
Abstract
Skeletal muscle is a major organ that is important for whole-body glucose metabolism. We found that when isolated rat epitrochlearis muscle was incubated with a Pu-erh tea hot-water extract (PTE) for 30 min, the rate of 3-O-methyl-D-glucose (3MG) transport increased in the absence of insulin. This activation was associated with an increase in Ser473 phosphorylation of Akt, a signaling intermediary leading to insulin-dependent glucose transport, but not Tyr458 phosphorylation of phosphoinositide 3-kinase p85, an upstream molecule of Akt. PTE-stimulated 3MG transport was also not accompanied by Thr172 phosphorylation of the catalytic α-subunit of 5′-AMP-activated protein kinase (AMPK). Gallic acid, a water-soluble ingredient in Pu-erh tea, stimulated Akt phosphorylation, but not AMPK phosphorylation. These results suggest that Pu-erh tea potentially promotes skeletal muscle glucose transport at least in part by activating Akt.
I
Pu-erh tea is made with leaves from old wild tea trees, Camellia assamica (Mast.) Chang. It is mainly produced in the Yunnan Province of China, and is consumed as a health beverage in Southeastern Asia. Pu-erh tea is used as a medicine, tonic, and energy source to enhance well being, and is believed to alleviate a number of physical disorders, including type 2 diabetes. Although the antidiabetic property of Pu-erh tea has not been proven yet in humans, recent work has reported the hypoglycemic effect of the Pu-erh tea extract in rodents. 7 In this study, we investigated whether the Pu-erh tea hot-water extract (PTE) stimulates the signaling mechanisms leading to glucose transport in skeletal muscle.
Dried Pu-erh tea leaves (Jindamo Pu-erh tea, No. DB53/103) were purchased from the Kunming Jinyuxuan Economy & Trade Co., Ltd. (Kunming, Yunnan, China). The leaves (2 g) were powdered and extracted with 100 ml of boiling water for 30 min. The extract was filtered through a filter paper (No. C030633; Toyo Roshi, Tokyo, Japan), concentrated to a volume of 1/10 (v/v) of the initial solution volume by heating at a nonboiling temperature near to 100°C, and then lyophilized with a DC401 Freeze Dryer (Yamato Scientific Co., Ltd., Tokyo, Japan). The dried extract (0.361 g, w/w yield=18.05%) was used as the PTE. PTE included gallic acid (GA) 9.11%, caffeine 2.18%, and epigallocatechin gallate (EGCG) <0.05% as measured by high-performance liquid chromatography.
Protocols for animal were reviewed and approved by the Kyoto University Graduate School of Human and Environmental Studies and the Kyoto University Radioisotope Research Center. Male Wistar rats, aged 5 weeks (Shimizu Breeding Laboratories, Kyoto, Japan), were killed by cervical dislocation, and the epitrochlearis muscles were removed and mounted on incubation apparatus. 8 The muscle was preincubated in 7 mL Krebs–Ringer bicarbonate buffer containing 2 mM of pyruvate (KRBP) for 30 min. To measure the dose response of changes in Akt phosphorylation, the muscles were then incubated in 7 mL of a fresh buffer containing various concentrations of PTE for 30 min. To measure the time course of changes in Akt phosphorylation, the muscles were incubated with PTE (1.5 mg/mL) for up to 60 min. The muscles were also incubated with 1 μM insulin for 30 min to elicit maximal activation of Akt by insulin, and with 2 mM metformin or to elicit maximal activation of AMPK. The muscles were also incubated with 0.14 mg/mL (0.80 mM) of GA, 0.03 mg/mL (0.17 mM) of caffeine, or 0.001 mg/mL (0.002 mM) of EGCG for 30 min to adjust the concentration of each constituent to the level corresponding to 1.5 mg/mL of PTE.
Muscles were transferred to 2 mL of KRB containing 1 mM [
3
H]3-O-methyl-
The muscles were homogenized in a lysis buffer (25 mg/ml). 8 The supernatant (10 μg of protein) was boiled with a loading buffer, and the denatured proteins were separated on a polyacrylamide gel. The proteins were then transferred to polyvinylidene difluoride membranes at 100 V for 1 h. The membrane was blocked with Block Ace (Yukijirushi Nyugyo, Tokyo, Japan) overnight at 4°C, and then incubated with AMPKα, AMPKα Thr172, acetyl Co-A carboxylase (ACC), Akt, Akt Ser473, glycogen synthase kinase (GSK) 3β, GSK3β Ser 9 , phosphoinositide 3-kinase (PI3K) p85 Tyr458 (Cell Signaling Technology, Danvers, MA, USA), ACC Ser79, or PI3K p85 (Millipore, Billerica, MA, USA) antibody. The membrane was reacted with anti-rabbit immunoglobulin G coupled to peroxidase, and developed with enhanced chemiluminescence reagents (GE Healthcare, Buckinghamshire, United Kingdom).
The data are expressed as the mean±SE. The mean values were compared using a one-way analysis of variance followed by a post hoc comparison with Fisher's protected least-significant difference method. Two mean values were compared using the Student's t-test. Differences between the groups were considered significant at P<.05.
Similar to insulin stimulation (1 μM, 30 min), PTE (1.5mg/mL, 30 min) increased the phosphorylation of Akt Ser473, with no change in the total amount of Akt (Fig. 1A, B). The incubation of epitrochlearis muscle with PTE (≥0.38mg/mL, ≥15 min) increased Akt phosphorylation in a dose- and time-dependent manner, with peak levels of phosphorylation at 0.75 mg/mL PTE or more after 30 min or more (Fig. 2A, B). Correspondingly, incubation with PTE (1.5 mg/mL, 30 min) significantly increased the rate of 3MG transport 1.7-fold in epitrochlearis muscle compared with that in the basal condition (Fig. 3). PTE stimulation also increased the phosphorylation of GSK3β, an endogenous substrate of Akt (Fig. 1B). In contrast, PTE did not affect PI3K p85 Tyr458 phosphorylation, an indicator of PI3K activation (Fig. 1B). 9

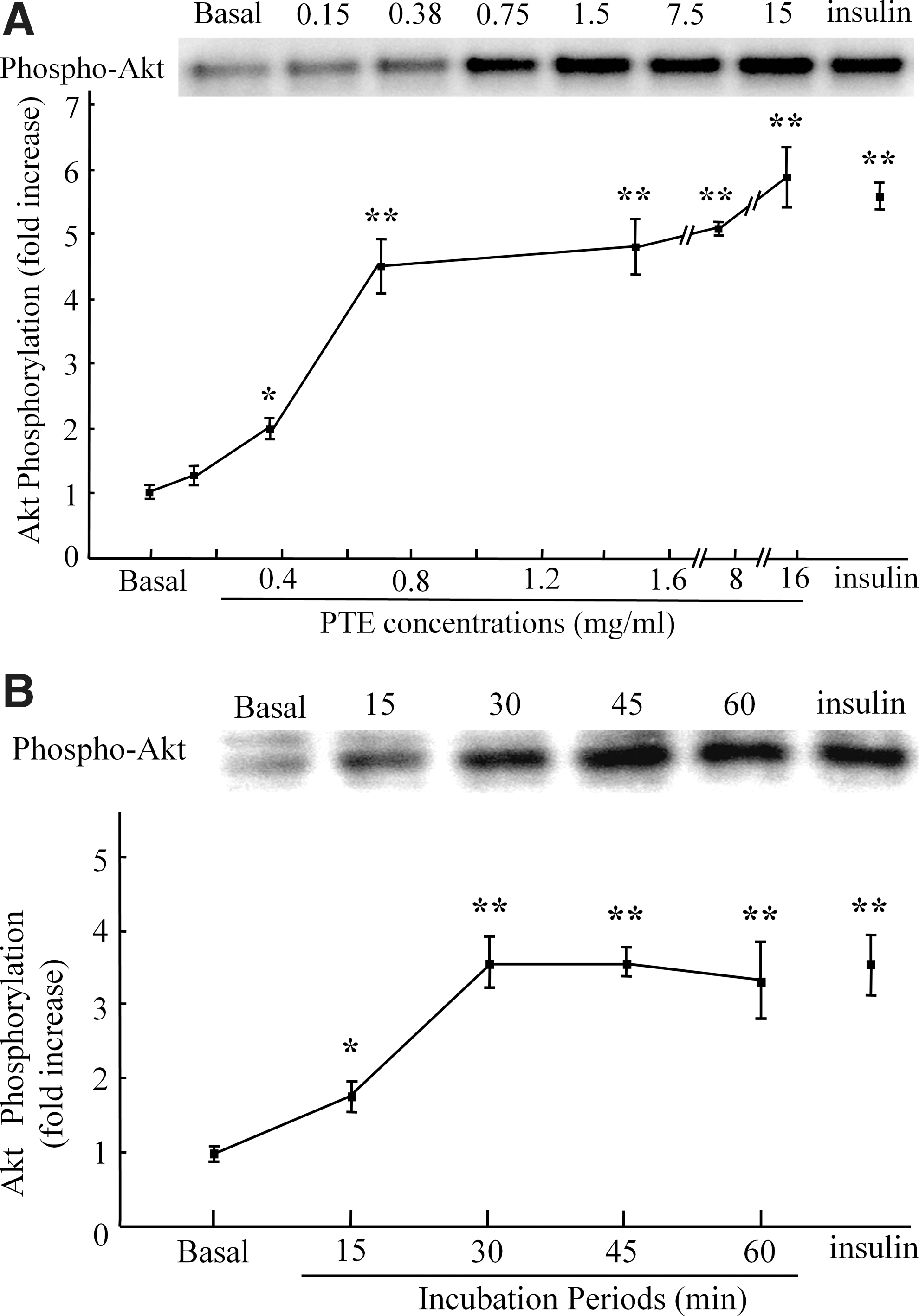
PTE increases phosphorylation of Akt in skeletal muscle in a dose- and time-dependent manner.
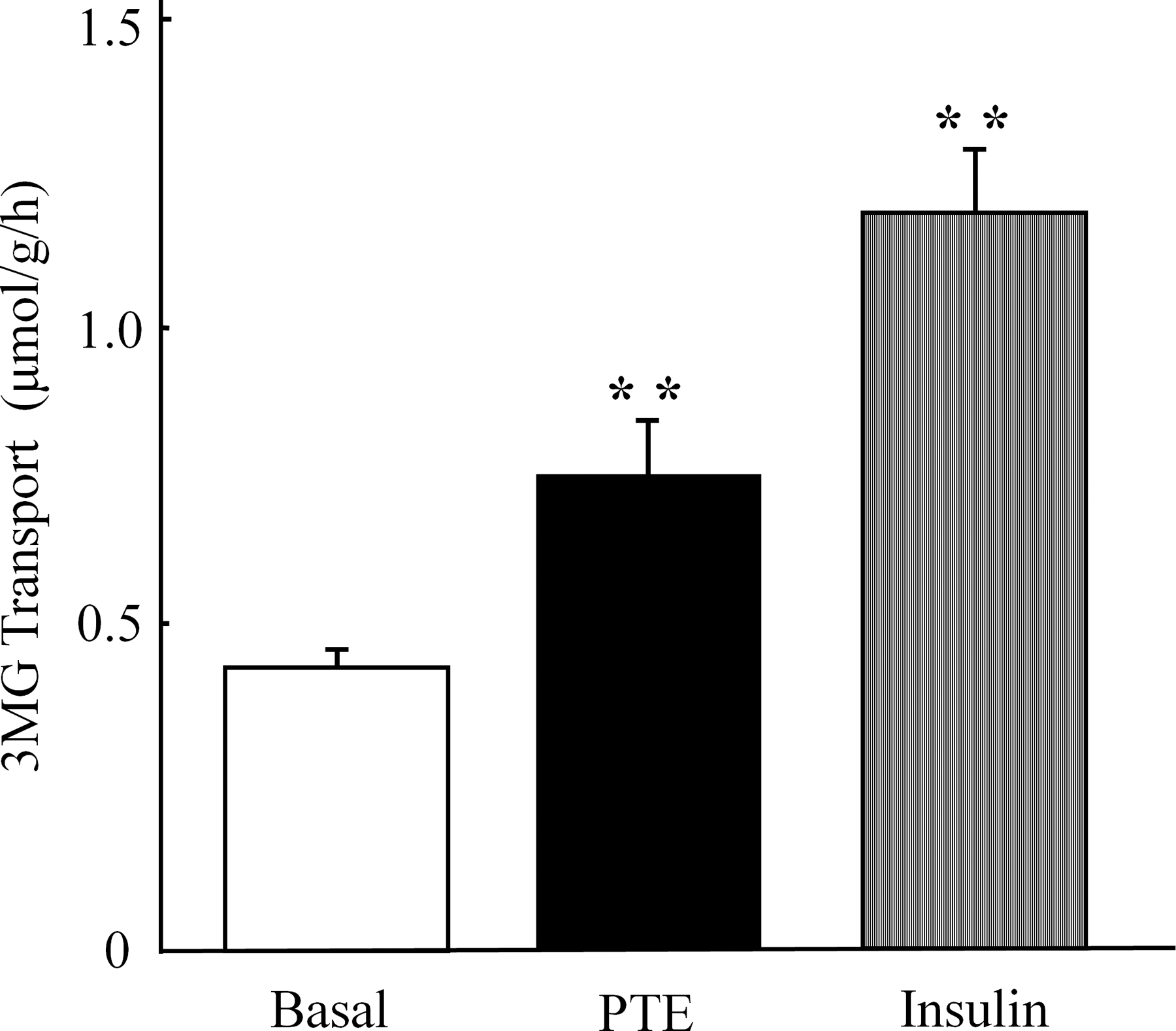
PTE increases 3-O-methyl-
The Thr172 residue in the catalytic α-subunit is the primary site responsible for AMPK activation. 10 PTE (1.5 mg/mL, 30 min) did not cause a change in the phosphorylation status of AMPKα Thr172 relative to the basal condition (Fig. 1). The phosphorylation of ACC, a downstream target of AMPK in skeletal muscle, was not affected by PTE treatment (Fig. 1).
We also examined whether GA, the major ingredient of PTE, stimulates Akt. Similar to PTE stimulation, GA increased the phosphorylation of Akt, with no change in the total amount of Akt (Fig. 4). GA did not alter the phosphorylation status of AMPKα (Fig. 4). On the other hand, caffeine and EGCG had no effect on Akt or AMPK phosphorylation (Fig. 4).

PTE and GA increase phosphorylation of Akt, but not of AMPK in skeletal muscle. Isolated epitrochlearis muscle was preincubated for 30 min, and then incubated in the absence (basal) or presence of EGCG (0.001 mg/mL), caffeine (0.03 mg/mL), PTE (1.5 mg/mL), GA (0.14 mg/mL), or insulin (1 μM) for 30 min. The concentrations of GA, caffeine, and EGCG were adjusted to the concentration of each constituent to the level corresponding to 1.5 mg/mL of PTE. The tissue lysate was subjected to Western blot analysis. Representative immunoblots are shown. EGCG, epigallocatechin gallate; GA, gallic acid.
Akt is a family of serine/threonine protein kinases that is important in glucose transport activity in skeletal muscle. The expression of constitutively active Akt increases both basal and insulin-stimulated glucose transport in cultured L6 myotubes. 11,12 Glucose disposal during euglycemic hyperinsulinemic clamp was reported to positively correlate with Akt phosphorylation and Akt2 activity in nondiabetic humans. 13 Correspondingly, Akt2 knockout mice exhibit hyperglycemia and hyperinsulinemia, 14 concomitant with reduced insulin-stimulated glucose transport in their skeletal muscle. 14,15 Further, siRNA silencing of both the Akt1 and Akt2 isoforms inhibited basal and insulin-stimulated glucose transport, whereas siRNA silencing of either Akt1 or Akt2 reduces basal or insulin-stimulated glucose transport, respectively, in primary human skeletal muscle myotubes. 16 Thus, skeletal muscle Akt is thought to play a crucial role in maintaining normal glucose homeostasis and insulin sensitivity.
Our data show novel findings relating to the metabolic effects of Pu-erh tea in skeletal muscle. First, PTE acutely increased 3MG transport in the absence of insulin. Second, PTE-stimulated glucose transport was accompanied by the phosphorylation of Akt and GSK3β, suggesting that PTE increases Akt activity in vivo. Third, PTE-stimulated glucose transport was not accompanied by the phosphorylation of PI3K, suggesting that the phosphorylation of Akt is not attributable to the activation of PI3K. Fourth, PTE did not affect the phosphorylation of AMPK or ACC, indicating that the AMPK-dependent pathway does not play a role in PTE-induced activation in glucose transport. Lastly, GA, the major bioactive compound in PTE, increased the phosphorylation of Akt, but not of AMPK. Collectively, our results provide fundamental evidence that PTE potentially acts on skeletal muscle and activates insulin-independent glucose transport, at least in part, through GA-induced Akt activation. PTE may induce a metabolic milieu that reduces the risk of developing type 2 diabetes, by activating skeletal muscle Akt. More research is needed to clarify the clinical relevance of the long-term effects of PTE administration at physiologically relevant doses.
Footnotes
Acknowledgments
The authors thank Dr. Shuhoi Hayashi (Hyogo College of Medicine, Nishinomiya, Japan), Dr. Wei Shi (Jilin University, Changchun, China), and Dr. Jun Sheng (Yunnan Agricultural University, Kunming, China) for suggestions. We thank the Radioisotope Research Center of Kyoto University and the Kyoto University Research Center for Low Temperature and Materials Sciences for instrumental support. This study was supported by research grants from the Japan Society for the Promotion of Science, Japan Vascular Disease Research Foundation, Urakami Foundation for Food and Food Culture, and Kao Research Council for the Study of Healthcare Science.
Author Disclosure Statement
No competing financial interests exist.