
Abstract
The present study aimed to evaluate the effect of kai xin san (KXS, at doses of 500, 250, and 125 mg/kg body weight per day), a well-known traditional Chinese medicine, on learning and memory in paradoxical sleep deprivation (PSD)-induced cognition deficit rats. Two behavior tests (the Open Field test and the Morris water maze task) were used for testing the effects of KXS on a PSD-induced learning and memory deficit model. Furthermore, its effect on the glutamic acid (GLU) and γ-amino-butyric acid (GABA) levels in the brain tissue, brain-derived neurotrophic factor (BDNF), cyclic AMP response element binding protein (CREB), and phosphorylated-CREB (p-CREB) expression in the hippocampus was also tested. KXS exerted the greatest cognition against the 48 h PSD-induced cognitive deficit and these effects may be mediated by decreasing the GLU and GABA levels and increasing the levels of BDNF, CREB, and p-CREB. This study indicates that the effect of KXS on learning and memory in a rat model of PSD could be associated with the modulation of neurotransmitter levels and the expression of some genes in the brain that contribute to memory functions.
Introduction
S
The regulation of learning and memory requires cooperation between excitatory and inhibitory neural systems, the representatives of which are the glutamatergic and γ-amino–butyric acid (GABA)–ergic systems, respectively. 5 The hippocampus is one of the essential brain regions activated in learning and memory. A certain balance of neuronal transmission between the major excitatory neurotransmitter glutamate and the inhibitory substance GABA is required to maintain normal functions of the brain, including learning and memory. 6 The decrease of glutamate or increase of GABA can lead to impairment of learning and memory. 7 The sole path to produce GABA in the brain is with glutamic acid (GLU) as the substrate and GLU decarboxylase as the catalyst.
The brain-derived neurotrophic factor (BDNF) and cyclic AMP response element binding protein (CREB) are also important modulators of hippocampal-dependent learning and long-term potentiation (LTP). Studies have shown that memory performance correlates well with the activation of the CREB, and with increases in both the pro- and mature levels of the BDNF in the hippocampus. 8 It has also been shown that increasing the BDNF levels significantly improves spatial learning and memory retention. 9 Phosphorylation of the CREB plays an important role in the formation of synapses and synaptic plasticity. 10
Kai xin san (KXS) is a traditional Chinese herbal formula that contains ginseng (the radix of Panax ginseng C.A. Meyer), fu ling (the white part of Poria cocos Wolf), yuan zhi (the radix of Polygala tenuifolia Willd), and shi chang-pu (the rhizome of Acorus gramineus Solander). KXS is a widely used formula of traditional Chinese medicine (TCM) derived from Bei-Ji-Qian-Jin-Yao-Fang, an ancient medical classic written by Sun Si-miao in 652 AD. Previous studies have shown that KXS can cure symptoms, including desolation, moodiness, and forgetfulness, which are similar to such neuroses as depression, anxiety, and impairment in learning and memory. KXS has various pharmacological effects, such as antidepressant-like effects in different depression models, 11,12 and it also can ameliorate the learning and memory deficit in different memory-lesioned mice. 13 Moreover, in clinical practice, it has been used for the treatment of neurasthenia, Alzheimer's disease, and neurosis.
Although this decoction has been used frequently, the effect and mechanism of KXS for PSD-induced cognition deficit is still unknown. In this study, 14 days of treatment with a chemically standardized herbal extract of KXS suppressed PSD-induced learning and memory lesion in rats. In the brain of KXS-treated rats, the levels of neurotransmitters (GABA and GLU), the gene expressions of neurotrophic factors, and its transcription factor were significantly altered.
Materials and Methods
Preparation of KXS
KXS, supplied in the form of a dried powder herbal extract, was obtained from the Beijing Tongrentang Drug store (Beijing, China). All medicines formulating KXS (namely, radix of P. ginseng, white part of W. cocos, radix of P. tenuifolia, rhizome of A. gramineus) were purchased from the Chengdu Luye Medicinal Material Plantation Co., Chengdu, China. The voucher specimens of the four plants were identified by Prof. Ping Liu and registered under the numbers NU-90111, NU-82003, NU-79015, and NU-80617, respectively, and were preserved at the herbarium of the Traditional Chinese Medicinal Pharmacy, Chinese People's Liberation Army (PLA) General Hospital. Preparation of the total extract is the same as previously reported. 14
Analysis of chemical contents of KXS by HPLC
The chemicals used for the identification and quantification of compounds in the KXS extract included the following: 3,6′-disinapoyl sucrose, tenuifoliside A, and cis-2,4,5-trimethoxy-1-propenybenzene. A sample of the KXS extract (400 mg) was dissolved in 10 mL of solvent (methanol: H2O=1:1) and filtered through a membrane filter (0.45 μm; Alltech) before injection into the HPLC system. The HPLC system was composed of the L-2200 Autosampler (Hitachi), L-2130 pump (Hitachi), DAD L-2450 Detector (Hitachi), and Aglient HC C18 (4.6 mm×250 mm) column. The mobile phase was composed of acetonitrile (A) and 0.1% (v/v) phosphoric acid in water (B) was used as follows: 10% A/90% B (start), 16% A/84% B (15 min), 20% A/80% B (20 min), 26% A/74% B (40 min), and 90% A/10% B (90 min). The flow rate was 1.0 mL/min. The detector wavelength was set at 310 nm.
Regression equations for 3,6′-disinapoyl sucrose, tenuifolisde A, and cis-2,4,5-trimethoxy-1-propenybenzene were prepared from determinations using standard solutions of different concentrations quantified by HPLC.
Animals
Male Sprague–Dawley rats (body weight 250–270 g) were purchased from the Animal Breeding Center of the PLA General Hospital. They were housed one group per cage with free access to food and water, and maintained on an automatic 12-h light/12-h dark cycle (lights on at 7:00 a.m.) under controlled conditions of temperature (22°C±2°C) and humidity (55%±10%). The experimental protocols followed the Guide for the Care and Use of Laboratory Animals of Shanghai Institute of Material Medica and carried out according to international guidelines on the ethical use of laboratory animals. The experiments in the present study were designed to minimize the number of animals used and their suffering.
Experimental protocol
The rats were randomly assigned to six groups of six each: control, vehicle, modafinil (supplied by Chinese PLA Military Academy of Medical Sciences; 30 mg/kg body weight [b.w.] per day), KXS (500 mg/kg b.w. per day), KXS (250 mg/kg b.w. per day), KXS (125 mg/kg b.w. per day). The extracts of KXS were dissolved in distilled water and were administered during 14 days, including the training trial and the behavioral tests. The control and vehicle groups were administered distilled water (10 mL/kg b.w.) daily. Modafinil was also suspended in distilled water. Modafinil and KXS were administered intragastrically 40 min before the training trial and the behavioral tests. Previous studies in our laboratory and reported in the literature demonstrated that behavioral test performances between rats placed on large platforms as the PSD environment control group and rats in the control group did not differ significantly. Therefore, there is no PSD environment group in this study. After drug or distilled water for 6 days, all animals performed four consecutive trials each day over a 5-day training period in the Morris water maze. Later, animals were submitted to 48 h PSD except the control group. Behavioral tests were performed after 48 h sleep deprivation. After performing the probe trial test, the rats under deep anesthesia with 10% chloral hydrate (0.4 mL/kg b.w.) were sacrificed. Figure 1 shows the protocol of the PSD test.

Experimental protocol of the timeline depicting the behavioral tests adaptation and the behavioral tests.
PSD procedure
Rats in PSD groups were deprived of REM sleep for 48 h using the modified multiple platform method (MMPM). 15 Loss of muscle tone during the REM sleep phase caused the rat snout to dip into the water or sometimes the rat itself to fall into the water and awaken. In the PSD groups, six rats from the same cage were placed in a large aquarium (70 cm×47 cm×45 cm) at room temperature (23°C±1°C). The aquarium contained 8 columns (cross-section diameter: 6.5 cm, with platforms 1 cm above the water level), spaced 10 cm apart (edge to edge) such that rats could move freely from one platform to another. During the sleep deprivation period, rats had free access to water bottles and food pellet baskets hanging from the aquarium cover. 16 The water in the tanks was changed daily.
Open Field test
The Open Field test was carried out at the beginning and the end of 48 h PSD. The test was modified from previously described methods. 17 Briefly, the Open Field apparatus consisted of a square plastic arena (80 cm×80 cm×40 cm) with a black surface covering the inside walls. In the test, a single rat was placed in the center of the arena and allowed to explore freely. Each rat was placed individually in the center of the apparatus and following behaviors were scored for 5 min. The distance of the rats movement and the number of rearings (raising of the front paws) were recorded by ANY-maze (Stoelting Co.). This apparatus was cleaned with a detergent and dried after occupancy by each rat.
Morris water maze task
The Morris water maze was a black circular tank (160 cm in diameter, 50 cm deep), and was divided into four quadrants (1–4) of equal size on the monitoring screen of a computer. A hidden black platform (10 cm in diameter) was placed in the middle of the quadrant 3, submerged 1.5 cm below the water surface. The water in the tank was colored by a nontoxic black dye, and was kept at 23°C±1°C to a depth of 30 cm.
Daily, for 5 days, the rats were trained between 9:00 a.m. and 2:00 p.m. in four trials to find the hidden platform according to the distal cues provided by colored posters on the walls of the room. Each rat was released facing the wall of the maze from different positions around the perimeter, respectively. In each trial, a rat was allowed up to 2 min to locate the platform and was then left on it for 30 sec. If the platform was not found in 2 min, the rat was guided onto it and left there for 30 sec. A 3–5-min interval was allowed between each trial. The rats were dried with a towel and returned to their cage at the end of each trial. The time (escape latency) and distance (escape path length) swum by a rat to find the hidden platform were recorded. The escape latency to find the hidden platform was recorded automatically using the Video Tracking and Analysis System (Yishu Technology).
After 48 h PSD and the Open Field test, a place navigation test was given to detect the effect of PSD on the maintenance of memory. The place navigation test was the rats were released from the quadrant 1 to find the hidden platform in the quadrant 3 and its escape latency were recorded. After 3 h of the place navigation test, the hidden platform was removed from the water maze in the probe trial. The animal was placed in quadrant 1, which was opposite to the target quadrant (quadrant 3), and was allowed to swim for 120 sec in the water maze. The time and the distance that an animal spent in the target quadrant and the number of times that the same animal crossed the former platform area were used to measure the spatial memory maintenance.
Determination of GLU and GABA levels in brain tissue
After the rats were sacrificed, the whole brain of each rat was rapidly removed and chilled in an ice-cold saline solution. The brain was separated on ice, longitudinally bisected along the axes. The left cerebral hemisphere was homogenized with ice-cold saline to make a 10% (w/w) homogenate, and then centrifuged at 5600 g at 4°C for 10 min. The clear supernatants were collected and stored at −80°C for detection. The hippocampus of the right hemisphere was dissected on a cold plate and immediately frozen in liquid nitrogen. We assayed GLU and GABA in supernatant fluid using the ELISA kit (Nanjing Jiancheng Institute of Biological Engineering).
Western blotting
Western blot for BDNF, CREB, and phosphorylated CREB (p-CREB) protein in hippocampus were performed to assess the quantitative changes in the levels of these proteins. Frozen specimens were homogenized in 300 μL of the RIPA buffer containing 50 mM Tris-HCL (pH7.4), 150 mM NaCl, 1% NP-40, 0.1% sodium dodecyl sulfate (SDS), and 1 mM PMSF. Following centrifugation (13,100 g for 10 min, 4°C), the supernatant was used to estimate the total amount of protein. Samples were subjected to 12% SDS–polyacrylamide gel electrophoresis with equal amount of protein loaded per lane. Proteins were transferred to a nitrocellulose membrane. After blocking of nonspecific binding sites with T-TBS containing 3% bovine serum albumin 2 h at room temperature, membranes were incubated 4°C overnight with one of the primary antibodies used rabbit anti-BDNF (1:1000), rabbit anti-CREB (1:1000), or rabbit anti-p-CREB (1:500; Abcom). The secondary antibody, HRP-conjugated goat anti-rabbit IgG (1:5000; Abcom), was applied for 1 h. After rinsing with a buffer, the immunocomplexes were visualized by chemiluminescence using the ECL kit (Engreen Biosystem Co., Ltd.). Protein expression in the hippocampus was determined by measuring the optical density using an image analysis system (EG3 Imaging system; UVP Ltd.).
Statistical analysis
All data were presented as mean±SEM. The data were analyzed by one-way analysis of variance and significance of differences was determined by Dunnett's post hoc tests, P<.05 and P<.01 were considered significant.
Results
HPLC analyses of KXS
The major components in KXS extract were determined to be the following: 3,6′-disinapoyl sucrose 1.81±0.08 mg/g, tenuifolisde A 1.36±0.07 mg/g, and cis-2,4,5-trimethoxy-1-propenybenzene 2.56±0.10 mg/g (Fig. 2).
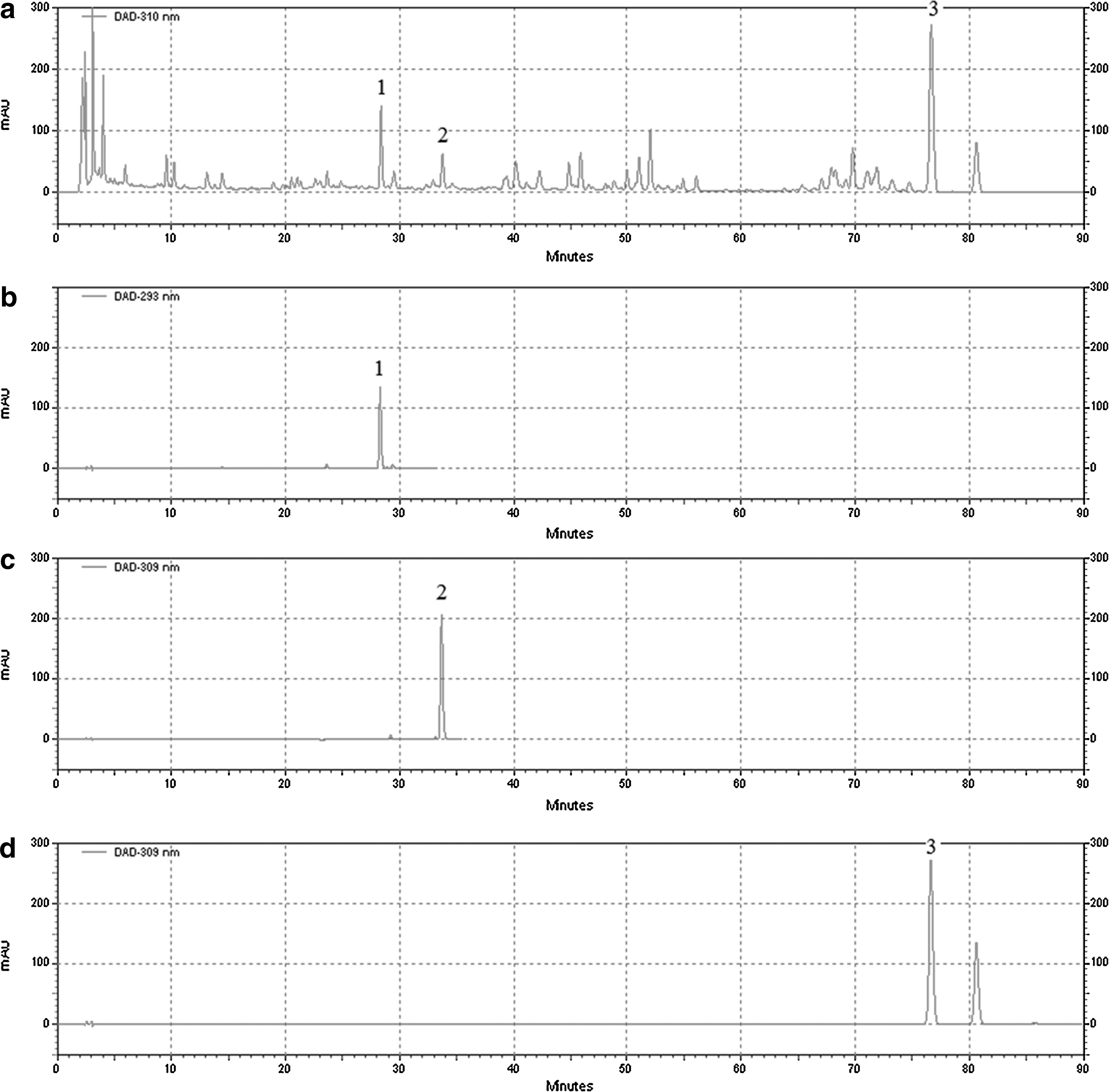
Chromatograms of standard components and kai xin san (KXS).
Effect of KXS on locomotor activity in the Open Field test
In the present study, we investigated the effect of KXS treatment on the locomotor activity in PSD rats (Fig. 3). The locomotor activity was assessed by the number of crossings and rearings in the Open Field test. Before PSD treatment was given, there was no significant difference in the number of crossings and rearings among rats (data not shown). At the 48 h of PSD, there was a significant reduction in the number of crossings and rearings in rats (Fig. 3a and b, respectively; P<.05), as compared to the controls. Treatment with KXS at daily doses of 125–500 mg/kg b.w. significantly and dose dependently increased the number of crossings and rearings in PSD rats (P<.05 or P<.01), as compared to the vehicle controls.

Effect of KXS treatment (500, 250, 125 mg/kg body weight per day) on the number of crossings
Effect of KXS on escape latency in the place navigation test
Figure 4a shows the effect of KXS treatment on escape latency in PSD rats. After 48 h PSD, the vehicle control rats had significantly increased escape latency (P<.05), as compared to the controls. Treatment with KXS at daily doses of 125 and 250 mg/kg b.w. significantly reduced the escape latency in PSD rats (P<.05 or P<.01), equally effective as modafinil.
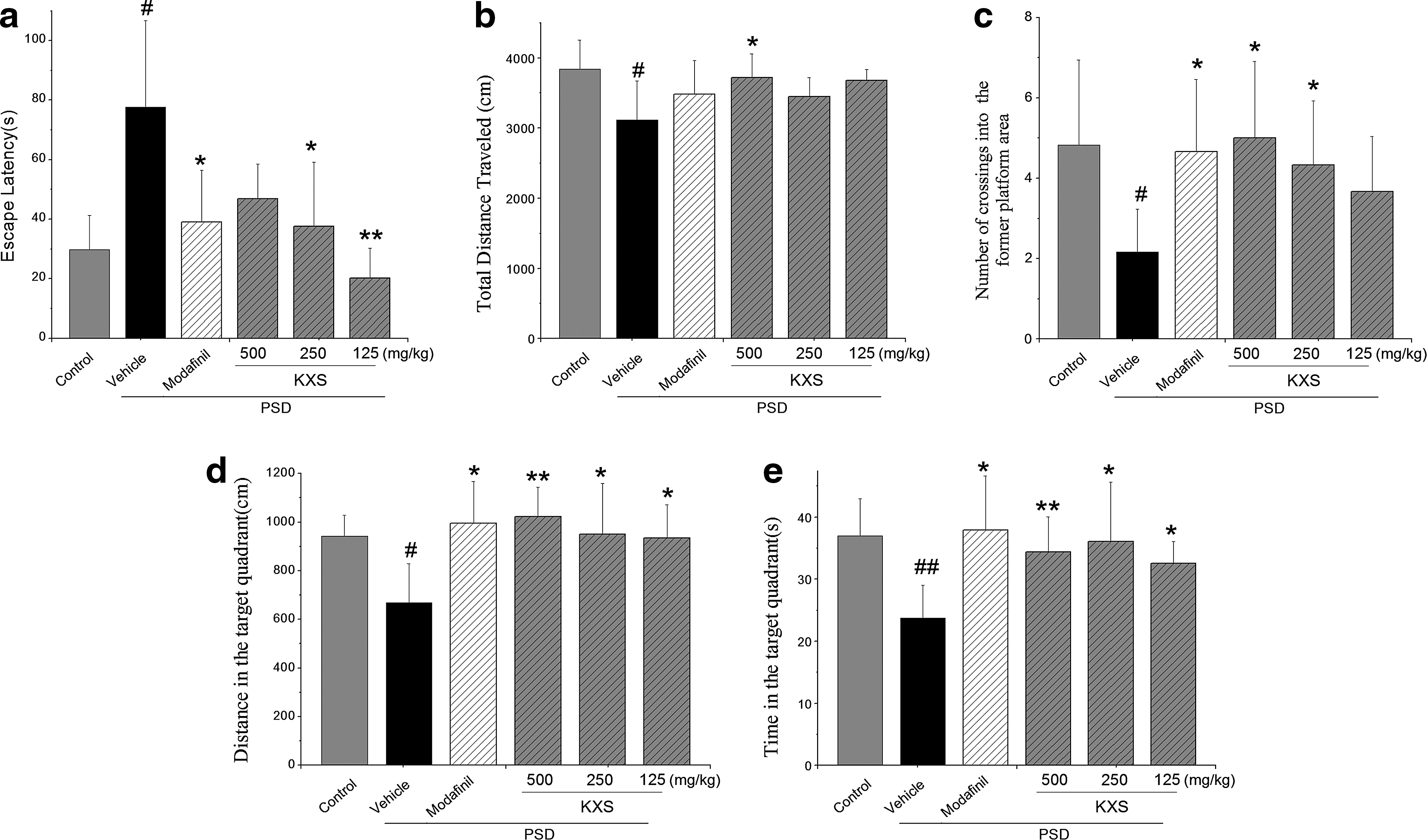
Effect of KXS on the Morris test, including escape latency in the place navigation test
Effect of KXS on spatial memory in the probe trial test
After PSD, a probe trial was used to assess the spatial memory. As shown in Figure 4b–e, it is notable that rats in the vehicle group traveled the shortest total distance, passed fewer numbers across the former platform, swam notably less distance in the target quadrant, and spent less time in the target quadrant (P<.05 or P<.01), as compared to the controls. However, the KXS with a different dosage, significantly increased the total distance, increased the number of crossings, and increased the distance and time in the target quadrant than the vehicle control (P<.05 or P<.01).
Effect of KXS on GLU and GABA levels
PSD significantly increased both GLU and GABA levels (P<.05 compared to control, Fig. 5). The modafinil group failed to show a significant reduction in the index level in the brain (P>.05 compared to vehicle). As shown in Figure 5a, KXS (500, 250, 125 mg/kg b.w. per day) treatment remarkably (P<.05 all, compared to vehicle) reduced the GLU level in cerebral tissues of PSD treated rats. Only rats subjected with high dosage KXS (500 mg/kg b.w. per day) showed a notably decreased GABA level (P<.01 compared to vehicle, Fig. 5b).
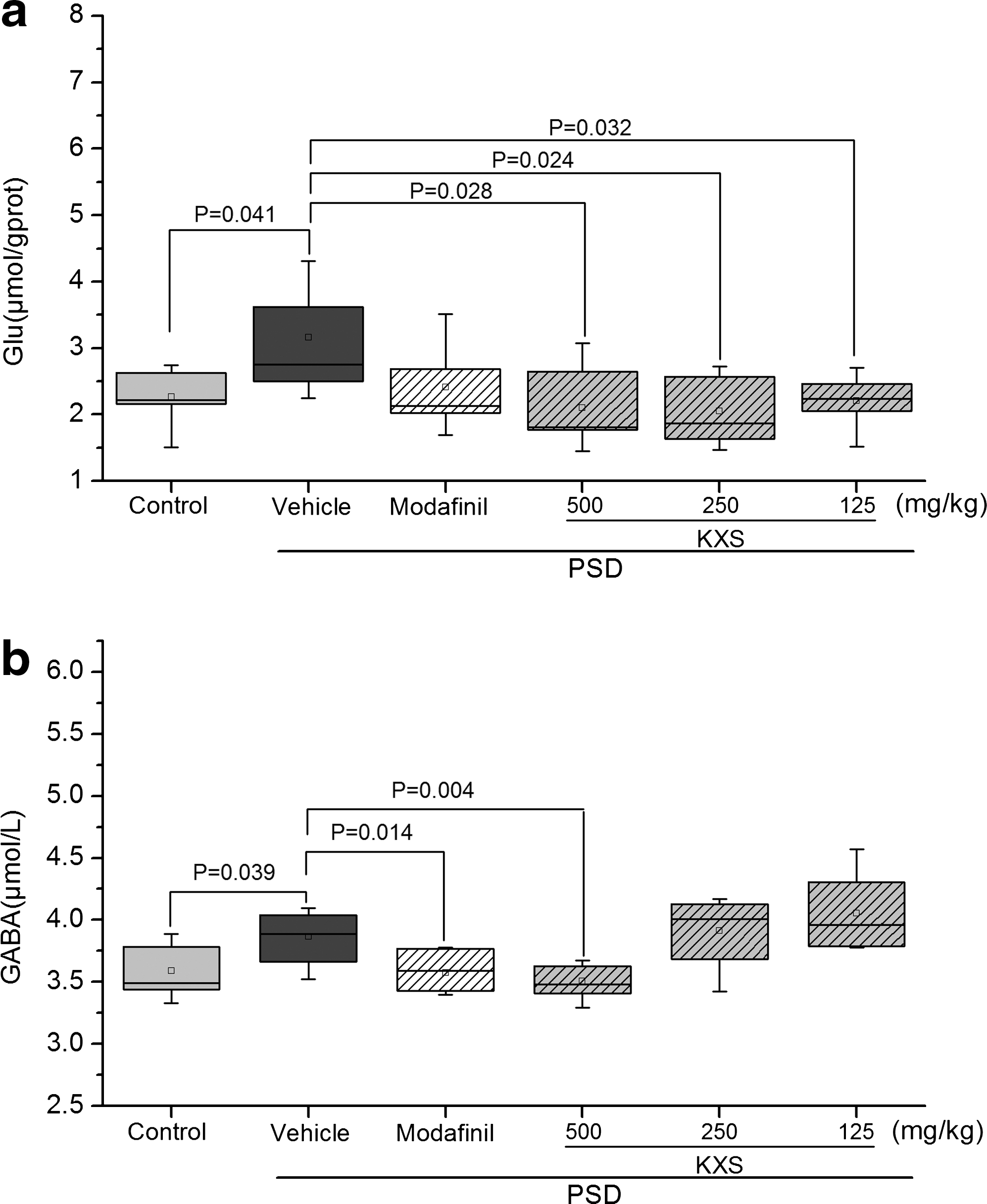
Effect of KXS treatment (500, 250, 125 mg/kg b.w. per day) on the levels of GLU
Effect of KXS on BDNF, CREB, and p-CREB protein levels in the hippocampus
PSD significantly decreased BDNF and p-CREB/CREB protein levels in the hippocampus. KXS administration significantly increased BDNF levels at 500 and 250 mg/kg b.w. per day, and also increased p-CREB/CREB levels at 250 mg/kg b.w., without dose dependency (Fig. 6).
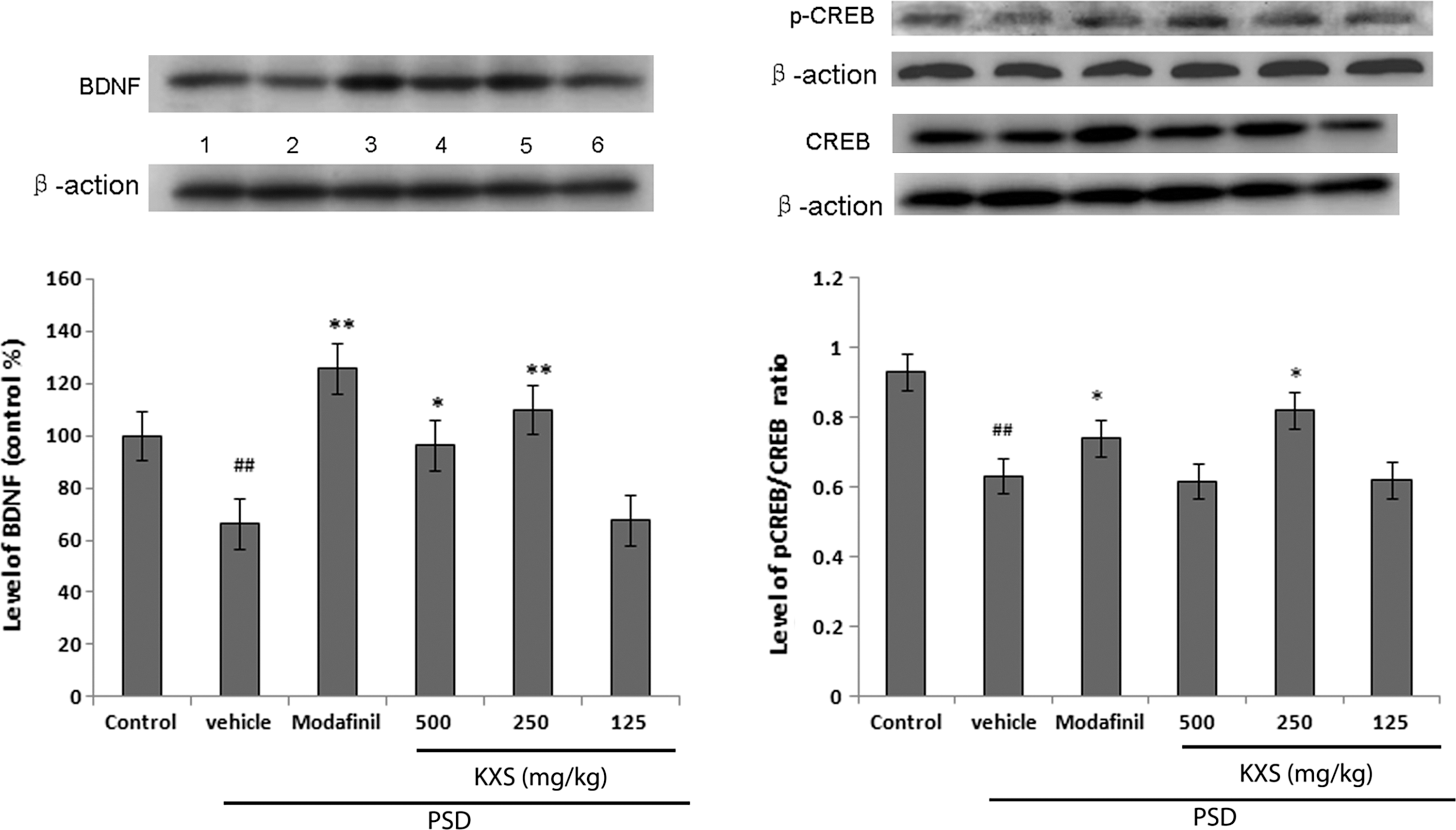
Effect of KXS treatment (500, 250, 125 mg/kg b.w. per day) on BDNF, p-CREB/CREB protein levels in the hippocampi of PSD treated rats. Western blot analysis of BDNF and p-CREB/CREB protein levels in representative gels and densitometry values. Rats were treated with (1) control; (2) vehicle; (3) modafinil 30 mg/kg b.w. per day; (4) KXS 500 mg/kg b.w. per day; (5) KXS 250 mg/kg b.w. per day; (6) KXS 125 mg/kg b.w. per day. Values given are the mean±SEM (n=6). ## P<.01 as compared with the control group; *P<.05 and **P<.01 as compared with the vehicle-treated PSD group. BDNF, brain-derived neurotrophic factor; p-CREB, phosphorylated cyclic AMP response element binding protein; b.w., body weight.
Discussion
PSD, by itself or as a consequence of the technique to induce it, is a stressor and influences a number of physiological mechanisms, such as food intake, thermoregulation, immune function, and modulation of the serotoninergic, dopaminergic, and cholinergic activity. PSD results in an elevation of the serotonergic turnover in the hypothalamus, which partly explained the hormonal results, and in the hippocampus, which appears to be related to adaptive responses to stress. 18 In addition, PSD also has effects on cognitive function not directly related to stress. In contrast to the clear physiological effects of PSD, however, its effects on the cognitive function still remain unclear. A number of studies have suggested that PSD impairs memory consolidation in an eight-arm radial maze, in the Morris water maze, and in other mazes. 19 The MMPM, is a widely used model in PSD 20 to overcome the single platform method limits of the rats moving, surmount the multiple platform method separating one rat from groups, and then decrease the interference of stress. Groups of six animals were used in this test such to reduce the social instability and the attacking behavior 15 between the different group rats. After PSD, animals displayed a decreased locomotor activity in the Open Field test 17,21,22 and impairment of spatial memory in the Morris water maze task. 23 –25 In our present study, KXS treatment with 500 or 250 mg/kg b.w. per day potently enhanced the locomotor activity, which could be considered as a stimulant effect increasing exploration and attention, as well as increasing the learning ability. 26,29,34 Moreover, KXS significantly increased the escape latency and reduced the distance and time in the target quadrant in the Morris test in PSD-induced rats, showing that it has notably improved this ability in PSD rats. On the other hand, KXS has also shown an effect on depression, which can also relieve the PSD-induced stress by adjusting the HPA and some transmitter. 11,12
The regulation of learning and memory requires cooperation between the excitatory and inhibitory neural systems, the representatives of which are the glutamatergic and GABAergic systems, respectively. 30 It has been reported that the spatial memory and avoidance performance deficits in ovariectomized mice are mediated by disturbances in hippocampal, glutamatergic, and GABAergic systems. 31 Also, some reports found that PSD significantly increased the contents of GLU in the brain stem and hypothalamus and raised the levels of GABA in the frontal cortex, hypothalamus, and brain stem. 32 In addition, Bettendorff et al. described that PSD remarkably augmented the levels of GLU in the rat cerebral cortex, whereas GABA only increased during the rebound period. 33 Our data are consistent with these findings suggesting that there would be increased GLU and GABA levels after PSD in brain. In fact, the research had stated that high levels of GLU, as a consequence of NMDA receptor overexcitation, may lead to neurotoxicity followed by neuronal death and therefore leading to induction of LTP. 8 The results of our study demonstrated that KXS decreased GLU levels and reduced GABA levels in brain tissue after 48 h PSD, suggesting that the cognitive-enhancing effect of KXS might be accompanied by decreased levels of GLU and GABA to protect the neurons from neurotoxic damage contributing to the survival of neurons in the brain.
Several studies in mammals have recently demonstrated that CREB is related to the process of learning and memory, such as in the Morris water maze. 35,36 The function of CREB is regulated largely by its phosphorylation at Ser133, which results in the activation of gene transcription for memory formation. 37 It has been reported that radial maze training in rats resulted in a significant increase in p-CREB in the hippocampus in the course of spatial learning. 38 Furthermore, BDNF as active CREB downstream target proteins is capable of augmenting ongoing neurogenesis in the brain. Recent studies demonstrated that the increased BDNF levels in the brain enhanced spatial learning and memory function. 12 In our previous studies, we demonstrated that KXS increases the expression of p-CREB in CA1, CA3, and DG in the hippocampus of the chronically stressed rats, 39 3,6′-disinapoyl sucrose, a bioactive component of KXS, reversed stress-induced alterations in CAM-L1, laminin, CREB and BDNF mRNA and protein levels in the hippocampus. 40 In this study, after 48 h PSD, KXS modulated the PSD-induced decrease in CREB, p-CREB, and BDNF protein levels.
Conclusion
In this study, KXS improved learning and memory deficits in a PSD rat model. The effect may be due to its ability to protect the neurons from neurotoxicity damage by decreasing the levels of GLU and GABA and to augment ongoing neurogenesis by increasing the CREB, p-CREB, and its downstream gene BDNF.
Footnotes
Acknowledgments
This work was supported by the National Nature and Science Foundation of China (No. 30973891, No. 81173430, No 81173579) and the funds from National Science and Technology Major Project of the People's Republic of China (No. 2008ZXJ09004-028).
Author Disclosure Statement
No competing financial interests exist.