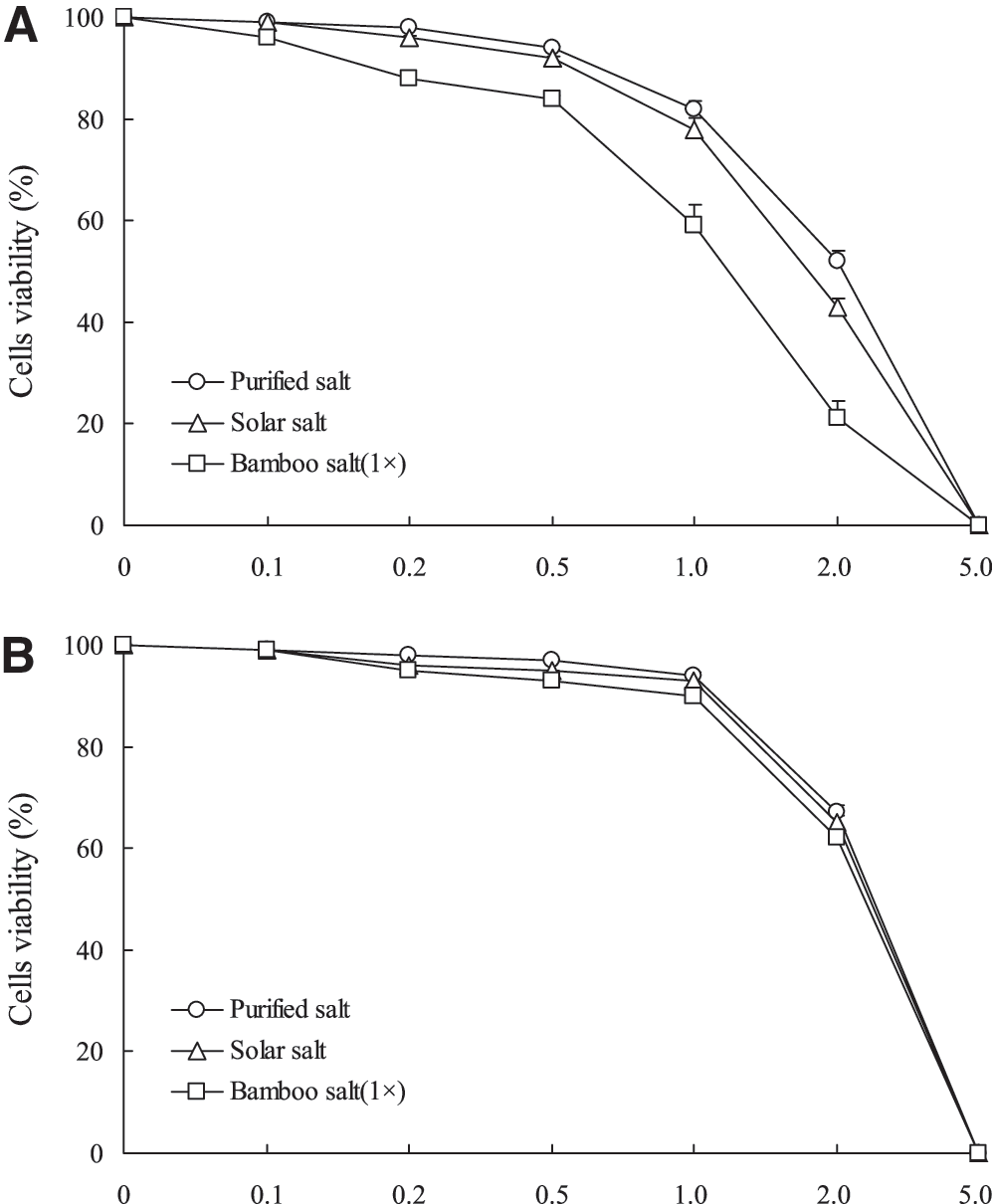
Effect of three different kinds of salt on the growth of HCT-116 human colon carcinoma cells
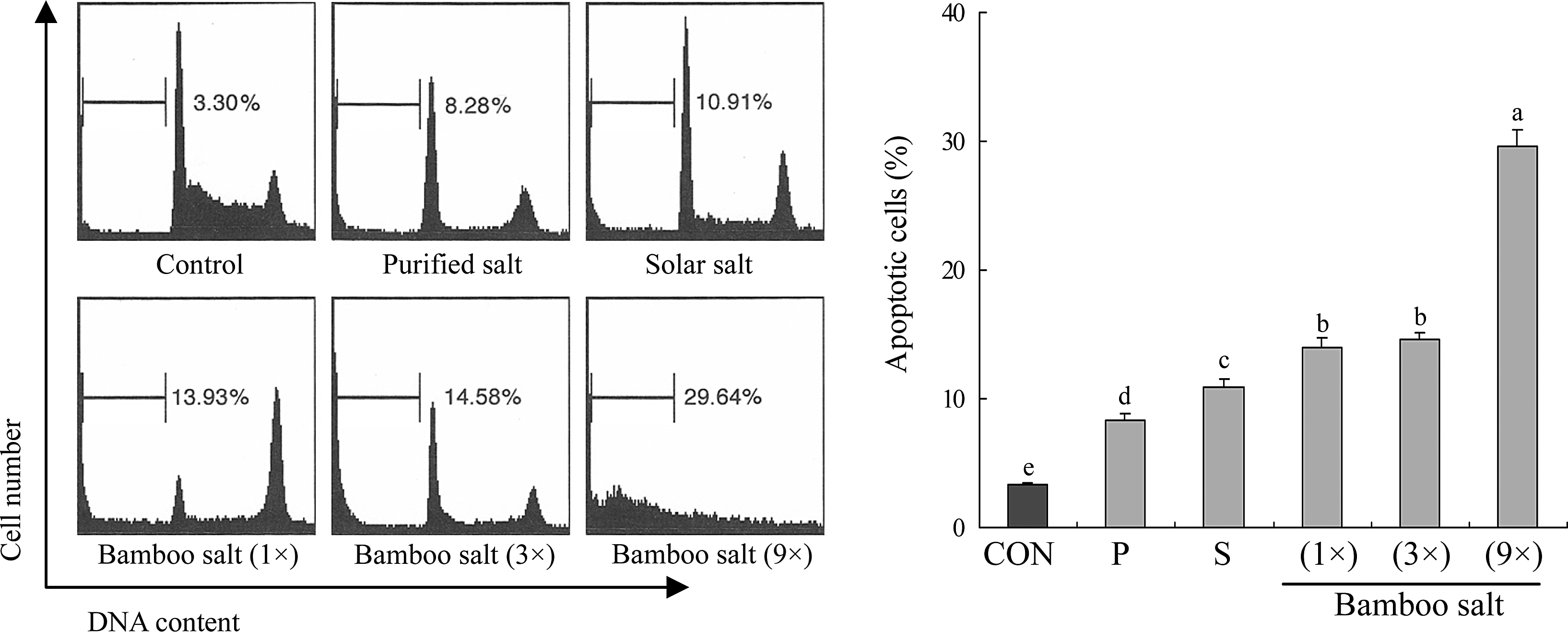
The level of apoptosis induced by various salts. The DNA content of sub-G1 HCT-116 human colon carcinoma cells was evaluated using a flow cytometer. Apoptosis was monitored by staining with annexin-V FITC. a–fMean values with different letters over the bars are significantly different (P<.05) according to Duncan's multiple-range test. CON, control; P, purified salt; S, solar salt.
The values in parentheses are the inhibition rates (%).
Mean values with different letters in the same column are significantly different (P<.05) according to Duncan's multiple-range test.
MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide.
Apoptosis-related gene expression of Bax, Bcl-2, and caspases
To elucidate the mechanisms underlying the inhibition of cancer cell growth by the bamboo salt samples, expression of Bax, Bcl-2, and caspase-9 and -3 in HCT-116 human colon carcinoma cells was measured by reverse transcription (RT)–polymerase chain reaction (PCR) and Western blot analysis after a 48-h incubation with 1% salt solutions. As shown in Figure 3, expression of pro-apoptotic Bax and anti-apoptotic Bcl-2 showed significant changes (P<.05) in the presence of 9× bamboo salt. These results suggest that the salt induced apoptosis in the HCT-116 cells via a Bax- and Bcl-2–dependent pathway. The mRNA expression levels of caspase-9 and -3 were very low in untreated control HCT-116 cells, but significantly increased after the cells were treated with 1% of bamboo salt (9×). With the bamboo salt treatment, mRNA expression of caspase-9 and -3 was gradually elevated with increased baking times (Fig. 4). More specifically, apoptosis induction by the bamboo salt (9×) was related to upregulation of Bax, caspase-9, and caspase-3, and downregulation of Bcl-2 in terms of mRNA and protein expression. The effects of this salt were greater compared with those of the 1× and 3× bamboo salt samples as well as the solar and purified salt samples.
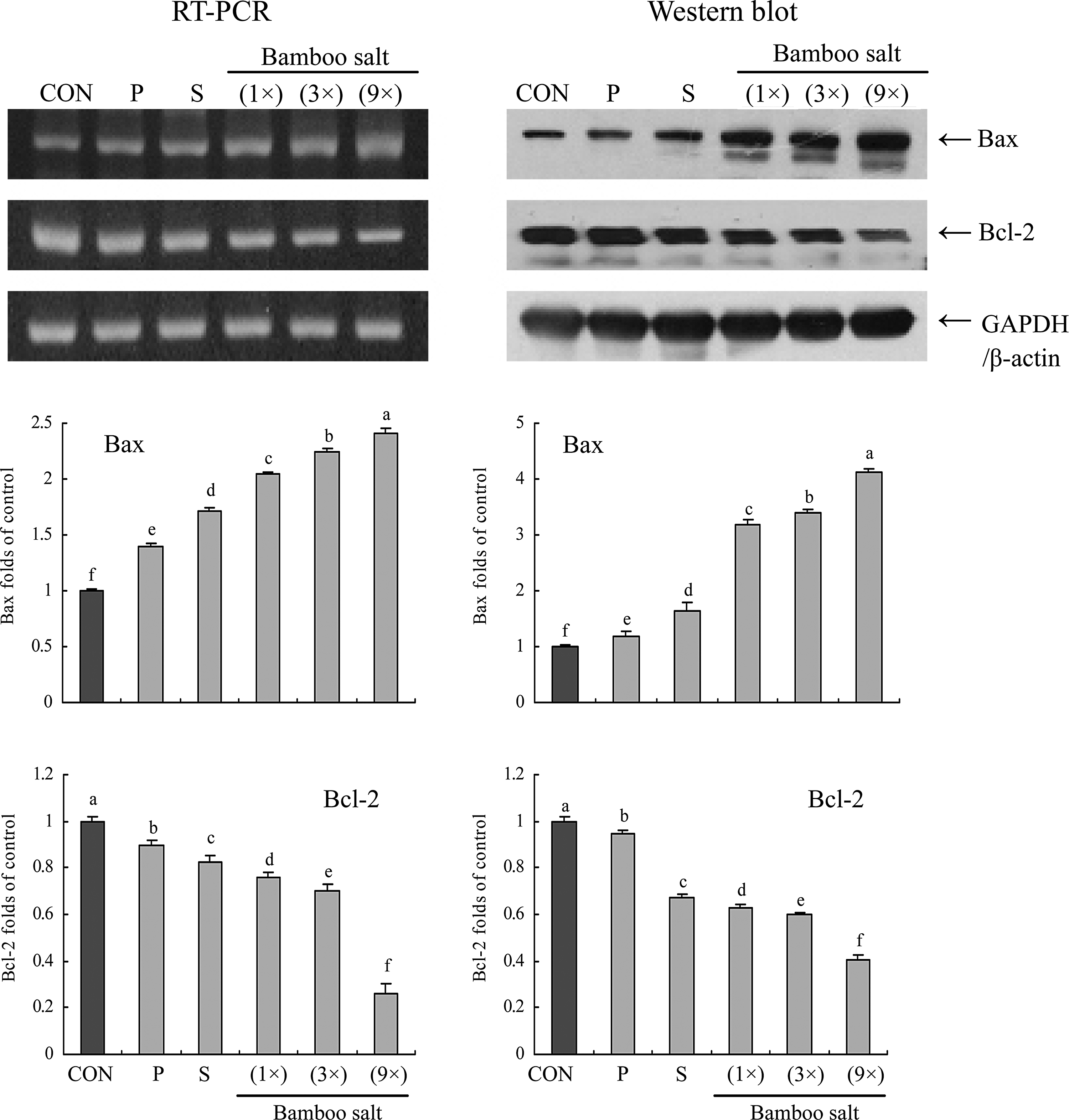
Effects of salt samples (1% solution) on the mRNA and protein expression of Bax and Bcl-2 in HCT-116 human colon carcinoma cells. Band intensity was measured with a densitometer and expressed as fold of the control. Fold ratio=gene expression/glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (β-actin)×control numerical value (control fold ratio: 1). a–fMean values with different letters over the bars are significantly different (P<.05) according to Duncan's multiple-range test. CON, control; P, purified salt; S, solar salt.
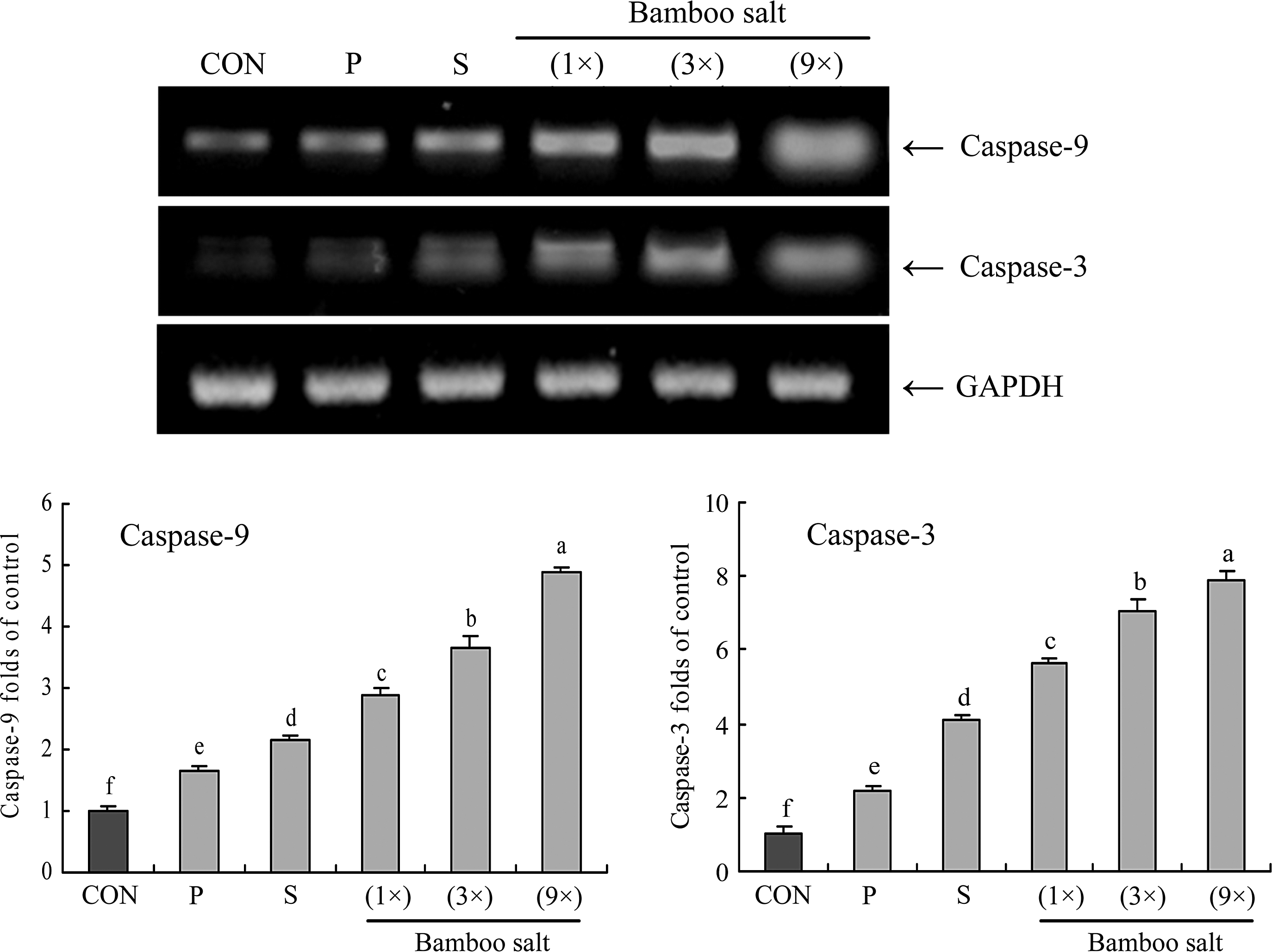
Effects of salt samples (1% solution) on the mRNA expression of caspase-9 and -3 in HCT-116 human colon carcinoma cells. Band intensity was measured with a densitometer and expressed as fold of the control. Fold ratio=gene expression/GAPDH×control numerical value (control fold ratio: 1). a–fMean values with different letters over the bars are significantly different (P<.05) according to Duncan's multiple-range test. CON, control; P, purified salt; S, solar salt.
Inflammation-related gene expression of NF-κB, IκB-α, iNOS, and COX-2
We determined whether the anticancer actions of bamboo salt were associated with inhibition of NF-κB-p65, IκB-α, iNOS, and COX-2 gene expression. As shown in Figure 5, mRNA expression of NF-κB-p65 and IκB-α was reduced in HCT-116 cells treated with a 1% bamboo salt (9×) solution. Bamboo salt (9×) significantly modulated the expression of genes associated with inflammation (P<.05). mRNA expression of NF-κB-p65 was decreased while IκB-α mRNA levels were increased. Additionally, mRNA and protein expression of COX-2 and iNOS was gradually decreased in the presence of the bamboo salt depending on the baking times (Fig. 6). Our findings indicate that 9× bamboo salt may help prevent cancer in the early stages by increasing anti-inflammatory activities. Overall, the results of this experiment showed that bamboo salts had a stronger anti-inflammatory effect on the colon cancer cells than solar and purified salts.

Effects of salt samples (1% solution) on the mRNA expression of nuclear factor-κB (NF-κB)-p65 and IκB-α in HCT-116 human colon carcinoma cells. Band intensity was measured with a densitometer and expressed as fold of the control. Fold ratio=gene expression/GAPDH×control numerical value (control fold ratio: 1). a–fMean values with different letters over the bars are significantly different (P<.05) according to Duncan's multiple-range test. CON, control; P, purified salt; S, solar salt.
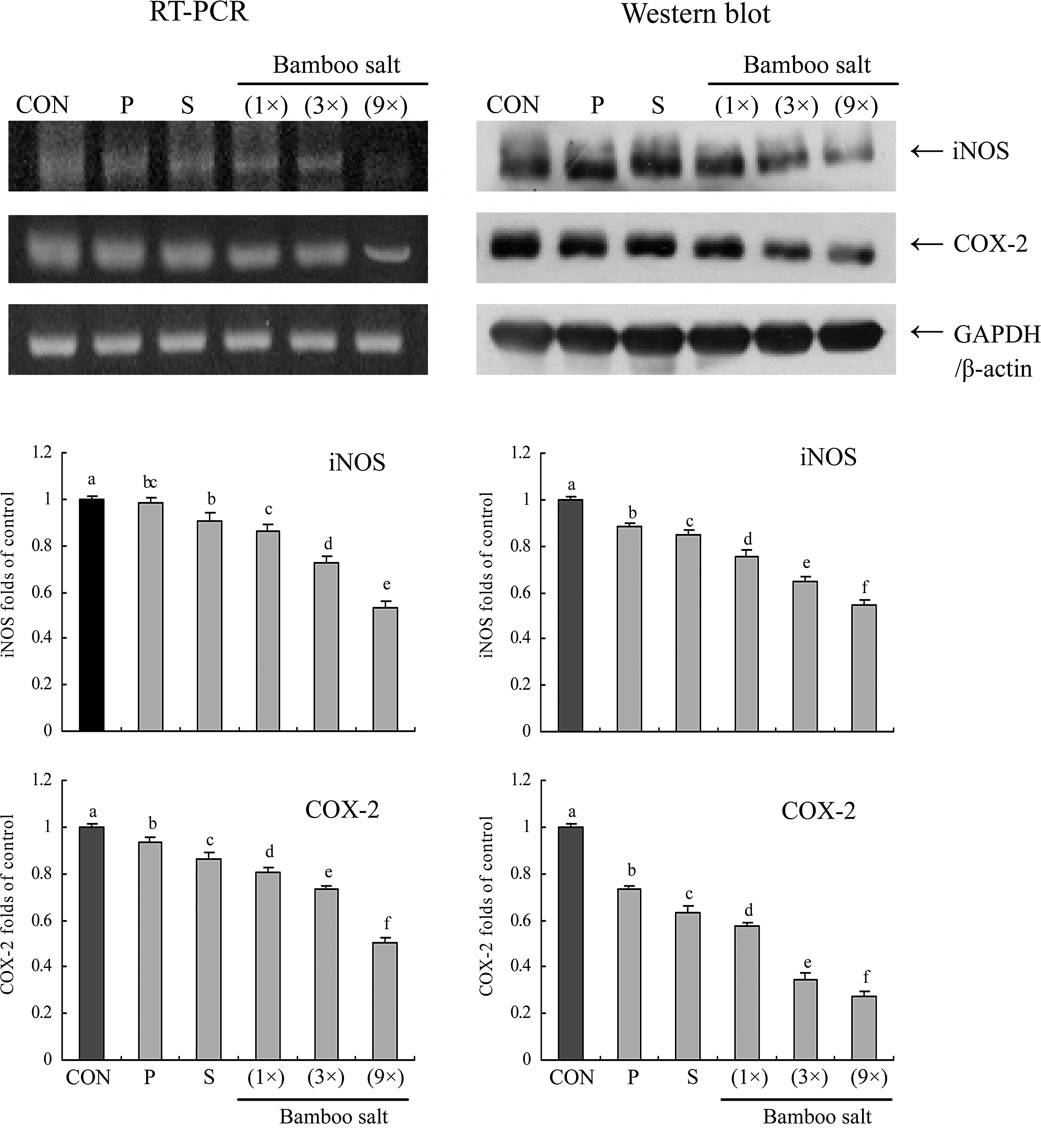
Effects of salt samples (1% solution) on the mRNA and protein expression of inducible nitric oxide synthase (iNOS) and cyclo-oxygenase-2 (COX-2) in HCT-116 human colon carcinoma cells. Band intensity was measured with a densitometer and expressed as fold of the control. Fold ratio=gene expression/GAPDH (β-actin)×control numerical value (control fold ratio: 1). a–fMean values with different letters over the bars are significantly different (P<.05) according to Duncan's multiple-range test. CON, control; P, purified salt; S, solar salt.
Metastasis-related MMP and TIMP gene expression
RT-PCR analyses were conducted to determine whether the anti-metastatic effect of the bamboo salts was due to gene regulation of metastatic mediators, specifically MMPs (MMP-2 and MMP-9) and TIMPs (TIMP-1 and TIMP-2), in HCT-116 cells. As shown in Figure 7, 9× bamboo salt significantly decreased mRNA expression of MMP-2 and MMP-9, and increased the expression of TIMP-1 and TIMP-2. These changes in TIMP and MMP expression resulting from bamboo salt treatment could effectively lead to metastatic inhibition in vitro. Our results also showed that 9×bamboo salt had stronger anti-metastatic activity than the other salts.
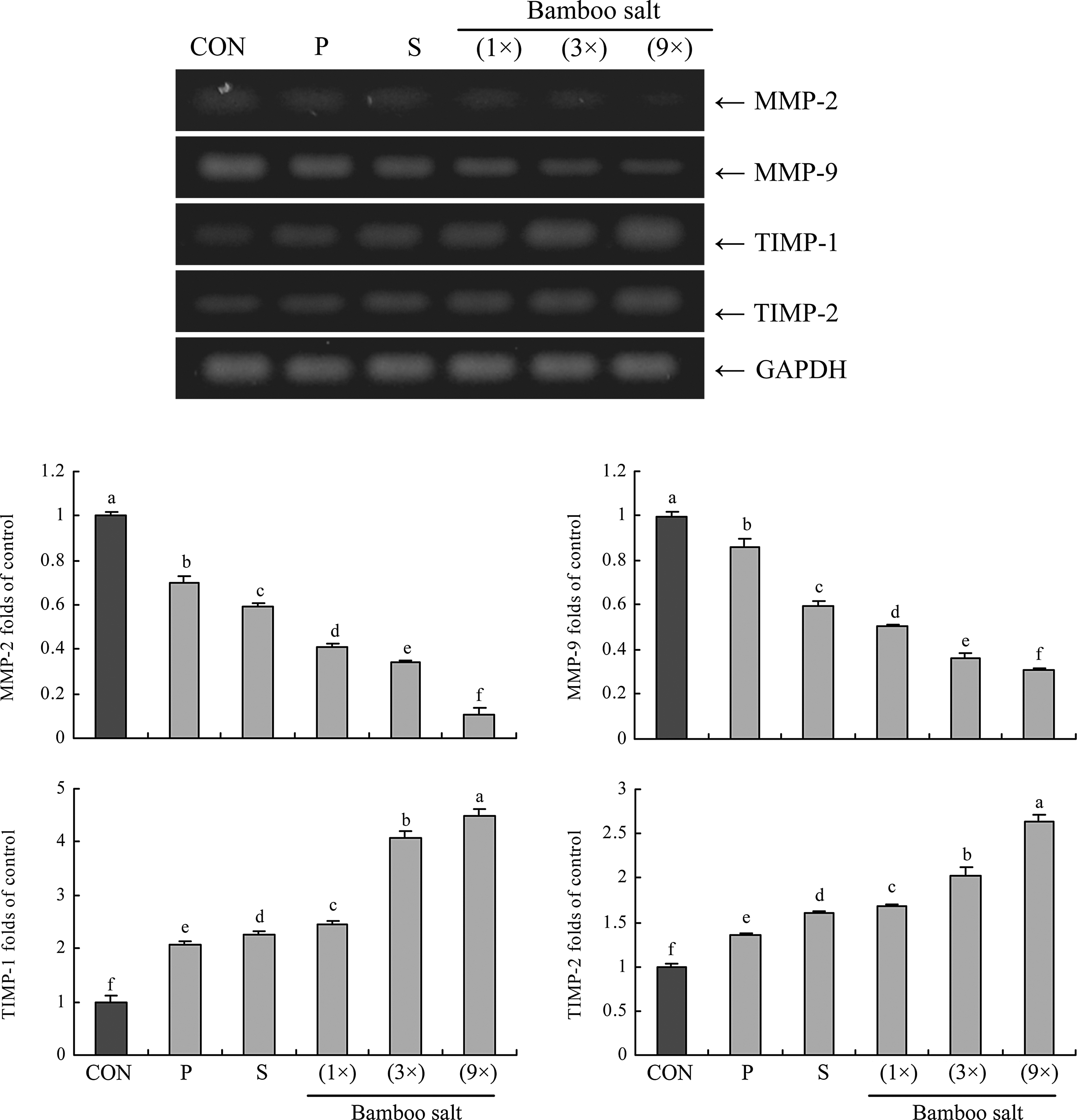
Effects of salt samples (1% solution) on the mRNA expression of matrix metalloproteases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs) in HCT-116 human colon carcinoma cells. Band intensity was measured with a densitometer and expressed as fold of the control. Fold ratio=gene expression/GAPDH×control numerical value (control fold ratio: 1). a–fMean values with different letters over the bars are significantly different (P<.05) according to Duncan's multiple-range test. CON, control; P, purified salt; S, solar salt.
In vivo anti-metastatic effect of the bamboo salts
Prophylactic inhibition of tumor metastasis by the bamboo salts was evaluated by using an experimental mouse metastasis model (Fig. 8). All bamboo-salt-treated mice had significantly fewer lung metastatic colonies than those treated with solar salt, purified salt, or control mice (number of metastatic tumors=50±3, n=10; P<.05). The 9×bamboo salt was the most effective for inhibiting lung metastasis at a concentration of 400 mg/kg. This bamboo salt (inhibitory rate=16%; number of metastatic tumors=42±2) inhibited tumor formation and lung metastasis to a greater degree than the 3× (inhibitory rate=8%; number of metastatic tumors=46±2) and 1× bamboo salts (inhibitory rate=4%; number of metastatic tumors=48±3). Solar (inhibitory rate=−2%; number of metastatic tumors=51±4) and purified salts (inhibitory rate=−4%; number of metastatic tumors=52±3) actually increased metastasis. At 800 mg/kg, the inhibition rates associated with 9×, 3×, and 1× bamboo salts along with solar and purified salts were 20%, 16%, 6%, −10%, and −14%, respectively (number of metastatic tumors was 40±2, 42±3, 47±3, 55±3, and 57±3, respectively).
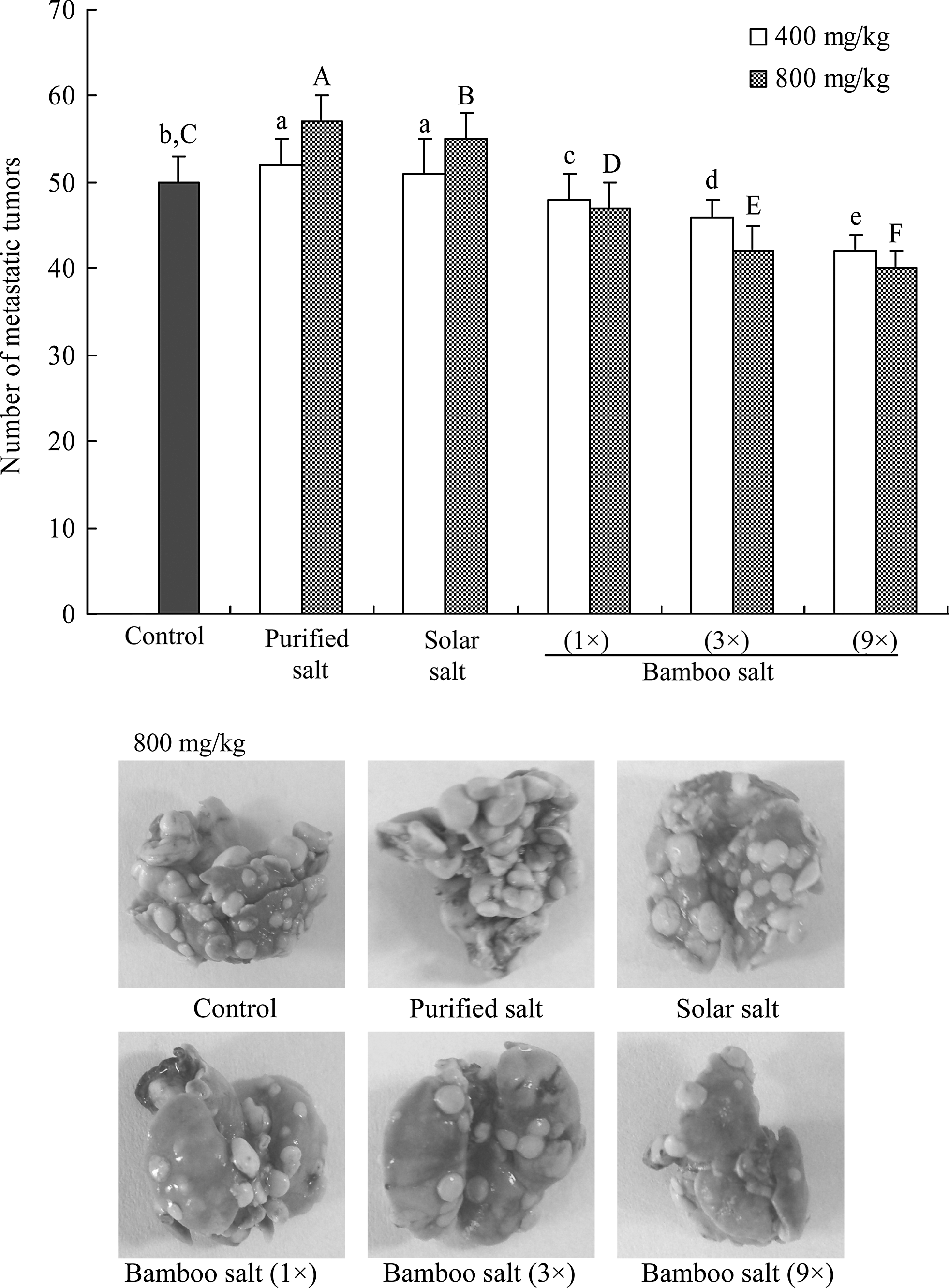
Inhibitory effect of salt samples on the metastasis of tumors produced by colon 26-M3.1 cells in Balb/c mice. The mice were administered with the indicated dose of salt by subcutaneous injection and then inoculated intravenously with 26-M3.1 cells (2.5×104 cells/mouse) after 2 days. Mice were sacrificed 14 days after tumor inoculation and tumor colonies in the lungs were counted. A–F,a–eMean values with different letters on the bars are significantly different (P<.05) according to Duncan's multiple-range test.
Discussion
Although bamboo salt has been used as a folk medicine, little scientific data on its effects are available. The 9× bamboo salt, also called purple bamboo salt since the color changes to purple after the baking process, has high concentrations of iron, silicon, and potassium compared with crude salt. 18 This salt has been recently reported to have various therapeutic effects on numerous pathologic conditions, such as inflammation, viral diseases, diabetes, and cancer. 26,27
Apoptosis is a fundamental cellular event, and understanding its mechanisms of action will help harness this process for use in tumor diagnosis and therapy. 28 In a healthy cell, the anti-apoptotic protein Bcl-2 is expressed on the outer mitochondrial membrane surface. 29 Because the Bax and Bcl-2 genes are mainly expressed during apoptosis, we determined that these genes regulate apoptotic activity. Apoptosis results from activation of caspase family members that act as aspartate-specific proteases. 30 Caspases form a proteolytic network within the cell whereby upstream initiator caspases are activated early in the apoptotic process (caspase-9) and in turn activate other downstream caspases (caspase-3). Cytochrome c and procaspase-9 processing is highly dependent on caspase-3, placing this caspase in a central position as a regulator of essential apoptotic pathways in cancer cells. 31 Caspase-3 was also reported to play a role as an amplifier of apoptotic signals (i.e., by cleaving Bcl-2). 32
Additionally, anticancer mechanisms underlying the effect of bamboo salt on human cancer cells involve the induction of apoptosis by increasing the number of apoptotic bodies, regulating the mRNA and protein expression of Bax and Bcl-2, and promoting anti-inflammatory effects by downregulating iNOS and COX-2 gene expression. COX-2 has been suggested to play an important role in colon carcinogenesis, and NOS, along with iNOS, may be a good target for the chemoprevention of colon cancer. 33 NF-κB is one of the most ubiquitous transcription factors, and regulates the expression of genes required for cellular proliferation, inflammatory responses, and cell adhesion. 34 NF-κB is present in the cytosol where it is bound to the inhibitory protein IκB. Following its induction by a variety of agents, NF-κB is released from IκB and translocates to the nucleus where it binds to the κB binding sites in the promoter regions of target genes. 35 These mechanisms could be involved in the anticancer effects of bamboo salt in colon cancer cells. Based on the results of the MTT assay and the expression patterns of proapoptotic genes observed in the present study, we concluded that cancer cells treated with bamboo salt underwent apoptosis. Similar to our findings, the anticancer effects of bamboo salt in HT-29 colon cancer cells were evaluated in a previous study by an MTT assay and RT-PCR or Western blotting analysis, and bamboo salt showed stronger anticancer effects than common salt (sea salt). 20
Metastasis is defined as the spread of cancer cells from one organ or area to another adjacent organ or location. 36,37 It is thought that malignant tumor cells have the capacity to metastasize. Cancer occurs after cells in a tissue are genetically damaged in a progressive manner, resulting in cancer stem cells possessing a malignant phenotype. After the tumor cells come to rest in another site, they penetrate the vessel walls, continue to multiply, and eventually form another tumor.
MMPs, a family of zinc-dependent endopeptidases, play a very important role in tumorigenesis and metastasis. MMPs can cleave virtually all extracellular matrix (ECM) substrates. Degradation of the ECM is a key event in tumor progression, invasion, and metastasis. 38 Among the MMP family members, MMP-2 and MMP-9 are molecules important for cancer invasion, 39,40 and highly expressed in breast and colon cancer cells. 41 –43 In fact, inhibition of MMP activity is useful for controlling tumorigenesis and metastasis. 44 TIMPs are naturally occurring inhibitors of MMPs, which prevent catalytic activity by binding to activated MMPs, thereby blocking ECM breakdown. 45 Disturbances in the ratio between MMPs and TIMPs have been observed during tumorigenesis. 46 Maintaining the balance between MMPs and TIMPs or increasing TIMP activity is a useful way to control tumor metastasis. 47 Experimental evidence demonstrating the role of MMPs in metastasis has been obtained by in vitro invasion assays and in vivo xenograft metastasis experiments.
MMP-2 and -9 are key factors in cancer cell invasion and metastasis both in vivo and in vitro. 48 Spontaneous and experimental metastasis to the liver is decreased in mice overexpressing TIMP-1, and increased in mice expressing antisense TIMP-1 mRNA. 49 Ectopic overexpression of TIMP-1 in the brain of transgenic mice also reduces experimental metastasis to the brain. 50 In particular, MMP-2 and -9 are important for tumor invasion and angiogenesis. Thus, tumor metastasis can be inhibited by blocking MMP synthesis and activity. 51 Colon 26-M3.1 carcinoma cells have been used to evaluate anti-metastatic effects in vivo. 52 Studies of salt-fermented foods like kimchi and doenjang have used this method. 53,54 The anti-metastatic effects of solvent fractions from doenjang fermented with bamboo salt indicated that this type of salt can increase the anti-metastatic activities of doenjang compared with that made with solar salt.
In the current study, we chose suitable concentrations of bamboo salt to perform our experiments. Concentrations of bamboo salt below 0.5% were not associated with apoptotic activity while concentrations above 1.0% led to high rates of growth inhibition. This inhibitory activity appears to be accomplished via apoptosis rather than necrosis. At a 1.0% concentration, the bamboo salt exerted anticancer and anti-metastatic effects on HCT-116 cells. All salt samples, including solar and purified salts, were found to have in vitro anti-metastatic effects based on the RT-PCR analysis of MMP and TIMP gene expression. However, solar and purified salts showed a prometastatic effect whereas bamboo salt had an anti-metastatic effect in vivo. These results demonstrated that bamboo salt had anti-metastatic effects in vitro and in vivo. In contrast, the solar and purified salts had only a modest anti-metastatic effect in vitro and actually promoted metastasis in vivo. More research is needed to explain the mechanisms associated with these effects.
The potassium, calcium, magnesium, and iron contents in bamboo salts are higher than those in purified and solar salts. Additionally, bamboo salt baked for longer periods of time contains more other minerals. 55 Increased levels of these minerals in the salt are important for enhancing the anticancer effects. 56 Bamboo salt also exhibits a higher reduction potential; this might be due to the fact that this type of salt contains more OH groups than purified or solar salt. 57 Overall, bamboo salt seems to have more potent anticancer and anti-inflammatory effects because it contains higher levels of minerals and more OH groups.
In summary, we used various in vitro experimental methods, including MTT, RT-PCR, and Western blot assays, to evaluate the anticancer effects of bamboo salts. We also used a mouse model bearing tumors produced by 26-M3.1 colon carcinoma cells to study the in vivo effects of these salts. Results from the present study demonstrated that the anti-metastatic effects of bamboo salts were stronger than the ones produced by solar or purified salts. Overall, bamboo salts showed more potent in vitro and in vivo anticancer activities than the other types of salts, especially for combating in vivo tumor metastasis. In contrast, purified and solar salts actually promoted metastasis. Increased mineral and phytochemical contents due to the processing of bamboo salt are important for augmenting these anticancer effects. Baking the bamboo salt nine times increased the anticancer properties of the salt in this study. The active compounds resulting from baking bamboo salt need to be identified and evaluated in future studies.
Footnotes
Acknowledgment
This research was supported by the National Academy of Agricultural Science, Rural Development Administration, South Korea.
Author Disclosure Statement
No competing financial interests exist.