
Abstract
Blue-green algae (BGA) are among the most primitive life forms on earth and have been consumed as food or medicine by humans for centuries. BGA contain various bioactive components, such as phycocyanin, carotenoids, γ-linolenic acid, fibers, and plant sterols, which can promote optimal health in humans. Studies have demonstrated that several BGA species or their active components have plasma total cholesterol and triglyceride-lowering properties due to their modulation of intestinal cholesterol absorption and hepatic lipogenic gene expression. BGA can also reduce inflammation by inhibiting the nuclear factor κ B activity, consequently reducing the production of proinflammatory cytokines. Furthermore, BGA inhibit lipid peroxidation and have free radical scavenging activity, which can be beneficial for the protection against oxidative stress. The aforementioned effects of BGA can contribute to the prevention of metabolic and inflammatory diseases. This review provides an overview of the current knowledge of the health-promoting functions of BGA against cardiovascular disease and nonalcoholic fatty liver disease, which are major health threats in the developed countries.
Introduction
B
BGA are nutritious natural products rich in essential amino acids, γ-linolenic acid (GLA), fibers, B vitamins, calcium, phosphorous, iron, pigments such as β-carotene, xanthophylls, and chlorophyll, and other bioactive compounds. 2,6,10 Studies have indicated that BGA have antiviral, antitumor, antioxidant, anti-inflammatory, antiallergic, antidiabetic, and antibacterial properties as well as lipid-lowering effects. 11 In particular, inhibitory effects of BGA on hyperlipidemia, inflammation, and oxidative stress can contribute to the prevention of the development of CVD and nonalcoholic fatty liver disease (NAFLD). This review presents the current understanding of the protective effects of BGA against metabolic and inflammatory diseases.
Discussion
Athero-protective effects of BGA
CVD is the primary cause of death in developed countries. 12 Dyslipidemia characterized by high concentrations of plasma total cholesterol, triglycerides and low-density lipoprotein cholesterol (LDL-C), and low plasma high-density lipoprotein cholesterol (HDL-C) levels is a primary risk factor for CVD. 13 Statins have been used to decrease plasma total cholesterol and LDL-C levels in patients with hypercholesterolemia. Statin therapy, however, is often accompanied by undesirable side effects, such as liver dysfunction or myalgia. 14,15 Therefore, natural hypocholesterolemic products are a desirable alternative. Water-soluble fibers, omega-3 fatty acids, plant sterols, and soy have been used to prevent dyslipidemia. 16 –18 However, identification and development of new evidence-based natural products with a potent cholesterol and triglyceride-lowering effect are necessary to satisfy health-conscious consumers' need for attaining optimal health.
Lipid-lowering effect of BGA
Since the first report on a hypocholesterolemic effect of BGA in the early 1980s, several studies have shown potential health-benefits of various BGA species in the prevention of CVD. 19 Supplementation of Spirulina species with up to 15% by weight of diet significantly improved plasma lipid profiles of hyperlipidemic rats and mice fed a high-fat/high-cholesterol diet or a high-fructose diet. 20 –22 In rats with carbon tetrachloride (CCl4)-induced fatty liver, hepatic total cholesterol and triglyceride levels were significantly lower when rats were fed SM compared with control animals. 23 Aqueous extract of SM at 5% and 10% supplementation levels inhibited sucrose-induced increases in plasma concentrations of total cholesterol, LDL-C, and triglyceride, while it increased HDL-C concentrations in Wistar rats. 24 In addition to Spirulina species, 5% NO supplementation showed a hypolipidemic effect in male Wistar rats fed a high-cholesterol diet. 5 In our laboratory, we also reported that male C57BL/6J mice fed an AIN-93M diet supplemented with 5% of NO for 4 weeks showed significantly lower plasma total cholesterol and triglyceride levels. 4
In human clinical trials, supplementation with several species of Spirulina exhibited lipid-lowering effects. In patients with type 2 diabetes, subjects who consumed 2 g/day of SP for 2 months showed significantly lower plasma triglyceride concentrations as well as a significant reduction in ratios of total cholesterol:HDL-C and LDL-C:HDL-C. 2 Eight grams per day of SP supplementation for 12 weeks significantly reduced plasma triglyceride concentrations and blood pressure in type 2 diabetic patients with higher initial triglyceride levels, whereas subjects with high initial total cholesterol and LDL-C showed significant reductions in the plasma lipids. 25 In patients with a hyperlipidemic nephrotic syndrome, 1 g/day of SP supplementation for 2 months significantly decreased plasma total cholesterol, LDL-C, and triglyceride concentrations. 26
Anti-atherogenic effect of BGA
BGA with lipid-lowering effects are expected to have athero-protective properties. Indeed, Riss et al. 27 reported that fatty lesion development and cardiac production of superoxide anion were significantly decreased in hamsters fed an atherogenic diet supplemented with phycocyanin (PC), a compound abundantly present in SP, for 12 weeks. In New Zealand white rabbits fed a high-cholesterol diet containing 5% SP powder, SP-fed animals showed marked reductions in plasma concentrations of total cholesterol, triglycerides, and LDL-C as well as in atherosclerotic lesion development. 28
Potential mechanisms of action for the lipid-lowering effect of BGA
We evaluated potential mechanisms for the lipid lowering effect of NO in HepG2 cells, a human hepatoma cell line. NO lipid extract significantly downregulated the expression of 3-hydroxy-3-methyl-glutaryl-CoA reductase (HMGR), the rate-limiting enzyme in cholesterol biosynthesis, the LDL receptor (LDLR) and lipogenic genes, such as fatty acid synthase and stearoyl-CoA desaturase 1. 29 The repression was concomitant with decreased mature forms of sterol-regulatory element-binding protein 1 (SREBP-1) and SREBP-2, which are major transcriptional regulators for the above-mentioned genes. 30,31 We also found that the SP lipid extract elicited the same effect (manuscript in preparation). The observations are conflicting with in vivo findings in that mice fed 5% NO for 4 weeks showed an increased HMGR and LDLR expression in the liver. 4 Interestingly, the hypocholesterolemic effects of NO were attributed to decreased intestinal cholesterol absorption with a concomitant increase in the fecal neutral sterol output. 4 Therefore, the in vivo lipid-lowering effect of NO is likely due to its bioactive components in the delipidated fraction, but not in the lipid fraction of NO. To test this possibility, we fed C57BL/6J mice an AIN-93M diet supplemented with 5% NO or its lipid extract equivalent to 5% of NO. The study showed that only mice-fed NO, but not NO lipid extract, had lower plasma total cholesterol and triglyceride levels (manuscript in preparation). NO contains high amount of fibers, which may be a major component for eliciting the lipid-lowering effect. Hori et al. 5 have shown that fiber components of NO play a critical role in lowering plasma cholesterol in Wistar rats fed a high-cholesterol diet supplemented with different fiber components highly present in NO. The study demonstrated that rats fed a neutral detergent fiber and oxalic acid soluble substance had a significantly lower serum cholesterol levels. Therefore, fibers are likely to play a major role in lowering plasma cholesterol levels. It is not known what components in BGA lipid extracts have strong inhibitory effects on SREBP maturation as well as cholesterol biosynthesis. As BGA contain small amounts of lipids, the contribution of bioactive components in the fraction to whole-body lipid metabolism may be minor when whole algae are consumed. However, given the potency of the repression as shown in our in vitro studies, identification of the bioactive components has a great therapeutic significance for developing a drug to lower blood lipids.
Up to now, BGA dosages used in human clinical trials have ranged from 1 to 8 g/day for up to 24 weeks, whereas ∼5 g/day is generally recommended by the manufacturers. 32 Although individual cases of discomfort or unpleasant condition have been reported with BGA supplementation from time to time, Dietary Supplements Information Expert Committee (DSI-EC) of the United States Pharmacopeial Convention has awarded Spirulina (SM and SP) a grade A safety rating and agreed that Spirulina is generally safe to be consumed. 32 Concern over using BGA during pregnancy and breast feeding is not clearly defined in humans. However, numerous animal studies indicated that SP consumption during pregnancy and lactation did not induce signs of maternal intoxication. 33 In contrast, Kapoor and Mehta 34 had demonstrated that SP supplementation could improve iron status during pregnancy, as evidenced by a higher hemoglobin count, serum iron, and serum ferritin. Further study is necessary to evaluate the effect of BGA supplementation in humans during specific nutrition status and pathological conditions.
Anti-inflammatory effects of BGA
Atherosclerosis is not only the passive accumulation of lipids within artery walls, but is an inflammatory disease. In the initial stage of atherosclerosis development, the normal functions of the endothelium are altered and an inflammatory response is promoted, which facilitates recruitment of circulating monocytes. 35,36 This, in turn, predisposes the vessel wall to lipid accumulation. Inflammatory proteins, such as C-reactive protein, interleukin-6 (IL-6), serum amyloid A, and tumor necrosis factor α (TNFα), are suggested as inflammatory markers for CVD to provide a clinical means for identifying patients at high risk for CVD. 37 The nuclear factor kappa B (NF-κB) is a dimeric transcription factor that regulates the production of inflammatory cytokines, for example, TNFα, IL-1β, and monocyte chemoattractant protein (MCP-1) in response to proinflammatory agents, including TNFα, IL-1, and a bacterial endotoxin lipopolysaccharide (LPS). 38 –41 Studies have demonstrated that NF-κB is activated in established human atherosclerotic plaques and many inflammatory genes in the region are regulated by NF-κB. 42 –45
Anti-inflammatory property of BGA
Consumption of BGA has been demonstrated to promote immunity and to protect against inflammatory diseases, such as colitis, arthritis, and allergic rhinitis in animal and human studies. 46 –49 In particular, oral administration of 800 mg SF per day for 8 days to adjuvant-induced arthritic mice significantly inhibited arthritis markers to near normal condition. 49 Additionally, when collagen-induced arthritic rats were orally administered with 400 mg/kg of SP for 45 days, SP relieved the mice of several symptoms of arthritis, such as redness, swelling, deformity, and ankylosis in hind paws and ankle joints, suggesting that SP protects against arthritis. 50 In rats with colitis, oral gavage of 2 g/kg Spirulina for 7 days decreased inflammation in mucosa and submucosa and tissue malondialdehyde (MDA) levels compared with control animals, indicating Spirulina may be beneficial for preventing inflammatory bowel disease. 48
Our in vitro investigation strongly supports anti-inflammatory properties of BGA. In LPS-induced RAW 264.7 macrophages, NO lipid extract markedly reduced the mRNA expression of proinflammatory cytokines, such as TNFα and interleukins 1β (IL-1β), by inhibiting the NF-κB DNA binding activity. 51 Our recent study also demonstrated that NO and SP organic extracts markedly decreased the secretion of proinflammatory cytokines, including the granulocyte-macrophage colony stimulating factor, IL-6, MCP-1, and TNFα, in RAW 264.7 macrophages stimulated by LPS (manuscript in preparation). Moreover, translocation of NF-κB from cytoplasm to nucleus was also inhibited by both NO and SP extracts. We also observed that primary splenocytes isolated from apolipoprotein E knockout mice fed 5% NO and SP showed significantly lower IL-6 secretion when the cells were challenged by LPS compared with control splenocytes (manuscript in preparation).
Taken together, anti-inflammatory effects of BGA have been shown in cell studies, animal studies, and human studies. More study, however, is needed to determine if different BGA species have a different degree of efficiency in preventing inflammation. Furthermore, understanding of the effect of BGA on acute or chronic inflammatory pathways is necessary because although the two pathways share some of common mediators, protective effects against chronic inflammation at a low degree for a long period of time are more relevant and beneficial to the prevention of metabolic diseases such as CVD and NAFLD.
Anti-inflammatory BGA components
Several components of BGA, including GLA and PC, have been implicated in their anti-inflammatory effects. SP contains ∼1.3% GLA. GLA can be metabolized to dihomo-GLA that undergoes oxidative metabolism by cyclooxygenases and lipoxygenases to produce anti-inflammatory eicosanoids. 52 GLA is frequently used to treat rheumatoid arthritis, atopic eczema, acute respiratory distress syndrome, cancer, and asthma, and to improve plasma lipid profiles. 53 An anti-inflammatory effect of SP is also often associated with antioxidant and antiviral properties of C-PC and R-PC. 54 C-PC is a natural blue pigment accounting for ∼14% of SP dry weight. 19,47,55 C-PC in SP is a potent inhibitor of inflammation as evidenced by decreased paw edema index when inflammation was induced to mice fed C-PC by glucose oxidase. 56 In addition, a recent study suggests that C-PC suppresses the activation of NF-κB by preventing degradation of cytosolic inhibitor of κB α, consequently reducing the production of TNFα in LPS-stimulated RAW 264.7 macrophages. 57 In Kupffer cells, liver resident macrophages, C-PC abolished thyroid calorigenesis-induced TNFα levels, indicating its repressive effect on TNFα secretion triggered by oxidative stress. 58 C-PC also significantly reduced myeloperoxidase activity, inflammatory cell infiltration, and colonic damage in rats with acetic acid-induced colitis. 59
Antioxidant properties of BGA
Oxidative stress, mainly characterized by the formation of excessive reactive oxygen species (ROS), damages tissues and therefore is associated with several pathological conditions, such as atherosclerosis, nonalcoholic steatohepatitis (NASH), and aging. 60 Hydroxyl and peroxyl radicals are believed to be associated with oxidative damage because they are unstable and can react quickly with adjacent molecules to capture electrons. Free radical attacks can lead to chain production of radicals and eventually cause damage to cellular components such as lipids and DNA. 61 Therefore, administration of antioxidants can be beneficial for preventing ROS-related disorders. 62
Prevention of oxidative stress by BGA
Studies have reported the protective effects of BGA against oxidative damage in vivo and in vitro. Golden Syrian hamsters fed an atherogenic diet supplemented with SP or purified PC showed decreases in superoxide production and the expression of p22phox subunit of NADPH oxidase, indicating antioxidant properties of SP and PC in vivo. 27 Kim et al. 63 reported that in hypercholesterolemic New Zealand white rabbits, 5% SP supplementation for 8 weeks inhibited lipid peroxidation, while it markedly elevated hepatic glutathione (GSH) levels and activities of GSH peroxidase (GPx), GSH reductase, and GSH S-transferase (GST) compared with control animals. SP also significantly decreased thiobarbituric acid reactive substance (TBARS) values, an index of lipid peroxidation and oxidative stress, in plasma, liver, kidney, and heart, and reduced oxidative stress-induced DNA damage in lymphocytes. When mercuric chloride-intoxicated mice were given a diet supplemented with SF at a dose of 800 mg/kg for 30 days, the BGA supplementation restored hepatic GSH levels as well as activities of superoxide dismutase (SOD), catalase, and GST, while suppressing lipid peroxidation. 64 Ethanol extract of Spirulina laxissima abrogated the losses of antioxidant enzymes and decreased TBARS and liver function markers in the serum of CCl4-treated rats when the animals were orally treated with the extract three times a week for 5 weeks at 50 or 100 mg/kg. 65
Human studies have confirmed the BGA protection against oxidative stress. SP supplementation of 8 g/day for 12 weeks significantly decreased plasma levels of MDA, a biomarker of oxidative stress, in diabetic patients. 25 Similarly, healthy elderly Korean subjects who consumed 8 g/day of SP for 16 weeks showed a decrease in the lipid peroxidation level, whereas the total antioxidant status and levels of antioxidant enzymes, such as SOD and GPx, were elevated in plasma, indicating that SP supplementation was able to improve antioxidant status in the human subjects. 66
Antioxidant components in BGA
The protective effect of BGA against oxidative stress has been largely attributed to their high content of C-PC, a water soluble phycobiliprotein in BGA. 55 Phycobiliproteins are highly conserved chromoproteins that constitute phycobilisome, a protein complex important for photosynthesis in BGA. 67 It has been shown that C-PC markedly inhibited the production of alkoxyl radicals that are generated by the reaction of tert-butyl hydroperoxide with ferrous ions in the presence of luminol. 56 In addition, C-PC prevented DNA damage and scavenged hydroxyl and peroxylradicals. 56,68 Free radical and peroxyl radical scavenging abilities have also been reported in C-PC extracted from AFA. 69 Furthermore, the C-PC-rich extract from AFA inhibited peroxyl radical-induced oxidative hemolysis and lipid peroxidation in normal human erythrocytes. 70 The AFA extract demonstrated protective effects against cupric chloride-induced lipid oxidation in human plasma samples. Cysteine-rich cyanopeptide beta 2 isolated from C-PC of SF demonstrated free radical scavenging, inhibition of DNA damage, and decreased ROS production. 71 Selenium-containing allophycocyanin (Se-APC) extracted from selenium-enriched SP inhibited 2,2′-azobis-2-methyl-propanimidamide, dihydrochloride (AAPH)-induced oxidative hemolysis, and morphological changes in human erythrocytes. 72 Se-APC, furthermore, inhibited AAPH-induced intracellular ROS production and MDA accumulation.
In addition to the aforementioned radical scavenging effects, studies have shown an inhibitory effect of C-PC on lipid peroxidation. ROS generated during normal metabolic processes can easily initiate the peroxidation of membrane lipids, leading to the accumulation of lipid peroxides. 73 Moreover, membrane lipid peroxidation is responsible for the leakage of cytosolic enzymes into circulation during chemical-induced toxicity. 68 When lipid peroxidation was triggered by ferrous ascorbic acid in isolated rat liver microsomes as a model for cellular membranes, C-PC extracted from SP decreased TBARS, indicating its inhibitory role in lipid peroxidation. 56 Bhat and Madyastha 68 reported C-PC inhibited nearly 95% of peroxyl radical-induced lipid peroxidation. C-PC contains three types of chromophores: one binding to the α-subunit and two binding to the β-subunit of a C-PC monomer. 74 C-PC with a reduced chromophore can efficiently inhibit peroxyl radical-induced lipid peroxidation in a dose-dependent manner. 68 The in vitro evidence supports that C-PC has strong antioxidant properties by scavenging radicals and inhibiting lipid peroxidation in cell membranes.
Although PC has been suggested to be the major constituent of BGA with an antioxidant capacity, BGA also contain vitamin C, E, and B12, carotenoids, phenolic compounds, and minerals that can function as antioxidants. 63,75 Due to the presence of multiple antioxidant components, the administration of whole algae may result in synergistic antioxidant effects. The major components of BGA added as a functional ingredient in food products is a water extract, which contains C-PC and other water-soluble antioxidant substances. Chu et al. 76 compared antioxidant effects of SP water extract with purified PC. Treatment with SP water extract decreased free radical-induced apoptosis in 3T3 cells and the radical scavenging activity of the SP water extract was higher than that of purified PC, suggesting a mixture of antioxidant components in the extract may have stronger antioxidant effects than those of PC. In addition, diets enriched with SP increased cerebellar GSH concentrations and the activities of antioxidant enzymes, including SOD, catalase, and GPx, whereas lipid peroxide and amyloid β-protein deposition were decreased by 200 mg/kg of SP water extract supplementation in the brain of senescence-accelerated mice. 77 These studies, however, were not able to identify the bioactive components with antioxidant capacity in the SP water extract. NO contains ∼70 mg β-carotene per 100 g dry weight, which is higher than carrot, suggesting a potential antioxidant effect of NO. 51 Matsui et al. 78 purified two glycosylated mycosporine-like amino acids (MAA), water-soluble UV absorbing pigments, from NO. The novel MAA found in NO have been shown to exhibit radical scavenging and UV-protective activities, suggesting they may play a role in antioxidant activities of NO.
In summary, studies in cells, animals, and humans have suggested that several BGA species have a strong antioxidant property. C-PC has been recognized as a primary component with an antioxidant effect in BGA. However, other components, such as β-carotene, vitamins, and minerals, could also contribute to the effect. BGA decrease lipid peroxidation and ameliorate the oxidative damage caused by ROS and, therefore, they can prevent oxidative stress-induced chronic tissue damage and/or inflammation.
Protective effects of BGA against NAFLD
The prevalence of obesity and type II diabetes have led to increased incidence of NAFLD, the most common cause of chronic liver disease in Western countries. 79 In the Dallas Heart Study, the incidence of NAFLD was estimated to reach 30% of the general population in the United States. 80 Moreover, 60%–80% of obese individuals had fatty liver. 81,82 NAFLD was first described by Ludwig et al. 83 in the 1980s as hepatic inflammation in patients without alcohol consumption or other causes of hepatitis. Clinical symptoms of NAFLD range from fatty liver with over 5% of intracellular lipid accumulation to liver steatosis with infiltrated inflammatory cells and liver injury. 84 Although fat accumulation is a benign form of NAFLD with a minimal risk of progression, inflammation or oxidative stress can facilitate the progression to NASH, fibrosis, cirrhosis, and ultimately, hepatocellular carcinoma. 85 Additionally, fatty liver can increase the vulnerability of hepatocytes to secondary injurious insults such as oxidative damage or inflammation by decreasing hepatic antioxidant capacities and increasing activation of proinflammatory pathways. 86 Therefore, steatosis is a very important therapeutic target for the prevention of NASH.
Effect of BGA on NAFLD
Emerging evidence has suggested that BGA may lower hepatic lipids. As mentioned earlier in this review, previous studies from our laboratory demonstrated that NO lipid extract reduced lipogenic gene expression in HepG2 cells by inhibiting maturation of SREBP-1. 29 Therefore, NO lipid extract may have a protective role against the development of hepatic steatosis. The effects of SP on the prevention of NAFLD have been well documented. SM and SP are the most studied BGA species for their role in NAFLD. In animal models, fatty liver is commonly induced by CCl4 due to increased fatty acid synthesis and decreased release of lipoproteins. 87 Torres-Duran et al. 88 demonstrated that when Wistar rats received a single intraperitoneal dose of CCl4, supplementation of 5% whole SM prevented the accumulation of hepatic lipid and decreased plasma very low-density lipoprotein and LDL-C levels. Furthermore, when the rats were fed 5% defatted or lipid extract of SM, both treatments ameliorated CCl4-induced elevation in hepatic levels of total lipids, triglyceride, and total cholesterol, suggesting both fractions may contain active substances to prevent lipid accumulation. 23
Diabetes is often associated with NAFLD. 89 Rodríguez-Hernández et al. 90 investigated an effect of SM on the development of fatty liver in alloxan-induced diabetes in CD-1 mice. Dietary supplementation of 5% SM for 4 weeks after a single dose of alloxan prevented fatty liver in both male and female mice. Similarly, the hepatoprotective activity of SM was demonstrated in SM-supplemented diabetic Wistar rats that exhibited less fat accumulation in the liver and lower plasma LDL-C levels compared with control animals. 91 As alloxan-induced diabetes has similar characteristics to those of type I diabetes in humans, further study is necessary to evaluate effects of BGA on type II diabetes-associated NAFLD. In the livers of a monosodium glutamate-induced NASH mouse model, Spirulina supplementation at 5% for 12 weeks markedly decreased the NAFLD activity score and inflammation score. 92 The effects were observed concomitantly with lower levels of total cholesterol, triglycerides, nonesterified fatty acids, lipid peroxidation, TNFα, and IL-6 in the livers compared with control animals. In humans, patients with NAFLD were treated with 4.5 g/day of SM for 3 months and they showed significant improvement in plasma alanine transaminase, triglycerides, and the total cholesterol/HDL-C ratio. 93 In addition, ultrasonography demonstrated improved liver tissue texture in all patients after SM treatment. This study indicates potential pharmacological properties of SM as an alternative treatment for patients with NAFLD and dyslipidemic disorders. However, presently, no large-scale human studies have been conducted to evaluate hepatoprotective effects of BGA and, therefore, more clinical trials are needed.
Prevention of NASH via inhibition of inflammation and oxidative stress by BGA
Inflammation and oxidative stress can aggravate the progression of hepatic steatosis to NASH. In female Wistar albino rats treated with cadmium to induce oxidative stress, fat accumulation, and fibrosis, daily administration of 300 mg/kg SP for 30 days reduced hepatic MDA levels by 42%. 94 SP supplementation also elevated GSH, SOD, and nitric oxide levels, whereas vacuolar degeneration, fatty infiltration, and fibrosis were prevented in the liver. Similar antioxidant effects of SP supplementation on the prevention of hepatotoxicity were also described with decreases in the leakage of liver enzymes in plasma, hepatic lipid peroxidation, hemorrhage, and hepatocyte necrosis in the livers of rats with mercury-induced hepatic damage 22 as well as CCl4-induced hepatotoxicity. 95 Intraperitoneal administration of 200 mg/kg of C-PC from SP significantly reduced hepatotoxicity caused by R-(+)-pulegone or CCl4 in rats. 96
Overall, studies have demonstrated the role of SM or SP in the prevention of NAFLD. The beneficial effects of BGA appear be strongly associated with their antioxidant properties. Although mechanisms underlying the hepatoprotective effect of BGA have not been fully understood, BGA may be able to prevent fatty liver, at least in part, by inhibiting hepatic lipogenesis as described earlier. BGA can prevent progression of fatty liver to NASH by inhibiting lipid peroxidation and scavenging free radicals, or indirectly enhancing the activity of antioxidant enzymes in the liver. More studies are needed to identify functional components of BGA that are responsible for the hepatoprotective effects. One study up to date demonstrated that 4.5 g/day of BGA for 3 months was sufficient for reversing liver damage and exerting hypolipidemic effects in NAFLD patients. 93 However, information on the optimal BGA dose and duration of supplementation to gain beneficial effect of BGA for NAFLD is lacking. It is also unknown whether BGA intake can prevent the progress of steatosis in healthy subjects. As only limited human trials have focused on the protective effects of BGA against NAFLD, randomized double-blind, placebo-controlled clinical trials with a larger sample size are needed to support the role of BGA in hepatic protection in humans.
Conclusion
There has been an increasing demand for dietary interventions for the prevention of chronic inflammatory diseases, such as CVD and NAFLD. Studies in cells, animals, and humans, have demonstrated that edible BGA can be an effective natural product for improving blood lipid profiles and for preventing inflammation and oxidative stress. As described in Figure 1, BGA contain bioactive components, namely, carotenoids, GLA, PC, fibers, and plant sterols, which can be beneficial for preventing CVD and NAFLD. BGA inhibit intestinal cholesterol absorption and decrease hepatic lipids, lowering plasma total cholesterol, and triglyceride concentrations. The anti-inflammatory function of BGA is mediated, at least in part, by inhibiting the NF-κB pathway to decrease the production of proinflammatory mediators. BGA can also decrease oxidative stress due to their free radical scavenging activity and inhibition of lipid peroxidation. In conclusion, BGA can be consumed as a dietary supplement or a food component to obtain health benefits against CVD and NAFLD. However, safety assessments of any BGA species should precede recommendations for the BGA consumption in humans because contamination of toxin-producing BGA has been reported in some naturally harvested BGA products.
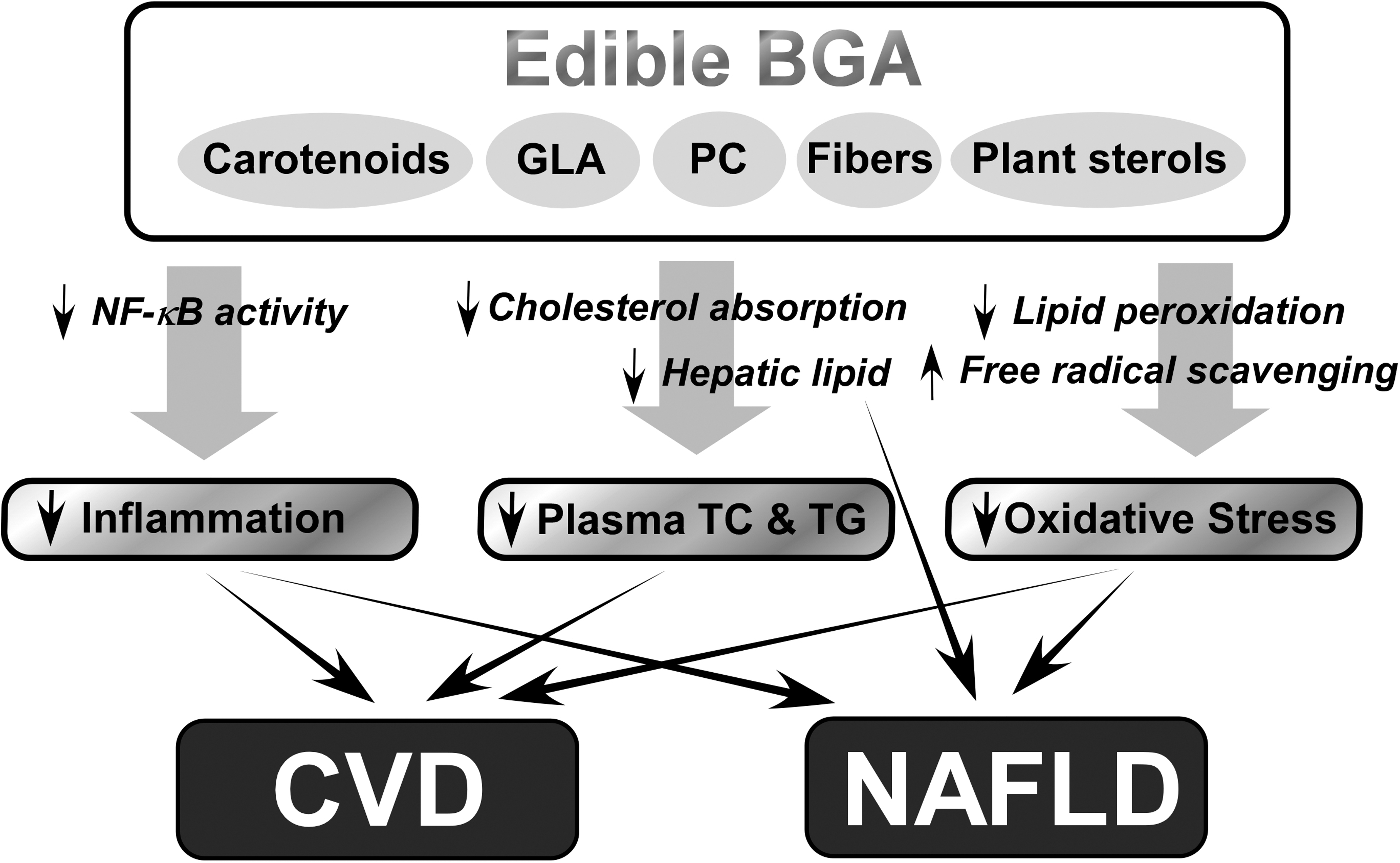
Summary of the protective effects of blue-green algae (BGA) against the development of cardiovascular disease (CVD) and nonalcoholic fatty liver disease (NAFLD). Chronic diseases, such as CVD and NAFLD, are highly related to impaired lipid metabolism, oxidative stress, and inflammation. Consumption of edible BGA containing carotenoids, γ-linolenic acid (GLA), phycocyanin (PC), fibers, and plant sterols provides multiple health-promoting properties: inhibition of inflammation via decreased nuclear factor kappa B (NF-κB) activity; lowering plasma lipid concentrations by decreasing intestinal cholesterol absorption and hepatic lipogenesis; and prevention of oxidative stress by blocking lipid peroxidation and increasing free radical scavenging.
Footnotes
Acknowledgments
This work was supported by the National Institute Health grant R21AT005152 and funds from College of Agriculture and Natural Resources at the University of Connecticut to J.L.
Author Disclosure Statement
All authors read this manuscript and claim no conflicts of interest.