
Abstract
In this study, we investigated the effects of genistein, daidzein, and soy protein on paraoxonase and arylesterase activity, malondialdehyde (MDA) levels, and lipid profiles of arthritic rats in vivo and the results were compared with that of dexamethasone. Seventy-two female Sprague-Dawley rats were divided into six groups: healthy control, animals with collagen-induced arthritis (CIA), CIA-soy protein (7 g/kg)-treated rats, CIA-genistein (20 mg/kg)-treated animals, CIA-daidzein (20 mg/kg)-treated rats, and CIA-dexamethasone (1 mg/kg)-treated rats. Rheumatoid arthritis was induced using collagen type II and the treatments were carried out by daily gavages feedings for 50 days. The paraoxonase activity in serum was measured spectrophotometrically using paraoxon and phenylacetate as substrates. Serum MDA and lipids levels were determined using enzymatic colorimetric methods. Arthritis-induced decreases in paraoxonase and arylesterase activity was restored after treatment with soy protein and isoflavones (P<.05). MDA concentrations were lower after treatment with all tested compounds. However, only soy protein could partially improve the lipid profile.
Introduction
R
We recently demonstrated the protective effects of soy protein plus two isolated isoflavones from soy, genistein, and daidzein, in comparison with dexamethasone on arthritic rats. 2,3 Soy protein, genistein, and daidzein were shown to decrease arthritis symptoms and the serum concentration of inflammatory mediators, such as tumor necrosis factor alpha (TNF-α), interleukin (IL)-6, adiponectin, and leptin. Prevention of the tissue damage and joint inflammation was also observed following treatment. 2,3 Apart from TNF-α, IL-6, adiponectin, and leptin, paraoxonase and reactive oxygen species (ROS) are also thought to play key roles in the pathogenesis of RA. Considering the antioxidant effects of soy isoflavones and their protection against RA, there is the possibility that soy intake may reverse the oxidative processes in RA. Paraoxonase is a Ca-dependent esterase distributed in liver, kidney, intestine, and the serum, 4 which prevents from lipoperoxide aggregation in low-density lipoprotein–cholesterol (LDL-C). Paraoxonase is supposed to possess peroxidase, arylesterase, and paraoxonase activities and has been associated with a protective role in oxidative stress and atherosclerosis pathogenesis, which returns to paraoxonase's ability in hydrolysis of lipid peroxidases. 5,6 Studies indicate that paraoxonase concentrations are low in RA patients and are associated with a lower high-density lipoprotein–cholesterol (HDL-C) and a higher level of malondialdehyde (MDA) and C-reactive protein. 7 –9 Mechanisms involved in the decreased paraoxonase in RA are unknown; however, it is speculated that ROS overproduction leads to increased deactivation of paraoxonase. 9 Due to the correlation of paraoxonase with lipid profiles, measuring factors involved in lipid profiles, such as HDL-C, are indicated. One recent study has shown that soy intake in menstruating women for 10 weeks (50 g/day) increases paraoxonase and HDL-C serum levels, but decreases the concentration of other lipids. 10 Paraoxonase is responsible for antioxidant effects of HDL-C and, thus, has also a key role in coronary artery disease. Further, lipid peroxidation is a common outcome of oxidative stress.
In the present in vivo study, the antioxidant effects of soy protein and its isoflavones, genistein, and daidzein on paraoxonase activity and lipid profiles in collagen-induced arthritis (CIA) rats as an animal model of RA have been investigated. The results obtained have been compared with those of dexamethasone, a potent NSAID.
Materials and Methods
Materials
Genistein and daidzein were purchased from LC Labs. Isolated soy protein (Soypro 900) was obtained from Qingdao Crown. Bovine collagen type II and incomplete Freund's adjuvant (IFA) were acquired from Chondrex and Sigma, respectively. Other chemicals were purchased from Sigma-Aldrich.
Animals
Seventy-two female Sprague-Dawley rats (8–10 weeks of age, weighing 180–200 g) were provided from Laboratory-Animal House of Tabriz University of Medical Sciences, Iran. Housing conditions and experimental procedures were in accordance with international standards. 11 Cages were placed under pathogen-limiting conditions in a colony room with strict temperature and humidity control (22°C–24°C and 45%–50% relative humidity) with a 12-h day–night cycle.
Induction of CIA rats
Collagen type II plus adjuvant was used to induce RA in rats according to the protocol provided by Chondrex Inc. Collagen type II was dissolved in 0.01 M acetic acid (2 mg/mL) by shaking overnight at 4°C and used as a fresh solution. To make an emulsion of collagen type II and IFA, ultra-homogenizer (Ultra-Turrax T 25 Basic IKA-WERKE, 24,000 rpm with a 5-mm blade) was used. All mixing processes were carried out in an ice tank. Then, 2.5 mL of IFA was added to an eppendorf tube and the prepared collagen type II was added drop-wise to yield a homogeneous solution. Mixing continued at 24,000 rpm until a thick emulsion was obtained and emulsion stability was tested by adding one drop of the emulsion into a beaker of water (A stable emulsion appeared as a solid clump in water without dispersion.). Finally, 100 μL of the cold emulsion was subcutaneously injected to the base of the tail 2–3 cm from the body on day 0 and the procedure was repeated at day 7.
Study design
Rats were randomly divided into 12 equal groups of six members (healthy controls fed 7 g/kg casein; CIA-controls fed 7 g/kg casein; CIA-soy protein-treated rats fed 7 g/kg soy protein; CIA-genistein-treated rats fed 7 g/kg casein+20 mg/kg genistein; CIA-daidzein-treated rats fed 7 g/kg casein+20 mg/kg daidzein; and finally, CIA-dexamethasone-treated rats fed 7 g/kg casein+1 mg/kg dexamethasone). Casein was used as an ineffective protein instead of isolated soy protein in volumes equal to final volumes of gavages. 12 Treatments were carried out for two time courses of 30 and 50 days after CIA induction. Doses were chosen based on approximate minimum doses prescribed in other animal studies. 13 –15
Genistein, daidzein, and dexamethasone were dissolved daily in dimethyl sulfoxide (DMSO)/water (1 mL/9 mL) and mixed with casein (20 mg isoflavone and/or 1 mg dexamethasone+1 mL DMSO+9 mL water+7 g casein). Then, according to the weight of an animal, 1.7 mL of the preparation (approximately) was fed by oral gavage to the rats two times daily. In the healthy control group, DMSO, water, and casein in the same ratios and volumes, but without isoflavone, were fed to the animal. The soy-isolated protein (7 g/kg per day) was prepared similarly (including DMSO and water), but without addition of casein.
Rats were gavaged with test compounds from 1 week before the induction of CIA two times a day (8 a.m. and 2 p.m.) for 30 and 50 days to achieve the required serum concentrations of the active compounds. 16,17
Measurement of paraoxonase and arylesterase activity
The paraoxonase activity was determined by spectrophotometric analysis using paraoxon (O, O-diethyl-o-p-nitro-phenylphosphate) as the substrate and measuring the increase in the absorbance at 412 nm due to 4-nitrophenol formation. 18 Briefly, the activity was determined by adding 20 μL of serum to the Tris-HCl buffer (100 mM, pH 8.0) containing 2 mM calcium chloride (CaCl2) and 2 mM of paraoxon at 25°C and the rate of 4-nitrophenol formation was determined at 412 nm with a spectrophotometer (Shimadzu 2550, UV/Vis with a temperature control unit). The paraoxonase activity is expressed in nM/mL serum per minute. 19
The arylesterase activity of serum was also measured spectrophotometrically using phenylacetate as the substrate. The generated phenol was measured using a spectrophotometer (Shimadzu 2550, UV/Vis with a temperature control unit) at 270 nm. The reaction mixture contained Tris-HCl (100 mM, pH 8.0), phenylacetate (2 mM), CaCl2 (2 mM), and 10 μL of serum. The activity of arylesterase was expressed in μM/mL serum per minute. Both assays were repeated three times.
Determination of MDA level
Serum MDA as a measure of lipid peroxidation was determined spectrophotometrically. The method is based on the reaction of MDA with thiobarbituric acid (TBA). The MDA-TBA colored complex was formed using 5% trichloroacetic acid, extracted by n-butanol, analyzed by a spectrophotometer (Shimadzu 2550, UV/Vis) at 532 nm, 20 and reported as μM. The calibration curve was constructed using malondialdehyde tetrabutylammonium salt.
Lipid profile
The total cholesterol level was determined by an enzymatic colorimetric assay described by Moghadasian et al.
21
The HDL-C level was also determined by an enzymatic method.
22
In this assay, the HDL-C concentration is analyzed after precipitating very low–density lipoprotein–cholesterol, LDL-C, intermediate-density lipoprotein–cholesterol, chylomicrons, and α-lipoprotein. Triglycerides were measured by an enzymatic method.
23
This method is more precise than other methods, since it does not measure phospholipids and glucose. LDL-C levels were calculated according to the Friedewald formula:
where TC is total cholesterol and TG is total glucose.
Statistical analysis
The results are expressed as the mean±SEM (n=6). All data were evaluated statistically by analysis of variance, and the significance of differences was assessed by the one-way Tukey–Kramer method; P values<.05 were considered as significant.
Results
Paraoxonase and arylesterase activity
No statistically significant differences were observed in the average level of serum paraoxonase activity in rats between 30- and 50-day treatment groups (data not shown). After 50 days of treatment, all tested compounds were successful in restoring the serum paraoxonase activity (Fig. 1).
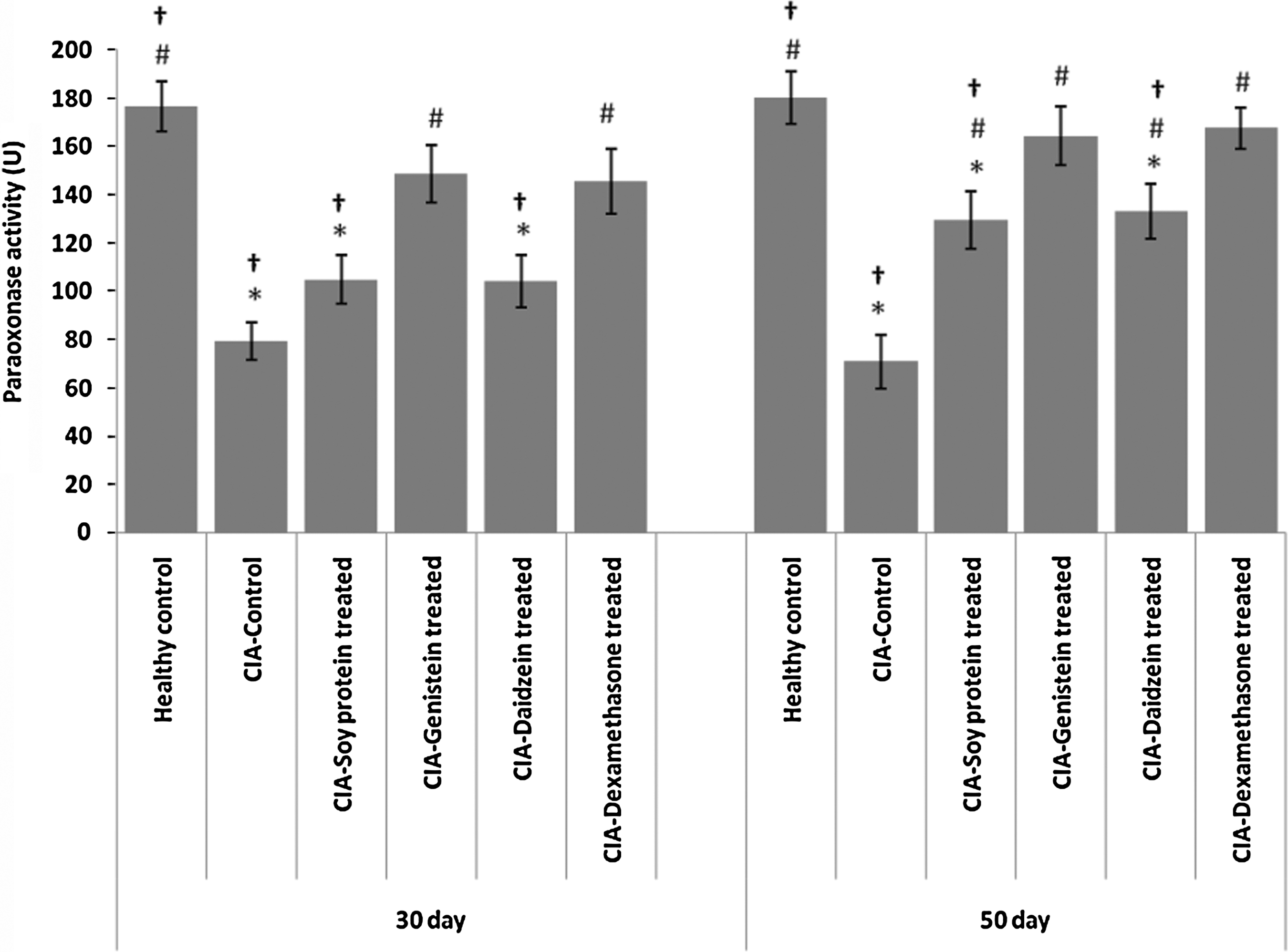
The effect of genistein, daidzein, and soy isolated protein on serum paraoxonase activity in collagen-induced arthritic rats after 30 and 50 days. All data are present as mean±SEM (n=6 for each group). *P<.05 when compared with healthy control; # P<.05 when compared with collagen-induced arthritis (CIA) control; † P<.05 when compared with dexamethasone group.
The serum arylesterase activity also decreased in CIA rats relative to healthy control rats after 30 days of CIA induction. Although the isolated soy protein and daidzein could not increase the arylesterase activity as much as genistein and dexamethasone after 30 days, all test compounds produced significantly different means from the CIA control group and restored the arylesterase activity. Both soy protein and genistein were found to be almost as effective as dexamethasone in restoring the arylesterase activity. Although the enzyme activity increased after treatment with daidzein, it was still statistically different from that of dexamethasone (Fig. 2).

The effect of genistein, daidzein, and soy isolated protein on serum arylesterase activity in collagen-induced arthritic rats after 30 and 50 days. All data are present as mean±SEM (n=6 for each group). *P<.05 when compared with healthy control; # P<.05 when compared with CIA control; † P<.05 when compared with dexamethasone group.
MDA activity
Induction of RA in rats increased the serum concentration of MDA considerably relative to the healthy control group (P=.000). It was interesting that after 30 days, all treatments could decrease the MDA concentration significantly compared to CIA control and genistein decreased it the most after dexamethasone. Results of the 50-day study were consistent with the 30-day treatments. Genistein efficacy increased significantly compared to the 30-day group, which indicates that the effect of these isoflavones on MDA reduction increases over the course of time (Fig. 3).
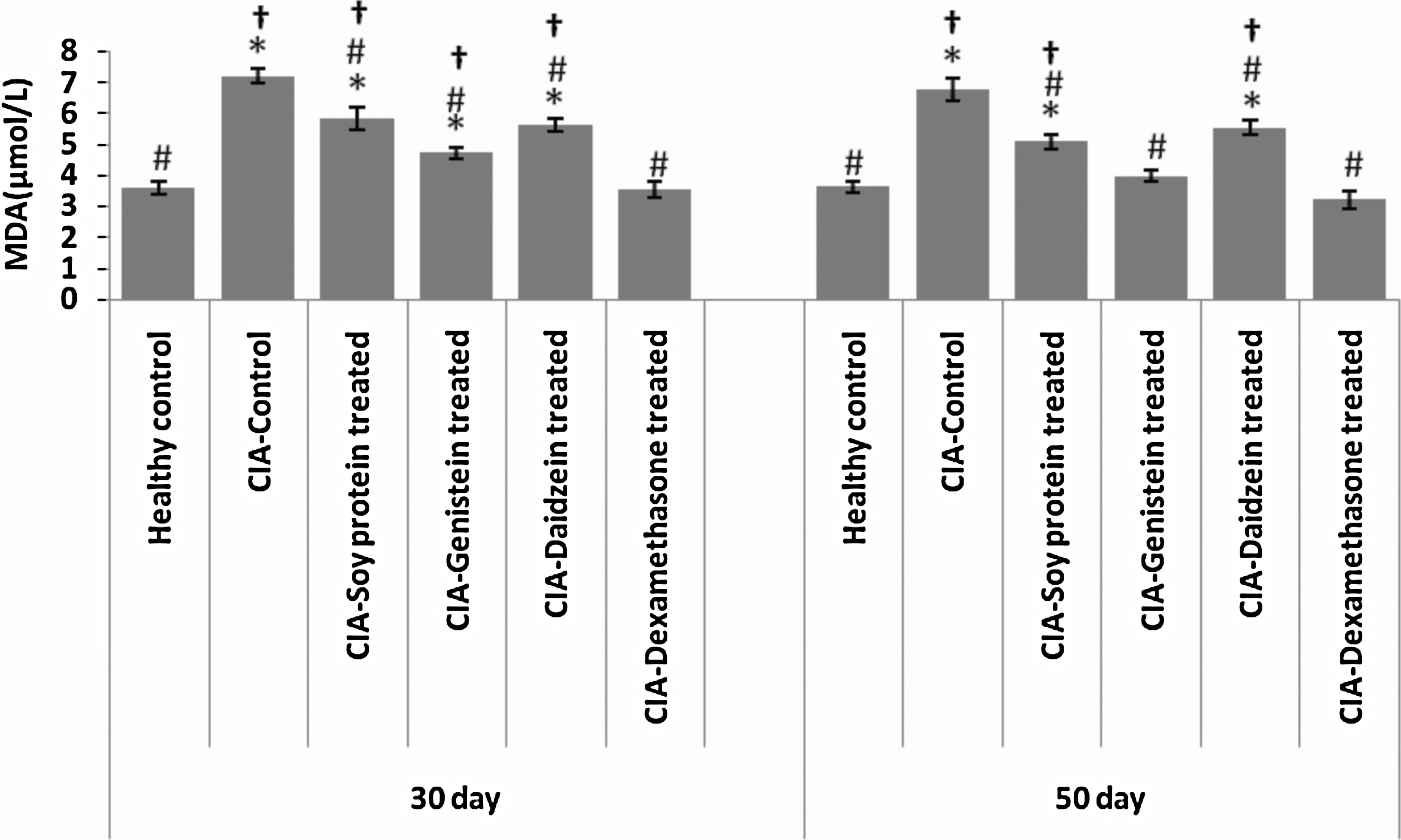
The effect of genistein, daidzein, and soy isolated protein on serum MDA in collagen-induced arthritic rats after 30 and 50 days. All data are present as mean±SEM (n=6 for each group). *P<.05 when compared with healthy control; # P<.05 when compared with CIA control; † P<.05 when compared with dexamethasone group.
Lipid profile
Table 1 summarizes the results of lipid profile analysis (total cholesterol, triglyceride, HDL-C, LDL-C, and LDL-C/HDL-C ratio) of rats after 30- and 50-day treatment with genistein, daidzein, soy isolated protein, and dexamethasone.
All data are presented as mean±SEM (n=6 for each group).
Compared to CIA-control (P=.000) and dexamethasone-treated group (P=.012).
Compared to CIA-control (P=.034).
Compared to CIA-control (P=.015) and dexamethasone-treated group (P=.011).
Compared to CIA-control (P=.036) and dexamethasone-treated group (P=.010).
Compared to CIA-control (P=.040) and dexamethasone-treated group (P=.018).
Compared to CIA-control (P=.049) and dexamethasone-treated group (P=.012).
HDL-C, high-density lipoprotein–cholesterol; LDL-C, low-density lipoprotein–cholesterol; CIA, collagen-induced arthritis.
CIA induction in rats did not have a significant effect on total cholesterol of serum. No treatment, neither after 30 days nor after 50 days, could produce significant changes in cholesterol levels.
After 30 days, no treatment had any significant effect on the serum triglyceride concentration, but after 50 days, the soy-isolated protein significantly lowered serum triglyceride levels compared to CIA control and dexamethasone groups.
CIA induction after 30 days had no significant effect on the HDL-C serum concentration compared to healthy control. After 30 days, only the soy-isolated protein could significantly elevate HDL-C in serum compared to CIA control and dexamethasone groups and was also marginally significantly different from healthy controls (P=.067). Results after 50 days were similar to and not significantly different from 30-day treatments. Similarly, the soy-isolated protein increased the serum HDL-C level compared to CIA control and dexamethasone groups.
While after 30 days, no intervention produced significantly different results in LDL-C levels; after 50 days, the soy-isolated protein significantly decreased serum LDL-C concentrations compared to the dexamethasone group.
After 50 days, only the soy-isolated protein decreased the LDL-C/HDL-C ratio compared to CIA control and dexamethasone groups.
The final body weights of the CIA control group were significantly lower compared with the healthy control group after 30 and 50 days. After 30 days, the soy protein, genistein, and dexamethasone significantly increased the body weight of the treated groups. There was also a significant increase in body weight in genistein- and dexamethasone-treated groups compared with the CIA control group after 50 days.
Discussion
In 2003, the level of paraoxonase activity and its relation to lipid profile in RA patients was investigated. 7 While the paraoxonase activity was lower in RA patients, lipid profiles were the same. Tanimoto et al. stated that structurally and functionally remodeled HDL-C in RA patients has lower anti-atherogenic properties and suggested that reduced activity of serum paraoxonase activity and remodeled HDL-C are responsible for a higher cardiovascular mortality in RA patients. 7 Baskol et al. demonstrated significantly a higher MDA and a lower paraoxonase activity in RA patients with a meaningful negative correlation between MDA and paraoxonase, indicating a negative correlation between the paraoxonase activity and lipid peroxidation. 8 They claimed that higher concentrations of TNF-α and IL-1 in RA patients are responsible for the decreased paraoxonase activity. 8,24 Researchers determined the serum level of paraoxonase and arylesterase of 47 RA patients and 23 healthy subjects and found lower levels of the enzymes in RA patients compared with healthy volunteers. The lower paraoxonase activity in serum will facilitate atherosclerosis progression. 9 Recent epidemiologic studies indicate that mortality rates are higher in RA patients than in the general population. Cardiovascular diseases have been declared the most important cause of death among RA patients, which may be due to impaired antioxidant systems, such as paraoxonase. One reason for a lower paraoxonase activity in RA is a higher production of free radicals and a higher lipid peroxidation that increases the deactivation of paraoxonase, since paraoxonase is involved in detoxification of lipid peroxides. 9 Oxidative stress increases in RA patients resulting in the lower endogenous antioxidant reservoirs both having important roles in the pathogenesis of RA. Paraoxonase is supposed to be an inducible enzyme and can be affected by nutritional regimens 25,26 as shown also by the current study. The increased activity of paraoxonase and arylesterase following treatment with the soy protein and its isoflavones in this study can, partly, be attributed to their antioxidant properties. 27 A similar study in menstruating women with a metabolic syndrome treated with the soy protein for 8 weeks found an increased total antioxidant capacity in serum in the patients. 28 Further, genistein has been shown to increase the antioxidant enzyme activity in serum and several tissues. 29 Oh et al. also found that the isoflavone intake in women with diabetic retinopathy increases the activity of glutathione peroxidase and improves the status of oxidative stress biomarkers. 30
MDA levels in current research were significantly higher in the CIA group than healthy controls, a result that is consistent with earlier studies 8 and is speculated to be associated with higher TNF-α levels associated with inflammation. Inflammatory cytokines increase the production of free radicals and disturb the oxidant/antioxidant balance leading to oxidative damages. Cellular membrane lipids are mostly attacked by the free radicals, consequently resulting in high lipid peroxidation and a greater oxidative damage. 8,9 Higher TNF-α and IL-1 levels cause oxidation of LDL-C and, thus, facilitates lipid peroxidation. 31 In oxidative stress, the concentration of enzymatic and nonenzymatic antioxidants in plasma and tissues decreases, 32 thus, increasing the importance of food antioxidants for neutralizing free radicals and impeding depletion of enzymatic (superoxide dismutase and glutathione peroxidase) and nonenzymatic (vitamins C and E and β-carotene) antioxidants in the biological system, eventually leading to a decrease in lipid peroxidation. 33 In this study, treatment with genistein, daidzein, and the soy-isolated protein decreased MDA levels in plasma after both 30 and 50 days of treatment. Tikkanen et al. in a study in healthy volunteers showed that soy intake containing 60 mg isoflavones delays the oxidation of LDL-C particles. 34 LDL-C resistance to oxidation increases with soy intake. 35 In another study, the serum MDA level decreased with daily intake of 30 g soy for 8 weeks and the authors attributed this effect to soy isoflavones. 28 It has been shown that flavonoids and isoflavonoids are able to neutralize ROS. 27 Possible mechanisms include chelating free metal ions and inhibiting the oxidation of LDL-C particles. 36,37 This study, for the first time, focused on the effects of the soy protein and its isoflavones on oxidative stress in RA. Genistein had a more potent effect on lipid peroxidation than daidzein and soy protein and this difference can be illustrated by the chemical structure of genistein and the positioning of hydroxyl groups on aromatic rings. 27 The genistein effect on lipid peroxidation marker was also time dependent as MDA levels were significantly lower after 50 than 30 days. Treatment with dexamethasone also decreased lipid peroxidation, which was similar to the results of earlier studies. 38,39
RA induction in rats could not exert a significant effect on lipid profiles compared with healthy rats. Treatment with the soy-isolated protein for 50 days caused a significant decrease in the serum triglyceride level compared to CIA controls and the dexamethasone group, and was marginally significant compared with the healthy control group (P=.061). After 30 days, the soy-isolated protein could also increase serum HDL-C compared to the CIA control and dexamethasone groups. After treatment with the soy protein for 50 days, a significant increase in the HDL-C concentration and the decrease in the LDL-C level were observed compared to the CIA control. The results of different studies on the effects of soy proteins and its isoflavones on serum lipid profile have been controversial. While some studies reported no significant effect of soy protein and/or isoflavones on serum lipids, 40,41 other studies reported positive and beneficial effects. 42 –44 It seems that some soy components, such as trypsin inhibitors, amino acids, saponins, and isoflavones, are involved in decreasing cholesterol concentrations. 45 Trypsin inhibitors may be able to decrease cholesterol by increasing cholecystokin in secretion and by inducing the synthesis of bile acids from cholesterol. 46 While methionine and lysine are known to increase serum cholesterol, arginine can decrease serum cholesterol. The higher ratio of arginine to methionine and lysine in soy protein may decrease the serum cholesterol concentration by decreasing the ratio of insulin to glucagon and subsequently suppressing lipogenesis. 29,47 Saponins may also decrease cholesterol by increasing the excretion of bile acids. 48 Like estrogens, phytoestrogens can reduce the LDL-C concentration and increase HDL-C levels, although these effects of phytoestrogens are weaker than the estrogen effects. Another mechanism by which the soy protein might lower cholesterol in serum may be related to modulation of the LDL-C receptor. Upregulation of the LDL-C receptor has been noted in human and animals after soy protein intake. An increased HDL-C level can facilitate the transport of cholesterol from tissues to liver.
Autoimmunity plays a major role in the development of dyslipidemia and atherosclerotic plaque formation in many rheumatological conditions, such as RA. The mechanisms underlying these changes include the interplay of inflammation and auto-antibody formation. 49 Recent research has shown that inflammation leads to proatherogenic changes in lipoprotein metabolism and an increased disease activity is associated with lower TC levels and even more depressed HDL-C levels and lowered apo A1 levels. Management of dyslipidemia should be considered as a part of cardiovascular risk management in RA patients. It is evident that tight disease control lowers the cardiovascular risk in RA and may have also some beneficial effects on lipid profiles. 50
In summary, CIA results in a significant decrease in serum paraoxonase and arylesterase activity similar to other reports on immune-related problems, such as Sjögren's syndrome and RA. 9,51 The isolated soy protein and its isoflavones, daidzein and genistein intake for either 30 or 50 days are able to restore the enzyme activity. Both soy protein and genistein were found to be almost as effective as dexamethasone in restoring the enzyme activity. Soy protein and its isoflavones can also reverse the increased levels of MDA resulting from CIA, with genistein having the greatest effect. However, only soy protein could partially improve the lipid profile.
Footnotes
Acknowledgments
This work was supported by a grant from the Nutrition Research Center, Medical Faculty and Students' Research Committee, the Tabriz University of Medical Sciences. This is a report of a database from thesis entitled “Effect of soy protein, genistein and daidzein on delay of rheumatoid arthritis in rats.”
Author Disclosure Statement
The authors declare no conflict of interests.