
Abstract
The aim of this study was to evaluate the effect of salidroside on lipopolysaccharide (LPS)-induced nitric oxide (NO) and prostaglandin E2 (PGE2) production in RAW 264.7 macrophages and related anti-inflammatory mechanism. PGE2 production was measured by enzyme-linked immunosorbent assay (ELISA); NO production was tested by Griess reagent. Inducible nitric oxidesynthase (iNOS) and COX-2 were determined by RT-PCR and Western blot analysis; IκB and P-IκB protein express were detected by Western blot analysis; cytosolic free Ca2+ ([Ca2+]i)was measured by a fluorescent microscope. The data showed salidroside inhibited LPS-induced NO and PGE2 production and reduced iNOS and COX-2 protein expression in RAW 264.7 macrophages. Consistent with these observations, salidroside inhibited LPS-induced cytosolic free Ca2+ concentration ([Ca2+]i) elevation. In addition, we further investigated signal transduction mechanisms and found that the activation of NF-κB was suppressed by salidroside in a dose-dependent manner. These results suggest that salidroside suppresses NO and PGE2 production by inhibiting iNOS and COX-2 protein expression, level of [Ca2+]i, and activation of NF-κB signal transduction pathway.
Introduction
I
Rhodiola rosea has been widely used in traditional Chinese medicine for a long time. 6 Previous studies prove that it has various pharmacological functions, such as improving exercise endurance and fatigue, antimicrobial functions, anticancer activity, preventing high-altitude sickness, promoting blood circulation, eliminating toxins from the body, and treating various endemic diseases. 7 –11 Salidroside (p-hydroxyphenethyl-β-D-glucoside, C14H20O7) is a phenylpropanoid glycoside extracted from R. rosea and is regarded as the most important bioactive component. 12 It has been reported to have various pharmacological properties including anti-aging, anticancer, antivirus, antioxidative, and hepatoprotective effect. 13 –15 The aim of the present study was to investigate effects of salidroside on LPS-induced NO and PGE2 production in RAW 264.7 macrophages and associated mechanisms. Our results would be beneficial for salidroside to be further used as a potential treatment choice of inflammation diseases.
Materials and Methods
Materials
Salidroside (standard material, purity >98%. Fig. 1) was ordered from the National Institute for the Control of Pharmaceutical and Biological Products and directly dissolved in water. Methylene Blue trihydrate, LPS (Escherichia coli 055:B5), and nuclear protein extraction kit were purchased from Sigma Chemical Co. Fluo-3/AM was obtained from Dojin Laboratories. 5-(and-6)-chloromethyl-2′, 7′-dichlorodihydrofluorescein diacetate and acetyl ester (CM-H2DCFDA) was obtained from Invitrogen (Molecular probe). RPMI 1640 was obtained from Gibco. Human cyclic guanosine monophosphate ELISA kit and Human cyclic adenosine monophosphate ELISA kit were obtained from Blue Gene. FBS was obtained from Hangzhou. All other reagents were from commercial suppliers and of standard biochemical quality.

The structure of salidroside.
Cell culture and sample treatment
The RAW 264.7 mouse macrophage cell were cultured in DMEM supplemented with 3 mM glutamine, antibiotics (100 U/mL penicillin and 100 U/mL streptomycin), and 10% heat-inactivated FBS and maintained at 37°C in a humidified incubator with 5% CO2. In all experiments, cells were allowed to acclimate for 24 h before treatments.
Methylene Blue trihydrate assay for cell viability
Cell viability was determined by Methylene Blue trihydrate assay. RAW 264.7 cells (4×105 cells/mL) were plated onto 96-well plates and incubated overnight. The cells were treated with varying concentrations of salidroside (0–160 μg/mL) for 2 h and then stimulated with 1 μg/mL of LPS for 18 h. Fifty microliters of 0.6% Methylene Blue trihydrate was added to each well, and the cells were incubated for another 4 h at 37°C with 5% CO2. Methylene Blue trihydrate was removed, and the cells were lysed with 100 μL/well Methylene Blue trihydrate elution. The optical density was measured at 570 nm on a microplate reader (Tecan).
Measurement of NO production
RAW 264.7 cells (4×105 cells/mL) were plated onto 24-well plates and pretreated with 30, 60, or 120 μg/mL of salidroside for 1 h prior to stimulation with 1 μg/mL of LPS for 24 h. The cell supernatants were collected and assayed for NO production using Griess reagent. Briefly, the samples were mixed with equal volume of Griess reagent and incubated at room temperature for 15 min. The absorbance was measured at 570 nm on a microplate reader (Tecan).
Measurement of PGE2 production
RAW 264.7 cells were plated onto 24-well plates (4×105 cells/mL) and treated with 30, 60, or 120 μg/mL of salidroside 1 h before or after stimulation with 1 μg/mL of LPS. After 24 h of incubation, the supernatant of cultured medium was collected, and PGE2 production was determined by using an enzyme immunoassay kit according to the manufacturer's instructions.
RT-PCR assay for iNOS and COX-2 mRNA expression
RAW 264.7 cells (2×106) were incubated in the presence of either 1 μg/mL LPS alone or LPS plus tyrosol 40, 80, and 160 μg/mL for 24 h at 37°C with 5% CO2. Total RNA was isolated using easy-BLUETM kits according to the manufacturer's instructions and stored at −70°C until use. Briefly, integrity of RNA was confirmed by agarose gel electrophoresis, and RNA was quantified by spectrophotometric analysis. The PCR mixture was prepared according to the manufacturer's instructions using the following primers: mouse iNOS, forward 5′-AAG TCA AAT CCT ACC AAA GTG A-3′, and reverse 5′-CCA TAA TAC TGG TTG ATG AAC T-3′ 409 bp, mouse COX-2, forward 5′-GGA GAG ACT ATC AAG ATA GTG ATC-3′, and reverse 5′-ATG GTC AGT AGA CTT TTA CAG CTC-3′ 861 bp. Each reaction was performed using 2 μg of total RNA, and the thermocycler was programmed for reverse transcription at 60°C for 45 min, 95°C for 5 min, and 5°C for 5 min. The initial denaturation of the cDNA was accomplished at 94°C for 5 min, followed by 30 amplification cycles, each of which consisted of denaturation at 94°C for 30 s, 30 s of annealing (52°C and 60°C for iNOS and COX-2, respectively), and extension at 72°C for 1 min 30 s. These were followed by a final extension at 72°C for 10 min. Amplified PCR products were electrophoresed on a 1% TAE agarose gel.
Western blot analysis
RAW 264.7 cells (4×105 cells/mL) were plated onto six-well plates and pretreated with 30, 60, or 120 μg/mL of salidroside for 1 h and then stimulated with 1 μg/mL of LPS for 1 h (for iNOS and COX-2) or for 30 min (for IκB). The proteins from cells were extracted according to the manufacturer's instructions (Amersham Life Science). iNOS, COX-2, IκB, and P-IκB protein expression were tested by Western blot analysis.
Determination of [Ca2+]i
RAW 264.7 cells (4×105 cells/mL) were plated onto 6-well plates and pretreated with 30, 60, or 120 μg/mL of salidroside plus 1 μg/mL of LPS for 40 min. The [Ca2+]i was determined using Fluo-3/acetoxymethyl ester (Fluo-3/AM). Briefly, RAW 264.7 cells were loaded for 20 min at 37°C with 20 μM Fluo-3/AM containing 1 μM pluronic acid F-127 for proper dispersal and 0.25 mM sulfinpyrazone, to inhibit the leakage of the Fluo-3 dye. Shortly before use, a sample of loaded cells was washed with Krebs–Ringer–HEPES buffer to remove nonhydrolyzed Fluo-3/AM. Fluorescence measurements were performed using an inverted fluorescent microscope (Motic AE31invert).
Statistical analysis
All data are expressed as the mean±SD (n=3). Data analyses were completed using one-way ANOVA and Fisher's least significant difference post hoc test with SPSS 15.0 for Windows (SPSS, Inc.). Statistical significance was accepted when P<.05.
Results
Effect of salidroside on LPS-induced NO and PEG2 production in vitro
NO and PEG2 concentrations in the culture supernatant of RAW 264.7 cells were measured by sandwich ELISA. Treatment of RAW 264.7 cells with LPS alone resulted in significant increases. However, when we pretreated with 60 and 120 μg/mL of salidroside, NO and PGE2 levels significantly decreased compared to those of LPS group (P<.05 or P<.01). Salidroside at dose of 30 μg/mL could not significantly attenuate NO and PGE2 production (P>.05; Figs. 2 and 3).
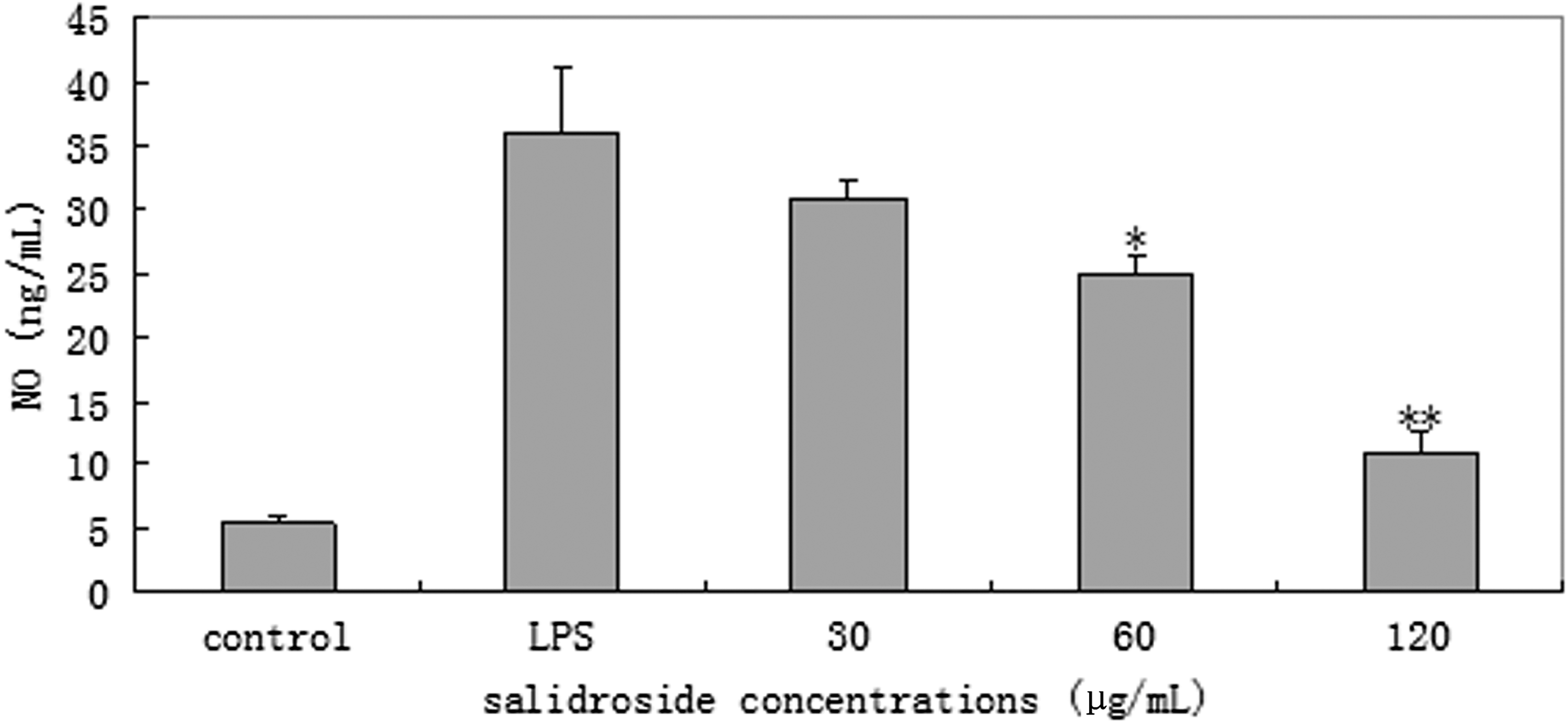
Effect of salidroside on the production of nitric oxide (NO) in lipopolysaccharide (LPS)-stimulated RAW 264.7 cells. RAW 264.7 cells (4×105 cells/mL) were pretreated with 30, 60, or 120 μg/mL of salidroside for 1 h prior to stimulation with 1 μg/mL of LPS for 24 h. The cell supernatants were collected and assayed for NO production using Griess reagent. The values presented are the mean±SD. *P<.05, **P<.01 versus LPS group.
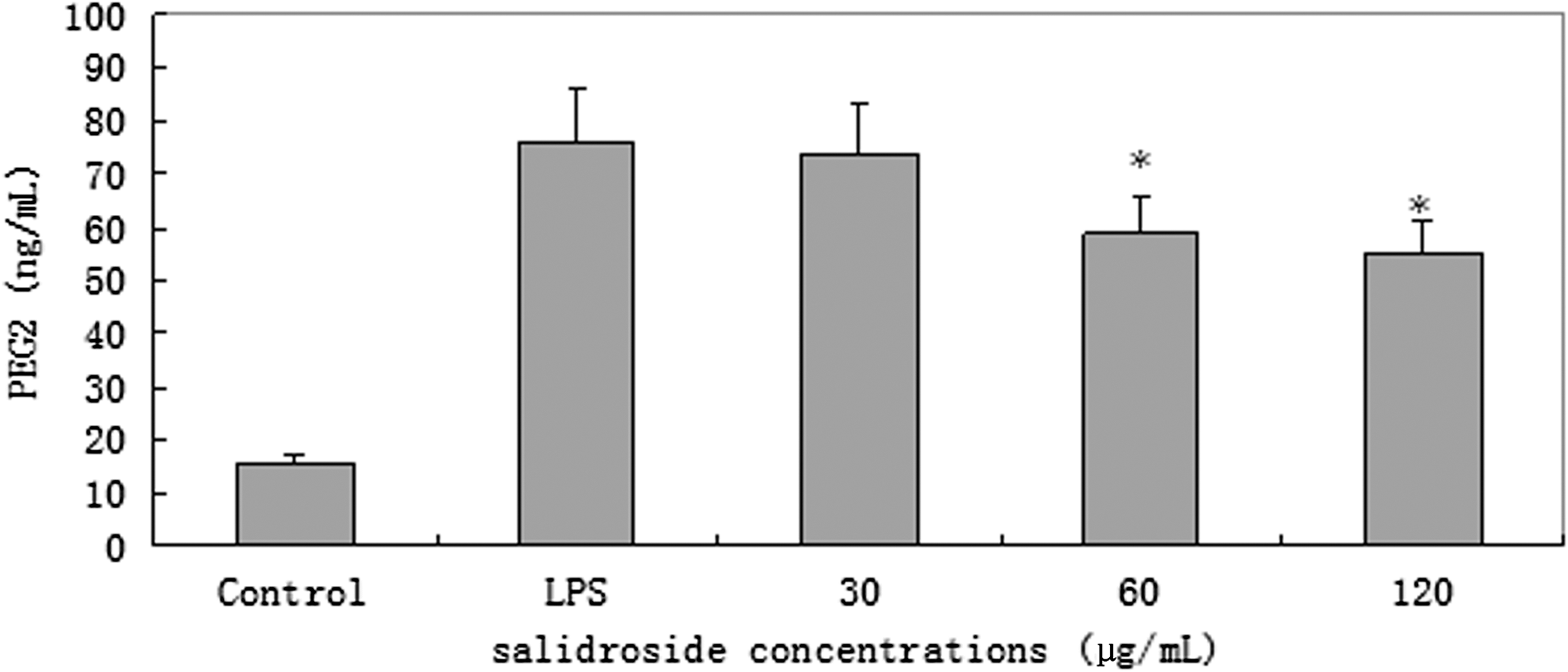
Effect of salidroside on the production of PEG2 in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells (4×105 cells/mL) were treated with 30, 60, or 120 μg/mL of salidroside 1 h before or after stimulation with 1 μg/mL of LPS. After 24 h of incubation, the supernatant of cultured medium was collected, and prostaglandin E2 (PGE2) production was determined by using enzyme-linked immunosorbent assay (ELISA). The values presented are the mean±SD. *P<.05 versus LPS group.
The cytotoxic effect of salidroside was evaluated by Methylene Blue trihydrate assay, and salidroside did not affect the cell viability of RAW 264.7 cells up to 160 μg/mL (data not shown). Thus, the inhibitory effects were not attributable to cytotoxic effects.
Effect of salidroside on LPS-induced iNOS and COX-2 production
RT-PCR analysis was performed to determine whether the inhibitory effects of salidroside on the inflammatory mediators (NO and PGE2) were related to the modulation of iNOS and COX-2. As shown in Figure 4, we found that LPS could significantly induce mRNA expression for iNOS and COX-2, whereas treatment of cells with salidroside inhibited these mRNA expressions. In the meantime, we also tested of the expression of iNOS and COX-2 by Western blot analysis (Figs. 5 and 6). The outcomes were consistent with RT-PCR. These results indicated that the inhibitory effects of salidroside on LPS-induced NO and PGE2 production are caused by iNOS and COX-2 suppression.
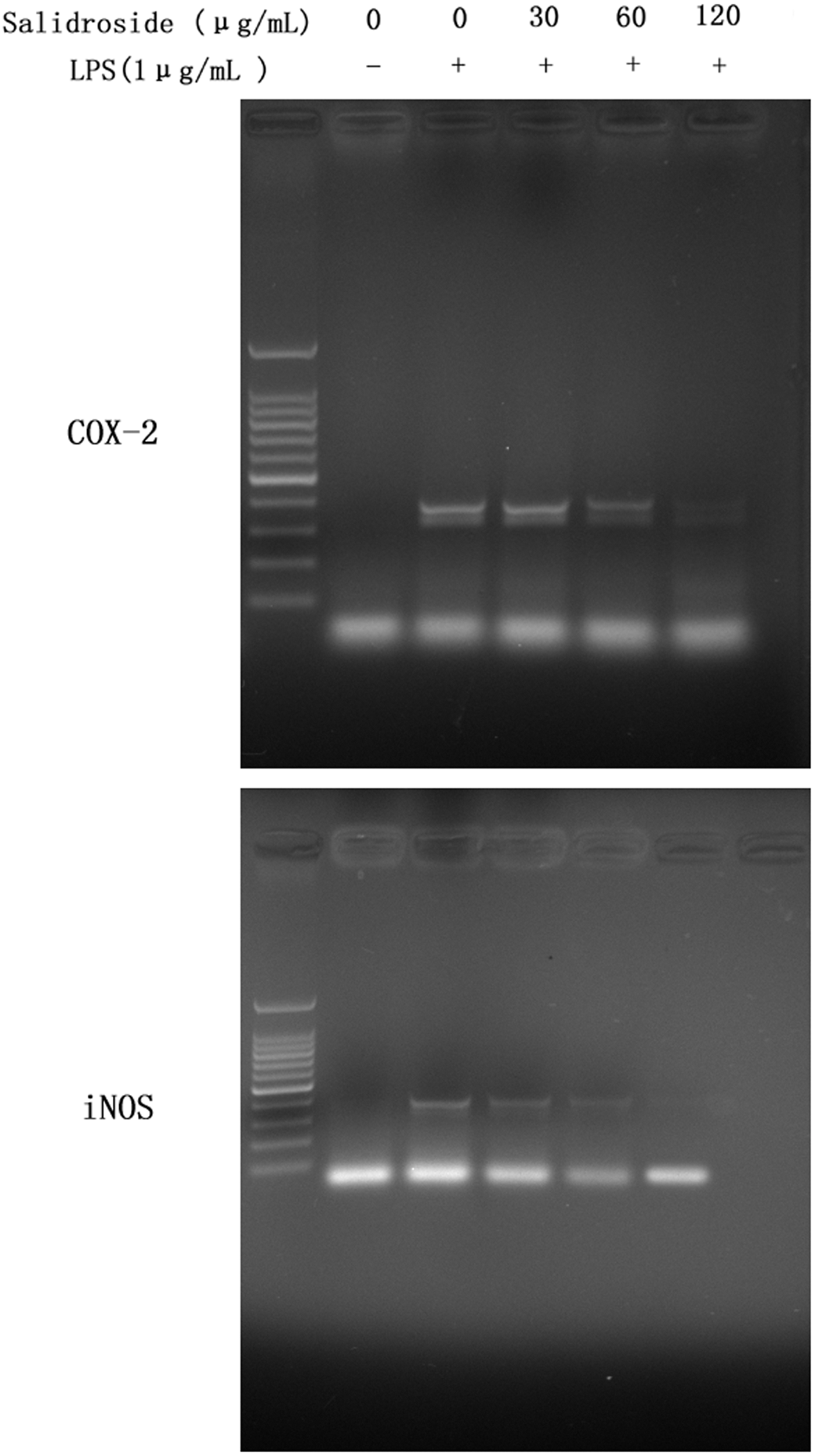
Effect of salidroside on mRNA expression of COX-2 and inducible nitric oxidesynthase (iNOS) in LPS-induced RAW 264.7 cells. The cells were treated with 1 μg/mL of LPS alone or LPS plus different concentrations (30, 60, and 120 μg/mL) of salidroside for 24 h. Total RNA was isolated and mRNA expressions were measured by RT-PCR. Shown in the down panel were data from three independent experiments. A representative RT-PCR is shown.
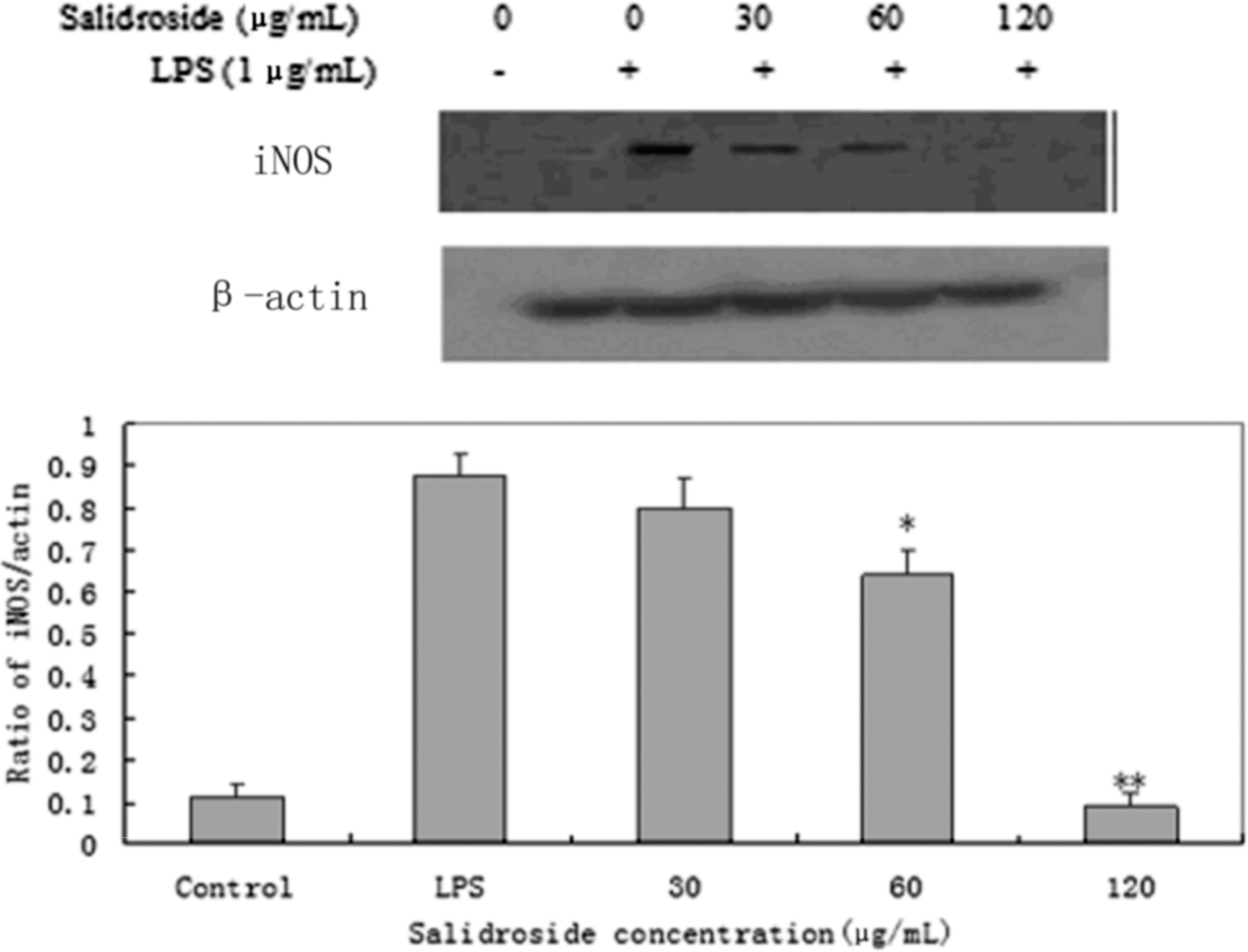
Effect of salidroside on the expression of iNOS protein in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells (4×105 cells/mL) were pretreated with 30, 60, or 120 μg/mL of salidroside for 1 h and then stimulated with 1 μg/mL of LPS for 1 h. The protein from cells was extracted according to the manufacturer's instructions. iNOS protein expression were tested by Western blot analysis. *P<.05 or **P<.01 versus LPS group.
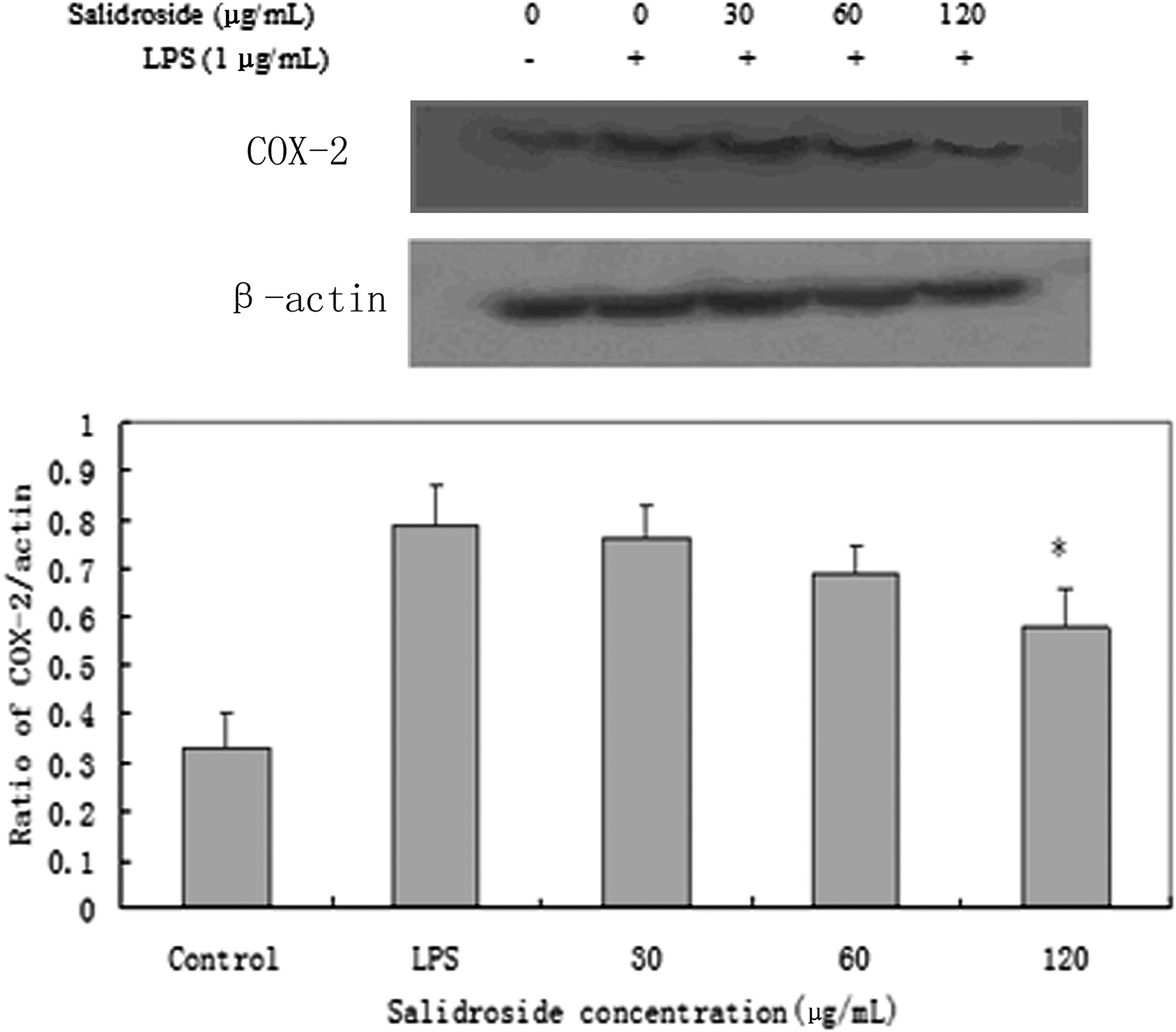
Effect of salidroside on the expression of COX-2 protein in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells (4×105 cells/mL) were pretreated with 30, 60, or 120 μg/mL of salidroside for 1 h and then stimulated with 1 μg/mL of LPS for 1 h. The protein from cells was extracted according to the manufacturer's instructions. COX-2 protein expression was tested by Western blot analysis. *P<.05 versus LPS group.
Salidroside inhibited LPS-induced [Ca2+]i elevation
We examined the effect of salidroside on [Ca2+]i production. We found that incubating with LPS for 40 min induced [Ca2+]i elevation significantly, while preincubation with salidroside dose dependently suppressed the elevation of [Ca2+]i (Fig. 7).
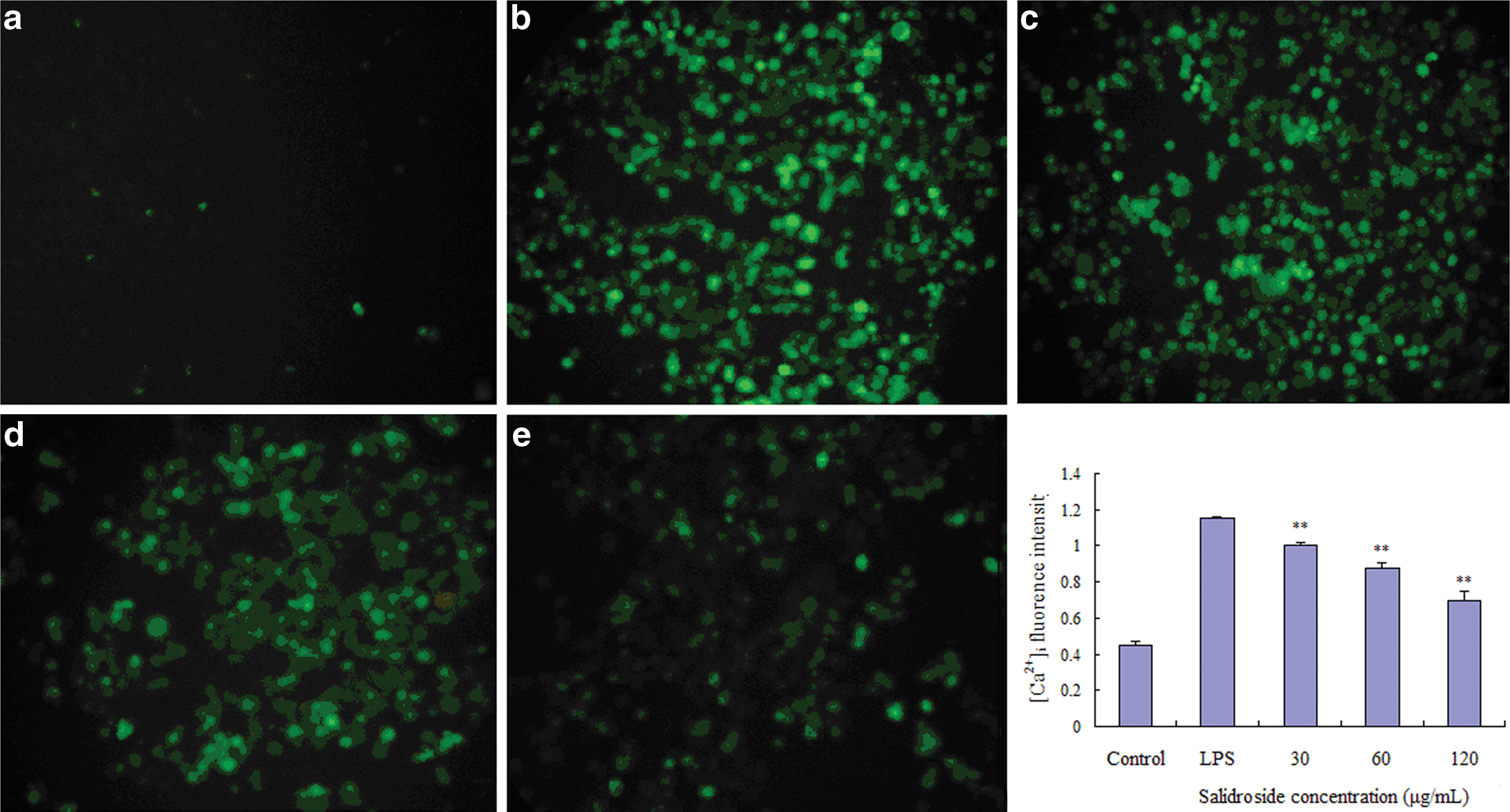
Effect of salidroside on the concentration of [Ca2+]i in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells (4×105 cells/mL) were pretreated with medium alone
Measurement of IκB and P-IκB
As shown in Figure 8, the results showed that the P-IκB in RAW 264.7 cells increased after LPS administration but was significantly inhibited by salidroside in a concentration dependent manner (P<.05).
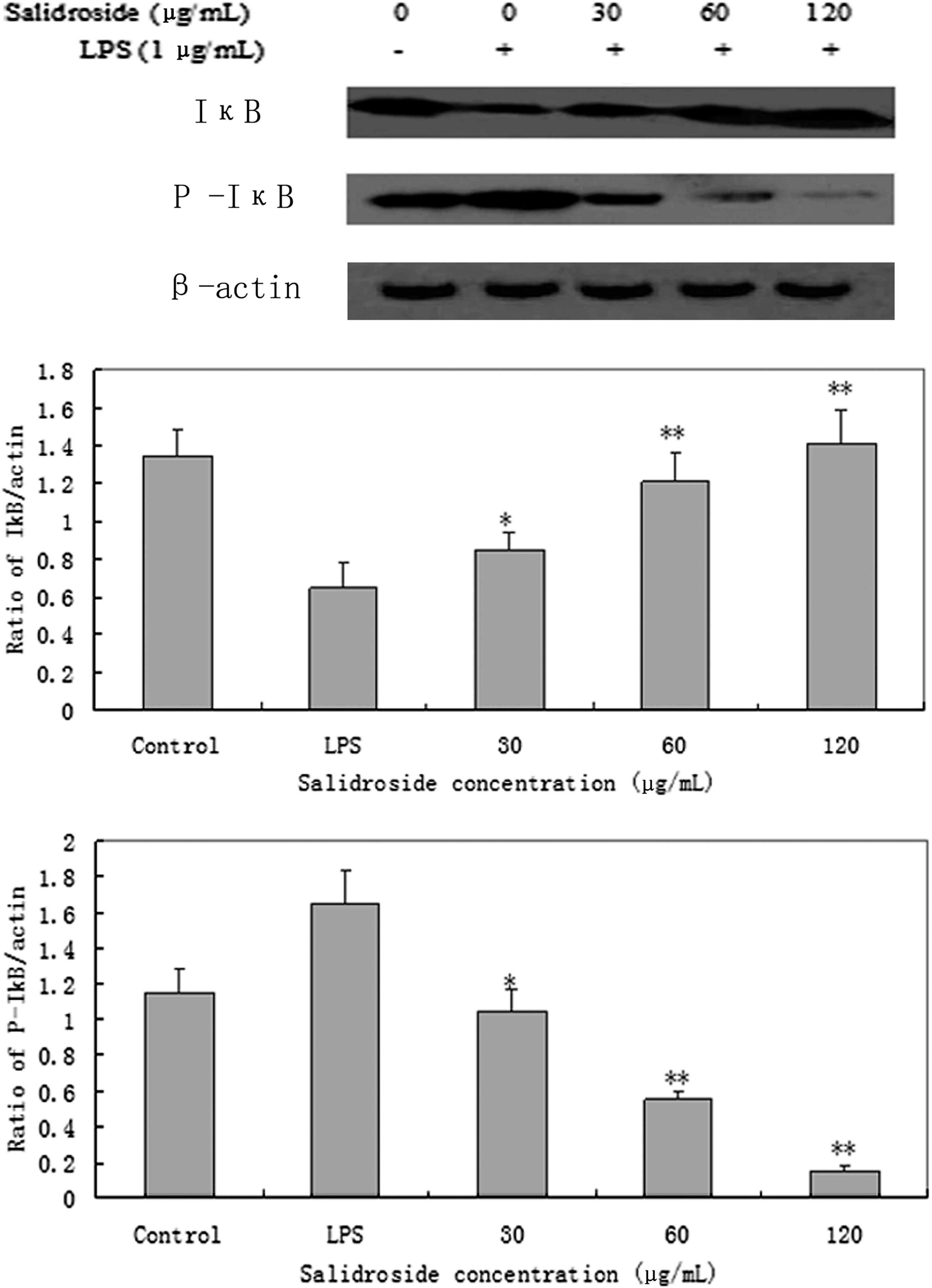
Effect of salidroside on the expression of IκB and P-IκB protein in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells (4×105 cells/mL) were pretreated with 30, 60, or 120 μg/mL of salidroside for 1 h and then stimulated with 1 μg/mL of LPS for for 30 min. The protein from cells was extracted according to the manufacturer's instructions. IκB and P-IκB protein expression were tested by Western blot analysis. *P<.05 or **P<.01 versus LPS group.
Discussion
NO and PGE2 are major inflammatory mediators during the inflammatory process and generated by the iNOS and cyclooxygenase-2 (COX-2). 16,17 Excessive production of these inflammatory mediators will result in many inflammatory-related diseases, such as SIRS, severe tissue damage, and septic shock. 18 Thus, inhibition of the production of these inflammatory mediators is an important target for anti-inflammatory treatment in clinic.
In this experiment, NO and PGE2 concentrations in the culture supernatant of RAW 264.7 cells were measured. Treatment of RAW 264.7 cells with LPS alone resulted in significant increases. However, when treated with 60 and 120 μg/mL of salidroside, NO and PGE2 levels significantly decreased compared with those of LPS group (P<.05 or P<.01). Salidroside at dose of 30 μg/mL could not significantly attenuate NO and PGE2 production (P>.05; Figs. 2 and 3). The cytotoxic effect of salidroside was evaluated by Methylene Blue trihydrate assay, and salidroside did not affect the cell viability of RAW 264.7 cells at the concentrations (0–160 μg/mL) used to inhibit NO and PGE2 (data not shown). Thus, the inhibitory effects were not attributable to cytotoxic effects.
Western blot analysis and RT-PCR were performed to determine whether the inhibitory effects of salidroside on the inflammatory mediators (NO and PGE2) were related to the modulation of the expression of iNOS and COX-2. We found that treatment with salidroside significantly inhibited the upregulations caused by LPS (Figs. 4 –6). These results indicated that the inhibitory effects of salidroside on LPS-induced NO and PGE2 production are caused by iNOS and COX-2 suppression.
We further study the change of calcium in this process. Calcium is one of the basic elements needed to maintain life activities. Cytosolic free Ca2+ ([Ca2+]i) could regulate many physiological and metabolic processes as an important second messenger. Meanwhile, it participates in and coordinates with other second messenger metabolism and signal transmission. [Ca2+]i elevation is a necessary factor of activation and release of some inflammatory-related enzymes and medium. Regulating the concentration of [Ca2+]i is an effective way to regulate inflammatory response. 19,20 In our research, LPS successfully resulted in [Ca2+]i elevation in RAW 264.7 cells. Treatment of salidroside significantly suppressed production of LPS-induced [Ca2+]i in a dose-dependent manner.
Activation of NF-κB transcription pathway could regulate the expression of many genes with important functions in immune responses and inflammation, and the activation of NF-κB depends on changes in intracellular Ca2+ levels. 21 In unstimulated condition, NF-κB is localized to the cytosol binding with IκB. However, when cells are activated by LPS, IκB is phosphorylated by IκB kinase and degraded, then the released NF-κB is translocated into the nucleus. 22 Therefore, the activation of NF-κB could be assessed in RAW cells by measuring the degree of phosphorylation of IκB protein. Treatment with LPS resulted in the degradation of IκB and this degradation was significantly blocked by salidroside. To determine whether this IκB degradation was related to IκB phosphorylation, we examined the effect of salidroside on LPS-induced P-IκB by Western blot. The results showed that the P-IκB in RAW 264.7 cells increased after LPS administration but was significantly inhibited by salidroside in a concentration-dependent manner (P<.05; Fig. 8).
In conclusion, these outcomes suggest that salidroside isolated from the whole plant of R. rosea L. attenuated LPS-induced NO and PGE2 by suppressing iNOS and COX-2 protein expression, inhibiting [Ca2+]i, and preventing NF-κB pathway activation. Further studies should focus on the clinical usefulness of salidroside.
Footnotes
Acknowledgments
This study was financed by the National Nature Science Foundation of China (No. 30972212).
Author Disclosure Statement
No competing financial interests exist.